관련 문서
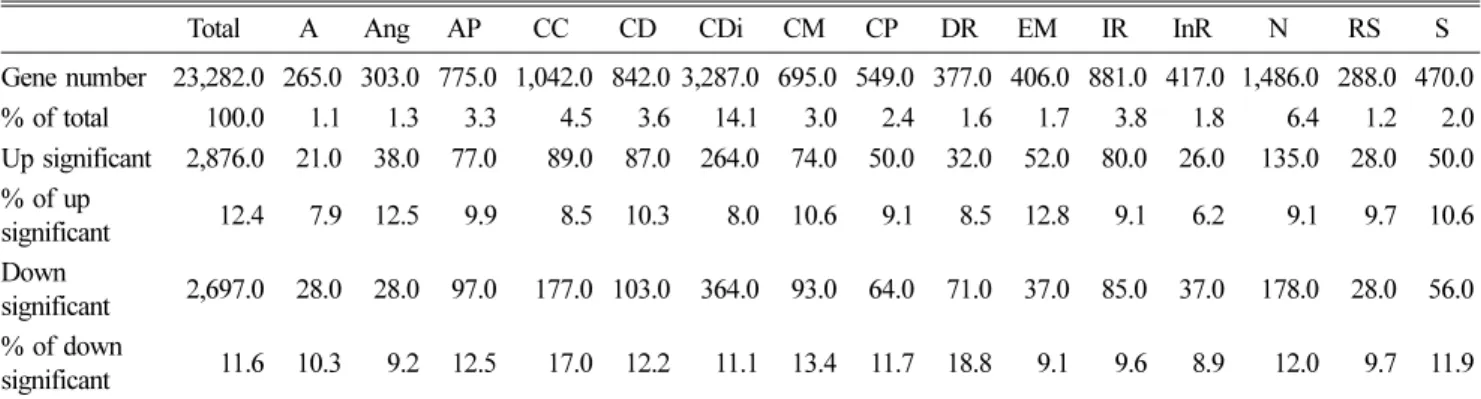
The melanoma cell line WM793 was employed for the Fascin and MST2 knock-down and then analyzed by the Western blot assay, and the melanoma xenograft followed by Fascin
co-treatment with hispidulin and TGF-β up-regulated the protein of expression E-cadherin and occludin against TGF-β-induced in MCF-7 and HCC38 cells.. The
protein levels in control or PIG3 depleted HCT116 and HeLa cells were tested. by
4 > Effects of Taro on COX-2 expression and iNOS expression(hot water) in human thyroid cancer cells. The cells were pretreated for 48hours with either
Design and synthesis of novel threosyl-5_-deoxyphosphonic acid purine analogues as potent anti-HIV agents. Synthesis of novel 3'-hydroxymethyl
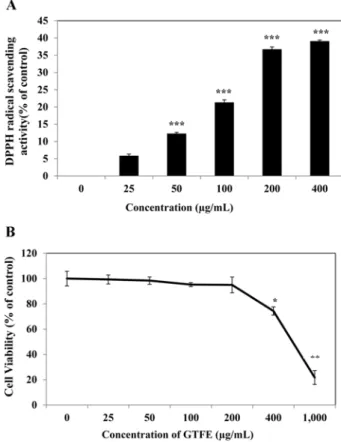
tricuspidata on the production of proinflammatory cytokines in TNFα+IFNγ-stimulated HaCaT cells ...15 Fig.5: The cell viability of sub-fractions from 70% EtOH
synthesis and application of silicon nano particles for improved efficiency in perovksite solar cells.. synthesis and application of silicon nano particles for improved
The biological effect of ionizing radiation is a consequence of the energy transfer, by ionization and excitation, to cells in the body. Factors of radiation effects