148
음식물 쓰레기 퇴비화 과정에 따른 세균군집 구조의 변화
신지혜1·이진우1·남지현1·박세용2·이동훈1*
1충북대학교 미생물학과, 2한양대학교 토목공학과
퇴비화과정은유기성폐기물을비료와같은유용한자원으로전환하는생물학적과정이다
.
본연구에서는음식 물쓰레기를2
달동안퇴비화시켜세균군집의변화를조사하였다.
온도의변화를기준으로하여퇴비화과정은1
단계
(2~55
oC), 2
단계(55~97
oC), 3
단계(50~89
oC)
로나뉘었다.
각단계별총세균수는1
단계1.66
×10
11cell/g, 2
단계0.29
×10
11cell/g, 3
단계0.28
×10
11cell/g
으로관찰되었다.
또한총세균수에대한고온미생물의비율은초기에33%
였으나
2
단계시료에서최대비율인89%
로증가하였다. 16S rRNA
유전자를대상으로T-RFLP
방법과염기서열 분석방법을이용하여세균군집의구조가퇴비화과정에따라변화됨을확인할수있었다.
초고온인2
단계의세균군집의발달은스타터접종의영향을받았으며
, Bacillus
및Pseudomonas
와유연관계가가까운세균군집이퇴비화과정을 이끄는주요미생물임을 확인하였다
.
Key words
□16S rRNA, composting, thermophilic bacteria, T-RFLP
현대사회는음식물쓰레기
,
축산폐수,
하수슬러지등유기성폐기물의양이폭발적으로 증가하고있다
(11).
유기성 폐기물은 미생물에의해유용물질로전환될수있는유기질자원이다.
국내의 대표적인 유기성 폐기물은 음식물류 폐기물이며
, 2007
년 환경부통계에의하면국내생활폐기물중음식물,
채소류의구성비는
28.7%
이다(1).
재활용을 위해 분리배출,
수거하는 남은음식물류 배출량도
2006
년11,463
톤/
일, 2007
년13,754
톤/
일로계속증가하고있다
.
따라서음식물쓰레기등을처리하기위한 효율적인방법에 관심이 증대되고 있으며,
처리방식의 하나로 미생물에의한호기성퇴비화가주목받고있다.
호기성퇴비화 는유기성 폐기물을 비료처럼안정한 물질로만드는 생물학적 전환시스템이다(5).
퇴비화공정의유기성폐기물분해는미생물에의해서주도되 며
,
온도변화에따라미생물의활성이달라진다(22).
온도는퇴비화과정과속도에영향을미치는중요한요인이며
(9),
퇴비공정의목적에따라최적온도는다양하다
(21). 45
oC
이하의중온에서도퇴비화 과정이진행될 수있으나 일반적으로고온미생물 이퇴비화에서최대의활성을갖는온도범위는
50~60
oC
이며(23), 40
oC~60
oC
의고온퇴비화과정에서lignocellulose
와같이분해하기힘든유기물의분해를관찰한보고도있다
(25).
또한,
고온퇴비화과정은미생물의활성을증진시키고병원성미생물을불활
성화시킨다는 장점을 갖고있다
(9, 26).
따라서 가능한 최대의온도에서 퇴비화가 진행되는 것이 낮은온도의 퇴비화과정에 비해효율성과안전성의측면에서유리하다고볼수있다
(19).
퇴비화과정은미생물활성에의한자체발열과정이며온도의
변화에 따라서크게
1)
중저온단계, 2)
고온단계, 3)
온도감소및안정화단계로나뉘어진다
(19).
퇴비화과정은또한유기물과환경조건
(
온도, pH,
함수율등)
의변화에적응하는미생물 군집의연속적인천이과정으로이해될 수있다
(25).
초기에는중온성군집이우점하며이용이 쉬운유기물을분해하고열을발생 시킨다
.
그러나온도가상승함에따라고온성군집으로대체되며 리그닌과같이분해가어려운유기물을이용한다.
온도가하강하는마지막 안정화단계에서는중온성군집이다시형성된다
.
따 라서 각단계에서 우점하는 미생물을 규명하고 미생물군집의 변화를조사하는연구는퇴비공정의효율적인개선과생산된퇴 비의품질제어측면에서매우중요하다.
본연구에서 조사한 음식물쓰레기의 퇴비화 과정은스타터
(starter)
를 폐기물에 식종하였으며,
초고온 단계에서는 온도가95
oC
이상상승하였다.
스타터는일본(Sanyu Co.)
에서수입한제품으로
70
oC~85
oC
에서활발하게생육하는Bacillus
와Geobacillus
외에도배양이어려운다양한초호열균이포함되어있으며
,
이들은
50
oC
이하에서는증식하지않는다고알려져있다(17).
초고온 이라는제한적인환경에서나타나는군집의변화를조사하기위하여
16S rRNA
유전자 분석을수행하였고,
총세균및고온미생물수의변화와병원성지표미생물의존재유무를조사하여효 율적인퇴비화공정의개발에유용한정보를제공하고자한다
.
재료 및 방법
시료 채취정읍시에서수거된음식물쓰레기와스타터
(starter; Sanyu Co., Japan)
를4:5
의비율로 혼합(
총34
톤)
하고실내(5 m
×8 m
×3 m)
에야적하여
2
개월동안퇴비화과정을진행시켰다.
인위적인산소*To whom correspondence should be addressed.
Tel: 82-43-261-3261, Fax: 82-43-264-9600
E-mail: [email protected]
공급및온도조절은없었으며
,
퇴비의온도증가가정체될때마 다셔블로더(shovel loader)
로섞어주기를반복하고깊이1 m
지 점의퇴비시료10 g
이상을총6
회(1
일, 10
일, 16
일, 29
일, 36
일, 56
일)
채취하였다.
퇴비화과정초기부터안정화시기까지온도, pH,
함수율을측정하였다.
총세균수 측정
시료
1 g
을취하여100 ml
의인산완충용액(phosphate buffered saline; KCl 0.2 g/L, KH
2PO
40.2 g/L, NaCl 8 g/L, Na
2HPO
41.14 g/L, pH 7.4)
에넣고5
분간혼합교반하여희석시킨후1 ml
을
polycarbonate membrane filter (pore size 0.2
µm, Millipore, USA)
로 여과하였다. Filter
에 여과된 시료는acridine orange (1 mg/ml) 500
µl
를이용하여 염색시키고 형광현미경(Excitation;
500 nm, Emission; 520 nm, Nikon, Japan)
을 이용하여1,500
배의 배율로미생물수를측정하였다(10).
고온 미생물수 측정
총세균수측정과동일한방법으로인산완충용액에희석된시료
1 ml
를80
oC
항온수조에서1
시간동안방치하여고온미생물을제 외한 다른 미생물을 사멸시켰다.
고온 처리한 시료는polycarbonate membrane filter
에여과한 후Live/Dead BacLight ™ Bacterial Viability kit (Invitrogen, USA)
를 이용하여 염색시키고,
형광현미경
(Excitation: 470 nm, Emission: 540 nm, Nikon)
을이용하여
1,500
배의배율로고온미생물수를측정하였다(3).
대장균 및 살모넬라 측정
대장균과살모넬라측정은수질오염공정시험방법
(2)
에제시된 최적확수법을이용하였다.
총세균수측정과동일한방법으로인 산완충용액에 희석된 시료를 라우릴 트립토스 부이온 배지(Difco ™ Lauryl Tryptose broth)
에접종한 후, 37
oC
에서24
±2
시 간배양하였다.
배양후가스발생이관찰된양성시험관의수와최적확수표로검체
1 g
중의대장균수를 산출하였다.
살모넬라측 정을위해셀레나이트 배지(Difco ™ Selenite broth)
에희석시료를접종하여
37
oC
에서18
∼24
시간배양하였다.
모든증균배지에 서 배양액을 한 백금이씩 취하여 비스무스 한천 선택배지(Difco ™ Bismuth Sulfite agar)
에 선상도말 하고37
oC
에서24~48
시간배양한후에살모넬라양성결과를판정하였고,
최적확수표를이용하여살모넬라수를산출하였다
.
분자생물학적 세균 군집분석 1) T-RFLP 분석
Miller
등(15)
의bead beating
방법으로시료0.2 g
에서핵산을직 접 추출하였다.
추출된 핵산은QIAamp DNA Micro kit
(QIAGEN, Germany)
로정제하였다.
시료속에존재하는세균군집의
16S rRNA
유전자를증폭하기위해eubacterial primer
인27F
에형광물질
6-carboxy-fluorescine (FAM)
을표식시킨27FF ( E. coli numbering 8-27; 5’-AGAGTTTGATCMTGGCTCAG-3’)
와1492R
( E. coli numbering 1492-1510; 5’-GGYTACCTTGTTACGACTT- 3’)
을사용하였다(14). PCR
반응은Core Bio System (Korea)
의Taq polymerase
와 시약을 사용하였으며, 95
oC
에서3
분간 초기 열처리한 후, 95
oC
에서30
초, 49
oC
에서30
초, 72
oC
에서1
분씩30
회반복하고, 72
oC
에서10
분동안반응하였다. PCR
증폭산물 은GENEALL ™ PCR SV Mini kit (General Biosystem, Korea)
로 정제한 후에 제한효소
Alu I (Quantum Biotechnologies, Canada) 5 unit
을첨가하여37
oC
에서5
시간이상반응시켰다.
제 한효소에 의해절단된 핵산단편을polyacrylamide gel
에서 전기영동
(ABI PRISM
®3100 Genetic Analyzer; Applied Biosystems, USA)
하여말단단편(T-RF; terminal restriction fragment)
의profile
을 확인하였다
. GelCompar II
프로그램(Applied Maths, Belgium)
를이용하여각시료에서확인된T-RF profile
의피어슨(Pearson)
상관계수를 계산하였고, UPGMA (Unweighted Pair Group Method with Arithmetic mean)
방법으로집괴분석(Cluster analysis)
을수행하였다.
2) 16S rRNA 유전자의 염기서열 분석
27F
와1492R
로 증폭한PCR
산물을pGEM-T vector (Promega, USA)
를이용하여E. coli DH10B
에형질전환시킨뒤시료당
100
개이상의재조합클론을선별하였다. T-vector
의염 기서열에상보적인prGTf (5’-TACGACTCACTATAGGGCGA-3’)
와
16S rRNA
의1492R primer
쌍을 사용하여direct amplified
PCR
을수행하고(4),
전기영동으로PCR
산물의크기를확인하여5 ’에서 3 ’ 방향으로삽입된재조합클론을 2
차선별하였다.
선별
2
차선별하였다.
선별된 재조합 클론들은
T7 (5’-TAATACGACTCACTATAGGG-3’)
primer
를 이용하여ABI 3730XL DNA Analyzer (Applied
Biosystems, USA)
로염기서열을분석하였다.
주요균주의염기서열은
DDBJ (DNA Data Bank of Japan; http://www.ddbj.nig.ac.
jp)
에등록하였다(accession no.: AB507770-AB507797).
3) 16S rRNA 염기서열의 계통분류학적 분석
시료에서 확인된
16S rRNA
유전자의염기서열과Ribosomal
Database Project (RDP; http://rdp.cme.msu.edu), GenBank (http://
ncbi.nlm.nih.gov)
의database
로부터확인된염기서열을CLUSTAL X (version 1.83)
프로그램을이용하여정렬하였다(24).
정렬된염 기서열은PHYLIP package (version 3.6a3)
를이용하여Jukes and Cantor distance model (12)
과neighbor-joining method (20)
로염기 서열간의진화적거리와계통도를추론하였다(8).
또한, bootstrap
값은
1,000
회의resampled data
로부터계산하였다(7).
결 과
온도 변화
퇴비화과정중의온도변화를 관측한결과
,
약2
oC
에서퇴비 화가시작되어6
일후급격히 온도가상승하여8
일에는95
oC
까지증가하였다
(Fig. 1).
그후80
oC
수준으로온도가 감소하였을 때 퇴비를 섞어주었고, 2
일 동안의 정체기를 보인 후 온도가90
oC
이상으로다시증가하였다.
고온퇴비섞어주기작업은총6
회실시하였으며
,
최고온도는26
일에97
oC
이었다.
퇴비화 과정은
60
oC
를기준으로온도상승단계(1
단계; 1~6
일),
초고온단계(2
단계; 7~43
일),
안정화단계(3
단계; 44~56
일)
의3
단계로구분하 였다.
퇴비화과정동안고온으로인한수분증발로시료의함수 율은초기40%
에서26%
까지지속적으로 감소했다. pH
범위는7.5~7.9
로약알칼리성이었으며,
초기에는다소감소하였으나29
일이후에는증가하였다
.
총세균수와 고온 미생물수
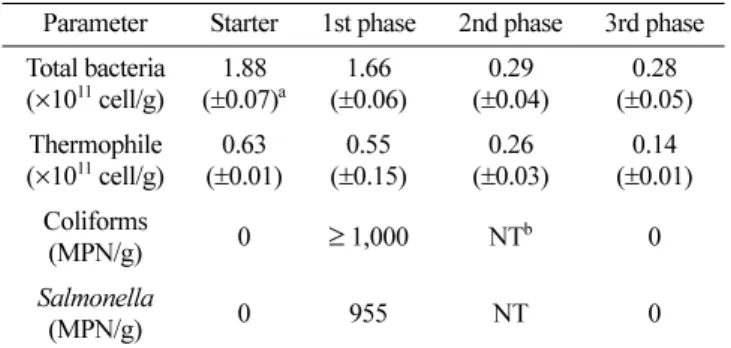
스타터
(starter)
의총세균수와음식물쓰레기와혼합한1
단계1
일 시료의 총세균수는 각각
1.88
×10
11cell/g (dry-weight), 1.66
×10
11cell/g
이었다(Table 1).
그러나초고온퇴비화가진행된2
단계29
일시료에서는0.29
×10
11cell/g
으로1
단계의18%
이하 로감소하였고, 3
단계56
일시료에서도0.28
×10
11cell/g
으로낮은 분포를나타내었다.
고온미생물수도1
단계의0.55
×10
11cell/g
에 서2
단계의0.26
×10
11cell/g, 3
단계에서는0.14
×10
11cell/g
로측정되었다
.
퇴비화진행에따른총세균의급격한감소로인하여, 1
단계에서총세균수의
33%
를차지하던고온미생물의비율은2
단계가되며
89%
로증가하였고, 3
단계에서는50%
를차지하였다.
대장균 및 살모넬라 측정
스타터에서는대장균과살모넬라가검출되지않았으나
,
음식물 쓰레기가 혼합된1
단계 시료에서는1,000 MPN/g (dry-weight)
이상의대장균과
955 MPN/g
의살모넬라가검출되었다(Table 1).
그러나높은온도에노출된후의
3
단계시료에서는대장균과살모넬라가검출되지않았다
.
세균군집 분석
1) T-RF profile의 집괴분석
각단계퇴비시료의
T-RF profile
로부터계산된피어슨상관계수를이용하여집괴분석을한결과
,
온도변화에따라구분한 퇴비진행단계별로구분되는클러스터가확인되었다(Fig. 2). 1
단계시료는음식물쓰레기에포함된세균군집때문에스타터와 구분되는클러스터를나타내었으나
,
퇴비화가진행됨에따라10
일
, 16
일의2
단계 시료는 스타터와90%
이상의 유사도를나타내었다
.
퇴비온도가62.5
oC
이하로감소한36
일시료부터유사 도가낮아졌으며,
안정화단계의시료는독립된클러스터를형성하였다
.
따라서1
단계 시기와2
단계후반부터3
단계의 기간중 세균군집의변동이컸음을알수있다.
2) 16S rRNA 유전자를 이용한 군집 분석
16S rRNA gene
의염기서열을분석하고유사도가높은균주를확인함으로써 군집구조를조사하였다
(Fig. 3).
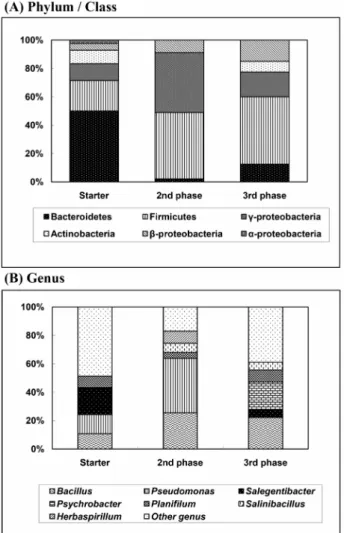
시료당40~45
개 클론의염기서열을분석한결과,
스타터의경우Bacteroidetes
문 이50.0%, Firmicute
문은21.4%,
γ-proteobacteria
강이11.9%
를Fig. 1. Variation of physicochemical parameters in the composting process. The arrow indicates sampling days. Filled circle ( ), empty circle ( ), and filled triangle ( ) represent temperature, moisture contents, and pH value, respectively.
Table 1. Numbers of total bacteria, thermophilic bacteria, Coliforms, and Salmonella in the composting process
Parameter Starter 1st phase 2nd phase 3rd phase Total bacteria
(
×10
11cell/g) 1.88
(
±0.07)
a1.66
(
±0.06) 0.29
(
±0.04) 0.28 (
±0.05) Thermophile
(
×10
11cell/g) 0.63
(
±0.01) 0.55
(
±0.15) 0.26
(
±0.03) 0.14 (
±0.01) Coliforms
(MPN/g) 0
≥1,000 NT
b0
Salmonella
(MPN/g) 0 955 NT 0
a
The numbers in a round bracket represent standard error
b
NT; not tested
차지하였다
.
속(genus)
수준에서는Bacteroidetes
문에 속하는Salegentibacter
가전체의16.6%,
γ-proteobacteria
강의Pseudomonas
가
11.9%, Firmicutes
문의Bacillus
가9.5%
를차지하였다.
그외에도
Bacteroidetes
문의Flavobacterium , Aequorivita , Firmicute
문의Planifilum
이각각7.14%
를차지하는등다양한미생물군집이확 인되었다.
음식물 쓰레기퇴비화 과정중가장높은온도가 관찰되었던
2
단계29
일시료의염기서열분석결과,
스타터시료에서9~11%
수준이었던
Bacillus
와Pseudomonas
속이각각26.6%, 40.0%
로 증가함에 따라21.4%
를 차지하던Firmicute
문이46.6%
로, 11.9%
였던 γ-proteobacteria
강은42.2%
로증가하였다.
스타터시 료에서4.7%
였던 β-proteobacteria
도8.8%
로증가하였다.
그러나스타터 시료에서
50.0%
로가장 우점하였던Bacteroidetes
문은2.2%
수준으로급격히줄어든것을확인할수있었다.
안정화 단계인
3
단계 시료에서는Firmicutes
가47.5%,
γ- proteobacteria
가17.5%
를차지하였으며,
그밖에도β-proteobacteria 15.0%, Bacteroidetes 12.5%, Actinobacteria 7.5%
로 나타났다.
속수준에서는
Firmicutes
에속하는Bacillus
가전체의20%
로우점하 였고,
γ-proteobacteria
인Psychrobacter
가각각전체의17.5%
를차지하였다
.
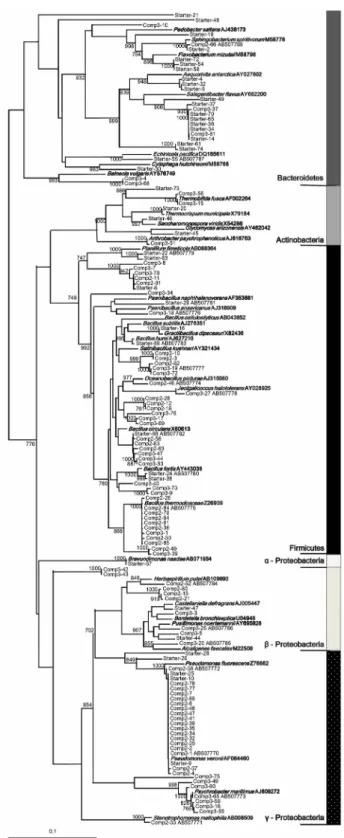
3) 계통분류학적 분석
각단계의 시료에서 확인된
16S rRNA
유전자염기서열들을GenBank (http://ncbi.nlm.nih.gov)
와RDP (http://rdp.cme.msu.edu)
의
database
서열과 비교 분석하였다(Fig. 4).
스타터시료에서16.6%
를 차지하며 우점하였던 균주는Bacteroidetes
문의Salegentibacter
속인S. flavus (AY682200)
와92~93%
의유사도를 나타내었다.
또한Firmicutes
문Bacillus
속의B. fortis (AY443038), B. circulans (X60613), B. humi (AJ627210)
와,
γ- proteobacteria
강Pseudomonas
속의P. veronii (AF064460), P.
fluorescens (Z7662)
와유사균주들이확인되었다. 2
단계시료에서40.0%
를 차지한Pseudomonas
속은Pseudomonas veronii (AF064460)
의16S rRNA
유전자와98%
이상의유사도를나타내 었다.
또한2
단계에서26.6%
를차지한Bacillus
속은B. circulans (X60613), B. thermocloacae (Z26939)
유사균주로 확인되었으며,
각각전체의
6.6%, 20.0%
를차지하였다. 3
단계시료의17.5%
를 차지한 γ-proteobacteria
강의Psychrobacter
속은P. faecalis (AJ421528), P. pulmonis (AJ437696), P. martimus (AJ609272)
유Fig. 2. Cluster analysis on Alu I T-RF profiles of the composting reactor. The dendrogram was constructed by Pearson’s correlation coefficient and UPGMA method. Numbers on the branch represent the similarity value of the samples belonging to the same cluster.
Fig. 3. Variation of bacterial communities in the composting reactor.
The proportions of phylum or class (A) and genus (B) were calculated
from the 16S rRNA gene clone libraries.
사균주로확인되었다
.
고 찰
퇴비화과정에 있어 온도는미생물 군집의천이를 유도하는 가장 중요한 인자로 작용하며
,
높은 온도에서진행되는퇴비화 과정은분해가어려운다양한물질을빠르게분해할수있는장점이 있다
(9).
본연구에서 측정된 퇴비화 과정의 온도는,
외부기온이 낮은
12~2
월에진행되었음에도불구하고, 2
oC
에서97
oC
까지상승하였으며일반적으로보고된퇴비의최고온도를초과 하는특징을나타내었다
.
또한온도의변화를기준으로구분한3
단계의 퇴비화과정에서미생물 군집의변화와 함께총세균및 고온미생물의분포변화가관찰되었다
.
16S rRNA gene clone library
에서확인된 균주들을속수준으 로구분하고세균군집다양성 지수(H; Shannon-Weiner diversity index)
와우점도(D; Simpson dominance index)
를 계산하였을때 스타터에서는H=4.07, D=0.07
이었으나초고온 단계의29
일시료에서는
H=2.43, D=0.25
로특정 군집의우점이 진행되었음을나타낸다
.
퇴비화 과정이 완료된 생산물의일부를 다음퇴비화 공정의스타터로공급하는상황을고려하면,
안정화단계를거친스타터에는중온성군집이다수포함되어있으며음식물쓰레기 에포함된군집과함께초기유기물분해와열발생에기여하였 으나변화된환경에서는고온세균의성장이촉진되었기때문이
라고생각된다
(9). T-RF profile
의비교분석에서도세균군집의연속적인 천이를 확인할 수있으며
,
스타터와16
일까지의시료는90.2%
이상의 유사도를나타내었으나29
일시료는87.9%
로낮아졌다
(Fig. 2).
특히음식물쓰레기와혼합한첫날의 시료가2
단계의 시료들과 스타터보다 낮은 유사도
(57.8%)
를나타낸 것은 고온발효에스타터 군집이중요함을암시한다. Pseudomonas
및Bacillus
속과 유연관계가 가까운균주들이 스타터 시료에서는각각
11.9%, 9.5%
로낮은비율을나타내었지만2
단계시료에서는
40.0%, 26.6%
로 증가하였으며, Pseudomonas veronii
와Bacillus thermocloacae
의증가가2
단계의세균군집에많은영향을끼친것으로보인다
.
Pseudomonas veronii
는4
oC~36
oC
의중저온에서생장하고오염된토양에서방향족화합물을분해하는특징을 가지고있는것 으로보고되었다
(16). 2
단계에서전체군집의40.0%
를차지한균 주는P. veronii
와16S rRNA
유전자의염기서열이98%
이상의유사도를나타내어동일종의유사도기준인
97%
보다크지만,
초고온환경에서 성장이가능한특징을 고려하면 분리 배양후
genomic DNA hybridization
실험 등분류학적실험을 수행해야 정확한동정이가능할것이다.
또한,
이균주의최적생장환경조건및우점을 유도한 환경요인의변화에대한 연구가진행된다 면고온퇴비화공정의효율적인제어에도움이될것이다
.
Bacillus thermocloacae (Z26939)
와98%
이상의유사도를갖는 균주는2
단계에서20.0%
를차지하였으나3
단계에서5.0%
로감소하였다
. B. thermocloacae
는 고온,
호기성 조건으로sludge
를 처리하는과정에서분리되었다고보고되었다(6).
Fig. 4. Phylogenetic tree based on the partial sequence of 16S rRNA genes cloned from the composting reactor. Prefixes “Starter”,
“Comp2”, and “Comp3” in clone name represent the 1st phase, 2nd
phase, and 3rd phase, respectively. The tree was constructed by using
the neighbor-joining method. The sequence of Methanothermobacter
defluvii (X99046) was used as an outgroup. The names of type strains
are bold faced. The bootstrap values above 70 % are shown for
internal nodes.
Pedro
등(18)
은농,
산업폐기물을이용하여현장수준으로진 행한 퇴비화의 고온 단계(60
oC~75
oC)
에서B. thermocloacae
와 유연관계가가까운균주가우점하는것을확인하였으며,
이세균 으로인해퇴비화과정이높은온도에서진행되었다고보고하였 다.
또한 고온처리를 한 퇴비화 과정에서Bacillus , Filobacillus , Planifilum
이우점한 사례도 보고되었다(27).
따라서2
단계 과정 에서다수발견된B. thermocloacae
유사균주도 음식물쓰레기 의퇴비화과정의 온도상승에중요한 역할을하였으며세균군 집의천이를유도하였다고생각된다.
온도가
50
oC
∼89
oC
수준으로 감소하는3
단계에서는2
단계의 우점종인Bacillus
와Pseudomonas
균주의비율이급격히감소하 고, 5
oC~37
oC
의성장범위를 갖는다고보고(13)
된 중저온성세 균Psychrobacter
속의균주가증가하는것을확인할수있었다.
선행공정의생산물인스타터의세균군집과안정화과정을거 친
3
단계56
일시료의 세균군집에 상당한 차이가 있음은T-RF
profile
과16S rRNA
염기서열 결과를 비교하면 알 수 있다.
Salegentibacter , Thermobifida , Bacillus
등은 스타터와3
단계 시 료에서함께확인되었고 스타터시료와 유사하게2
단계를 거친후
Bacteroidetes
문이증가하였다는공통적인특징이관찰되었지만
,
생산된퇴비의 일부를다음공정의 스타터로사용하는현재방법은퇴비화 공정이 반복하여진행될수록스타터의품질관리 가어려울것으로 예상된다
.
생산된퇴비의군집이 스타터의군집과차이가있으면
,
공정이반복됨에따라스타터의군집조성 변화가커지며주요고온균주가감소할가능성이있기때문이다.
따라서고온 퇴비화 과정에주요한세균군집을 고온 시료에서 확보하여스타터로사용하는방안이퇴비공정의효율적인관리 및시간단축에도움이될것이다
.
Ryckeboer
등(19)
은일반적인퇴비화과정에서총세균수는퇴비화가진행되는 동안 점점 증가하여 고온 단계에서 최고치를 이루고
,
온도가떨어지며안정화단계에진입하면서점점감소한 다고보고하였다.
본연구에서는2
단계에서퇴비의온도가최고치인
97
oC
까지 증가하면서1
단계에 비해17.4%
수준으로 총세 균수가급격히감소하였다.
그러나고온미생물의분포는1
단계에서 총세균의
33.1%
였으나, 2
단계에서는0.26
×10
11cell/g
으로 총세균의89.6%
수준으로오히려 증가하였다.
따라서97
oC
까지온도의상승으로인하여중온성세균의성장이저해되는환경적 압박이
60
oC
내외에서진행되는일반적인퇴비화과정보다컸으 며2
단계의 세균군집의대부분을고온성세균이 점유하는결과 를초래하였다고생각된다.
우리나라는퇴비의병원성지표미생물의검출에대한법규가 미비한 실정이다
.
또한 퇴비가 충분히 발효되지않으면 퇴비를 이용하여경작된 채소가 병원성 세균에 오염될 가능성이 있다.
일반적으로높은온도에서퇴비화를진행시키는 것이병원균을 사멸하는데효과적이라고 알려져 있다
(19). Wiley
와Westerberg (26)
는퇴비화과정의온도가60
oC~70
oC
이상이되면병원성지 표세균이사라진다고보고하였다.
본퇴비화과정은높은온도에서진행되었기때문에
,
음식물쓰레기의혼합으로인하여1
단계에서
1,000 MPN/g
이상으로많이 분포하였던대장균이나 살모넬라와같은병원성지표세균들이
3
단계에서는검출되지않았다.
16S rRNA
염기서열을이용한군집분석결과에서도병원성지표세균은 확인할 수없었다
.
퇴비 발효과정에서효율적인병원성 세균의 제거는안전한 퇴비생산에필수적인요소이며본퇴비 화공정에서는높은온도로인하여병원성세균이사멸됨으로써 생산된퇴비제품의안전성을높였다고할수있다.
자체발열반응에의하여온도가빠른속도로상승하는음식물 쓰레기의 퇴비화 과정에서미생물 군집의 변화를 관찰한 결과
,
퇴비화단계에따른세균군집의천이과정과초고온단계에서중 요한역할을할것이라예상되는미생물의우점을확인할수있 었다
.
또한스타터의중요성과고온으로인한병원성세균의사멸효과도 확인할수있었다
.
우수한퇴비를경제적으로만들기 위해서는퇴비공정의효율적인제어가필요하므로,
이번연구에 서확인된 초고온미생물을분리하여생리,
생화학적특성을조 사하고스타터를개발하여퇴비공정에이용한다면음식물쓰레 기의자원화에기여할수있을것이다.
감사의 말
이논문은
2007
년도 충북대학교학술연구지원사업의 연구비지원에의하여연구되었음