Promoter Analysis of the NTR1 Gene of Brassica campestris L.
ssp. pekinensis in Transgenic Tobaccos
Yeon Jong Koo†, Jong Tae Song1,† and Yang Do Choi*
School of Agricultural Biotechnology, Seoul National University, Seoul 151-742, Korea
1Research Center for New Bio-Materials in Agriculture, Seoul National University, Seoul 151-742, Korea Received August 19, 2004; Accepted September 6, 2004
The NTR1 gene is specifically expressed in the floral nectaries of Brassica campestris L. ssp. pekinensis, which encodes an S-adenosyl-L-methionine:jasmonic acid methyltransferase. To understand the regulation mechanism of the NTR1 gene during nectary development, promoter 5'-deletion series were constructed by fusing translationally to a β-glucuronidase (GUS) reporter gene. Histochemical GUS staining and quantitative fluorometric analysis with the transgenic tobaccos indicated that regions spanning about −4.4 kb to −2.1 kb and −1.0 kb to −0.5 kb from the translation initiation site of NTR1, were essential for reasonable expression of GUS reporter gene. Transgenic tobaccos transformed with encompassing either −1.7 kb or -0.5 kb region of the NTR1 promoter caused the basal level of GUS activity. These results suggest that sequences of NTR1 promoter flanking about −4.4 kb to −2.1 kb and about −1.0 kb to −0.5 kb are important as a strong activator and for the nectary-specific expression during nectary development, respectively.
Key words: NTR1, floral nectary, promoter analysis, transgenic tobacco
Nectaries may develop on all aerial parts of the plant.
Nectaries that are connected with the floral organs are termed floral nectaries, and those developing on the vegetative parts of the plant, extrafloral nectaries1). Plants offer a diversity of floral rewards: pollen, nectar, stigmatic exudates, oils, scent- producing compound, resins, gums, and food tissues. Among them nectar is considered the important one2), representing the major energy source for visitor, which inadvertently pick up and disperse pollen while withdrawing nectar. It is secreted by floral nectaries and its main constituents are sugars (sucrose, glucose, and fructose) and water3-5). Therefore, these glands perform a vital function in plant sexual reproduction by attracting pollinators.
Plants emit various volatiles to withstand various environmental stresses and defend themselves. The volatiles may be attractive for pollinators and seed-dispersing animals or repellent for a wide range of herbivores6). A variety of methyl esters of secondary metabolites constitute an important portion of the plant volatiles. For instance, methyl jasmonate (MeJA) is a fragrant compound initiallly identified from the flower of Jasminum grandiflorum7), and is ubiquitously distributed in the plant kingdom8). MeJA and its free acid jasmonic acid (JA), collectively referred to as jasmonates, are important cellular regulators mediating diverse development processes, such as seed germination, flower and fruit
development, leaf abscission, senescence, and defense responses9).
With the goal of understanding the molecular mechanism of nectary development and function, we have isolated and characterized a nectary-specific gene, NTR1, from B.
campestris10), which encodes an S-adenosyl-L-methionine:
jasmonic acid methyltransferase11). An Arabidopsis NTR1 homologue, JMT1, has been cloned and its enzyme activity was similar to that of NTR112). Transgenic Arabidopsis overexpressing JMT exhibited elevated resistance to the fungal and bacterial pathogens Botrytis cinerea12) and Pseudomonas syringae pv. tomato DC300013), respectively.
Both of two genes are strongly expressed during flower development, expecially in the floral nectaries10,12), suggesting that MeJA may important for nectary development and/or pollination by attracting pollinators.
In this paper, the NTR1 promoter of B. campestris was investigated based on GUS reporter gene expression in transgenic tobacco plants. To determine the cis-acting elements regulating expression pattern of NTR1, the promoter activities of a series of 5' deletion were also examined.
Materials and Methods
Isolation of the NTR1 genomic DNA and deletion construction. The genomic clone of NTR1 (g x 8) was obtained by screening a genomic library14). All promoter 5'- deletion series were constructed using either convenient digestion with several restriction enzymes or the polymerase chain reaction (PCR). About 4.4 kb upstream region was
†These authors contributed equally to this work.
*Corresponding author
Phone: 82-2-880-4941; Fax: 82-2-873-3112 E-mail: [email protected]
GACCCTAAC-3') and BCR1568 (5'-GGTGAATTCACATG TATTTTGTCT-3') as primers. The CR indicates −2.1 to −1.7 kb region of the NTR1 promoter from the translation initiation site. For pG1.5 or pG2 constructs, PCR was performed using BC1582 (5'-CTAAAGCTTTTTTTCTTGCCTACC-3') and BCR2211 (5'-TTGATAAGCTTAAGGAAAACAGAA-3'), and BC1126 and BCR2221 as primers, respectively. The PCR products digested were cloned into pG1. One construct using a set of BC1582 and BCR2211 was named pG1.5 and the other was pG2.
Plant transformation. For tobacco transformation, the constructs pG4, pG2, pG1.5, pG1, pG0.5 and pG0.5 + CR were introduced to Agrobacterium tumefaciens strain LBA4404. Agrobacterium-mediated transformation on leaf disks of Nicotiana tabacum cv. Xanti was conducted according to a published procedures15). Regenerated kanamycin-resistant plants were grown in a greenhouse and confirmed by PCR and Southern blot analysis.
Southern blot analysis. Southern blot analysis was performed according to the procedure of Sambrook et al.16) The DNA was separated by 0.8% agarose gel electrophoresis, transferred onto nylon membrane and then hybridized with the probe at 42oC for 12 hours or more.
Histochemical staining. Tissue for GUS staining was fixed by incubation in 90% acetone, on ice, for 15 to 30 min17) and then rinsed extensively with 100 mM phosphate buffer, pH 7.0. Staining solution contained 100 mM phosphate buffer, pH 7.0, 0.5 mM K3(Fe[CN]6), 0.5 mM K4(Fe[CN]6), 0.1%
Triton X-100, 10 mM EDTA, and 0.5 mg/ml 5-bromo-4- chloro-3-indolyl-β-D-glucuronic acid (X-glu, Gold Biotechnology, St. Louis, MO). Tissues were incubated in staining solution for 4 to 24 hr, washed with 100 mM sodium phosphate buffer, and washed with several changes of 70% EtOH to clear the chlorophyll.
GUS fluorometric assay. Quantitative GUS fluorometric assays were conducted according to the protocol of Jefferson et al.18) by measuring the fluorescence of 4-methylumbelliferone (MU, Sigma) formed as a result of the cleavage of 4- methylumbelliferyl-β-D-glucuronide (MUG, Sigma) by GUS with the fluorescence spectrophotometer (Hitachi F-4500, Japan). Protein content of the samples was determined according to the Bradford method19). Whole flowers including about 1 cm pedicel were harvested, and immediately used for the enzyme assay. Emission was measured at 455 nm after fluorescence excitation at 365 nm. Under these conditions accumulation of the reaction product was linear with regard to enzyme amount and time. No change in the enzyme activity was noted if the extract was kept on ice for 3 hr.
initiation codon was determined as shown in Fig. 1. In this study, we considered the −4.4 kb sequence as a full regulatory region of the NTR1 gene because it shows maximum activity and floral nectary-specific expression as described below.
Compared with promoter of Arabidopsis JMT gene, the region covering about −2.1 to −1.7 kb from the translation initiation codon of NTR1 showed high homology with the JMT promoter sequence (Fig. 1), which we named CR (conserved region), suggesting to be a potential functional target for NTR1 to be fully expressed during flower development. The region just near the translation initiation codon may not be a part of the promoter of NTR1, because it was so near to start codon.
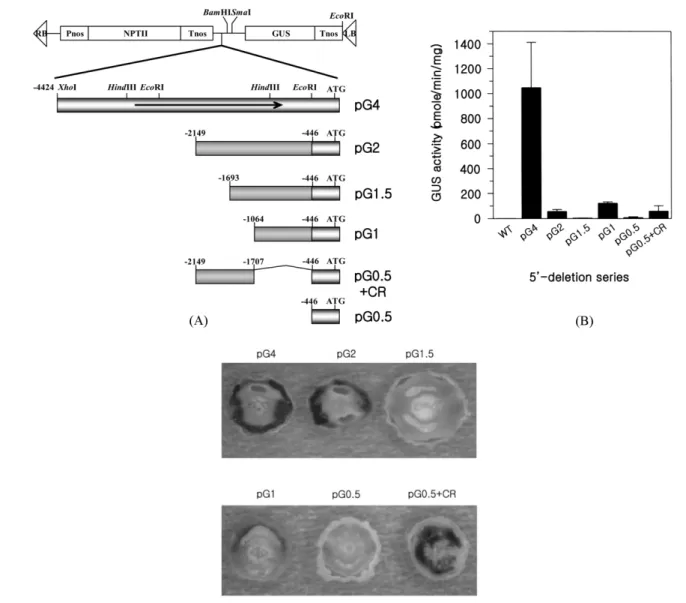
Identification of the transgenic tobacco plants. To identify the NTR1 promoter DNA sequences essential for strong promoter activity and for the floral nectary-specific expression, a series of promoter 5' deletions to −4.4, −2.1,
−1.7, −1.0 and −0.5 kb including −0.5 kb + CR from the translation initiation site, respectively, were constructed as translational fusions with GUS reporter gene (named pG4, pG2, pG1.5, pG1, pG0.5 and pG0.5 + CR, respectively) as shown in Fig. 2A. These expression cassettes were transformed into the tobacco plants. To confirm that the regenerated kanamycin-resistant tobacco plants contain each construct, we performed genomic Southern blot analysis, showing that each cassette DNA was inserted into genomic DNAs of all transgenic plants analysed (data not shown).
5' deletion analysis of the NTR1 promoter. To investigate the effects of 5'-deletion promoter on GUS expression, GUS activities of transgenic plants were quantitatively analysed.
Fluorometric GUS assays were performed using whole flower extracts including pedicel (about 1 cm in length) from at least 3 independent lines per each construct (Fig. 2B). The assay showed that the transgenic plants of the pG4 construct had the highest GUS specific activity. The activity of pG2 encompassing 2.1 kb promoter was declined 20 fold less than that of pG4.
Further deletion from −2.1 to −1.7 kb (pG1.5) caused the basal level of GUS activity. But when the GUS gene was regulated by just 1.0 kb promoter, the activity was recovered to the level of pG2. When the GUS gene was fused to 0.5 kb promoter, the shortest length of promoter in this study, the GUS activity was declined to the basal level. The activity level of pG0.5 + CR had similar to that of pG2. Taken together, these data indicated that −2.1 kb upstream region has a very strong enhancing activity for NTR1 expression, and the region spanning −2.1 to −1.7 kb has a silencing activity.
Histochemical localization of GUS expression driven by 5'-deletion series. Histochemical GUS staining experiments
were performed to localize the GUS gene expression pattern driven by 5'-deletion series. The flowers carrying these constructs were horizontally sectioned, and emersed in staining solution containing X-glu as a substrate. All samples were kept afterwards at 37oC for 20 hr except samples of the pG4 construct were less than for 3 hr. The expression level of pG4 construct was much higher than other constructs (Fig.
2C). pG4 showed that the GUS gene was preferentially expressed in the floral nectary of transgenic tobacco, consistent with our previous data10). pG2 and pG1 expressed GUS gene specifically in the nectary, but the promoter strength of these two constructs was declined, consistent with fluorometric GUS assay (Fig. 2B). The transgenic plants carrying pG1.5 and pG0.5 didn't show any GUS activities
even though their samples were incubated much longer. The pG0.5 + CR construct expressed GUS gene in various floral tissues including the floral nectary, and removal of 5' upstream region encompassing −1.0 to −0.5 kb caused loss of the nectary-specific expression of the reporter gene, suggesting that the nectary-specific regulatory region of the NTR1 gene was at the region encompassing −1.0 to −0.5 kb of promoter.
Thus, the entire 5'-sequence of the NTR1 gene, which encompasses about 4.4 kb, induces reasonable expression levels of the GUS reporter gene compared with other 5'- deletion series. Therefore, the full expression of NTR1 gene is likely to be almost controlled by −4.4 to −2.1 kb promoter region.
Fig. 1. 5' upstream sequence of the NTR1 promoter from B. campestris. The translation initiation codon is shown in bold and highly conserved region with Arabidopsis JMT gene is shown in uppercase letters. The shaded lower sequence is from Arabidopsis JMT gene.
Discussion
Gene transfer technology has enabled a number of plant genes to be introduced into heterologous plants and has facilitated the analysis of qualitative and quantitative aspects of their expression. The demonstration that genes retain regulated pattern of expression in heterologous plants has permitted the delineation of cis-acting sequence elements involved in tissue-specific and developmental control. To understand the spatial and developmental regulation of the NTR1 gene of B. campestris, the objectives of this work was to characterize promoter sequences regulating specific expression of NTR1 gene by introducing it into tobacco.
Previous in situ hybridization and immunolocalization data showed that NTR1 gene is specifically expressed in the floral
nectary tissue of B. campestris10). To determine fidelity of expression of NTR1 in tobacco in terms of nectary-specific expression, chimeric 5'-deletion promoter/GUS fusion constructs was transformed into tobacco and expression of GUS examined in floral nectary.
Expression of GUS in transgenic tobacco plants encompassing 4.4 kb promoter (pG4) is tightly regulated with respect to organ specificity and during nectary development.
Such tissue specific and strong expression is likely due to sequences spanning −4.4 to −2.1 kb for a strong activator and
−1.0 to −0.5 kb for a nectary-specific element involved in the control of transcription via interaction with specific DNA- binding proteins (Fig. 3). The 5' deletion form −1.7 to −1.0 kb abolished the promoter activity, implying that the region may be a repressor-binding element or silencer.
Fig. 2. Construction of 5'-deletion series and GUS activities. (A) Construction map of various 5'-deletion series of the NTR1 pro- moter. Deleted promoters were translationally fused to GUS gene. (B) GUS activities of tobacco flowers transformed with 5'-deletion series constructs. Values are means of five independent experiments. Error bars represent standard deviation. (C) Histochemical stain- ing of GUS enzyme in the nectary of transgenic tobacco transformed with promoter deletion series. Open flowers for 5'-deletion series except for pG4 were cross-sectioned and stained at 37oC for 20 hr. Flowers for 4.4 kb promoter were stained for 3 hr due to the strong GUS activity.
The region encompassing −2.1 to −1.7 kb was highly conserved through all the promoter of NTR1 gene compared with Arabidopsis JMT gene. High homology of this region led us to suggest that the region may implicate an important role on the expression of NTR1 gene during nectary development.
When this region was deleted, the GUS activity was declined to the basal level and disappeared from all the floral organs.
However, GUS activity in pG0.5 + CR lost nectary-specificity, but its level was recovered to pG2, implying that the region − 2.1 to −1.7 kb may operate a very weaker activator than the region −4.4 to −2.1 kb.
The region −4.4 to −2.1 kb is important for NTR1 expression but has very lower homology (<10%) compared with that of JMT. This led us to suggest that similar structural conformation between NTR1 and JMT promoters may require to bind a transcription factor important for NTR1 regulation during nectary development. In addition, AT-hook binding factor was very recently isolated by performing Southwestern screening in our lab and its specific binding was between −4.4 and −3.0 kb promoter region (Seo et al., unpublished), suggesting that the region may play a important role in NTR1 expression by binding of this AT-hook transcription factor.
Our previous data showed that NTR1 catalyzes MeJA production in the cytosol10,11) and MeJA is a strong candidate for airborne signals that mediate intra- and/or interplant communication for defense responses9,12,13). Therefore, further studies on NTR1 expression will be focused on its induction by environmental cues such as biotic and abiotic stresses.
Acknowledgments. This work was supported by a grant from the Crop Functional Genomics Center (Korea). Financial supports including graduate research assistantship to YJK from the Brain Korea 21 Project of the Ministry of Education are also acknowledged.
References
1. Fahn, A. (1990) Plant anatomy. In: The Flower: Nectaries (4th ed.) pp. 463-471, Butterworth-Heinemann Ltd, Oxford, London.
2. Simpson, B. B. and Neff, J. L. (1983) Evolution and diver- sity of floral rewards. In Handbook of experimental pollina- tion biology, Jones, C. E. and Little, R. J. (eds.) pp. 142- 159, Columbia University Press, New York.
3. Baker, H. G. and Baker, I. (1983) A brief historical review of the chemistry of floral nectar. In The biology of necta- ries, Bentley B. and Elias, T. S. (eds.) pp. 126-152, Colum-
bia University Press, New York.
4. Davis, A. R., Fowke, C. V., Sawhney, K. and Low, N. H.
(1996) Floral nectar secretion and ploidy in Brassica rapa and B. napus (Brassicaceae). II. Quantified variability of nectary structure and function in rapid-cycling lines. Ann.
Bot. 77, 223-234.
5. Bernardello, L. M., Galetto, L. and Juliani, H. R. (1991) Floral nectar, nectary structure and pollinators in some Argentinean Bromeliaceae. Ann. Bot. 67, 401-411.
6. Reymond, P. and Farmer, E. E. (1998) Jasmonate and sali- cylate as global signals for defense gene expression. Cur.
Opin. Plant Biol. 1, 404-411.
7. Demole, E., Lederer, E. and Mercier, D. (1962) Isolement et determination de la structure du jasmonate de methyle, con- stituant ordorant characteristique de l'essence de jasmin.
Helv. Chim. Acta 45, 675-685.
8. Hamberg, M. and Gardner, H. W. (1992) Oxylipin pathway to jasmonate: Biochemistry and biological significance. Bio- chim. Biophys. Acta 1165, 1-18.
9. Creelman, R. A. and Mullet, J. E. (1997) Biosynthesis and action of jasmonates in plants. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 48, 355-381.
10. Song, J. T., Seo, H. S., Song, S. I., Lee, J. S. and Choi, Y.
D. (2000) NTR1 encodes a floral nectary-specific gene in Brassica campestris L. ssp. pekinensis. Plant Mol. Biol. 42, 647-655.
11. Seo, H. S., Song, J. T., Koo, Y. J., Jung, C., Yeu, S. Y., Kim, M., Song, S. I., Lee, J. S. Hwang, I., Cheong, J.-J.
and Choi, Y. D. (2001) Floral nectary-specific gene NTR1 encodes a jasmonic acid carboxyl methyltransferase. Agri.
Chem. Biotechnol. 44, 119-124.
12. Seo, H. S., Song, J. T., Cheong, J.-J., Lee, Y.-H., Lee, Y.- W., Hwang, I., Lee, J. S. and Choi, Y. D. (2001) Jasmonic acid carboxyl methyltransferase: A key enzyme for jas- monate-regulated plant response. Proc. Natl. Acad. Sci. USA 98, 4788-4793.
13. Jung, C., Lyou, S. H., Koo, Y. J., Song, J. T., Choi, Y. D.
and Cheong, J.-J. (2003) Constitutive expression of defense genes in transgenic arabidopsis overproducing methyl jas- monate. Agri. Chem. Biotechnol. 46, 52-57.
14. Song, J. T. (1999) In Ph. D. Thesis: Isolation and charac- terization of a floral nectary-specific gene from Brassica campestris L. ssp. pekinensis. Seoul National University, Seoul, Korea.
15. Horsch, R. B., Fry, J. E., Hoffmann, N., Wallroth, M., Eicholtz, D., Rogers, S. G. and Fraley, R. T. (1985) A sim- ple and general method for transfering genes into plants.
Science 227, 1229-1231.
16. Maniatis, T., Fritsch, E. F. and Sambrook, J. (1989) In Molecular Cloning: a Laboratory Manual (2nd ed.) Cold Spring Harbor Laboratory Press, New York.
17. Hemerly, A. S., Ferreira, P., de Almeida Engler, J., Van Montagu, M., Engler, G. and Inze, D. (1993) cdc2a expres- sion in Arabidopsis is linked with competence for cell divi- sion. Plant Cell 5, 1711-1723.
18. Jefferson, R. A., Kavanagh, T. A. and Bevan, M. W. (1987) Fig. 3. Schematic presentation of the NTR1 promoter for its
function.