서 론
어류뿐 아니라 대부분의 동물들은 규칙적인 생식 주 기를 가지고 있는데, 송사리(박과 최, 1998)와 버들붕어 (송과 최, 2000) 등과 같이 적정한 광 조건과 수온 하에 서 짧은 산란주기를 가지는 어류가 있는 반면, fresh- water eels (Anguilla sp.)과 같이 10~14년의 긴 성숙기 간을 가지고 있는 것까지 다양하다(Lagler et al., 1962).
그러나 온대지방에 서식하는 대부분의 담수어류는 환경 적인 변화 즉, 온도와 광 조건의 계절적 변화가 호르몬 분비에 영향을 주며 어류의 생리적인 상태와 행동을 변
화시켜, 일정한 계절적 산란주기(spawning cycle)를 가 지고 산란을 하게 된다(de Vlaming, 1972). 산란시기 (spawning season)는 어류의 생리적 리듬과 함께, 난 발 생 및 부화한 개체군이 번성할 수 있는 적절한 수온, 자 원이용의 용이성, 포식자와의 관계 등이 복합적으로 작 용하여 가장 유리한 시기에 산란 할 수 있게 적응하여 진화하였다. 성 성숙과 산란시기의 연구는 생식능력을 획득하여 산란에 참여할 수 있는 연령과 크기를 추정하 고, 산란장의 환경조건, 생산력 등의 특성을 파악할 수 있어, 이를 토대로 대상 어류의 생활사와 생태를 이해하 는데 도움이 된다.
잉어과(Cyprinidae)에 속하는 납자루아과(Acheilog-
─
─ 278 ──
홍천강 상류에 서식하는 묵납자루
,
Acheilognathus signifer
의 성 성숙과 산란시기백 현 민∙송 호 복∙권 오 길 강원대학교 자연과학대학 생물학과
Sexual Maturation and the Spawning Season of the Korean Bitterling, Acheilognathus signifer in Upper Reachs
of the Hongcheon River
Hyun-Min Baek, Ho-Bok Song and Oh-Kil Kwon
Department of Biology, Kangwon National University, 200-701 Chunchon, Korea
The Korean bitterling, Acheilognathus signifer, collected in the Naechoncheon, Hongchun- gun, Kangwon-do, Korea, from March to October 2002, were examined for sexual maturation and spawning season. Gonadosomatic index (GSI) and ovipositor indices (OI) values reached the maximum in May. The body length at sexual maturity was estimated at 45 mm for the females and 40 mm for the males. Reproductive cycle can be classified into the growing stage (September to October), mature stage (March to April), ripe and spent stage (May to June), and degenerative and resting stage (July to August), Two-year-old populations were the highest participants in spawning. Fecundity of A. signifer ranged from about 141~314 (average 225) eggs. The sex ratio of female:male was not significantly different from 1 : 1 (x2 = 13.5, P⁄0.05).
Key words : Acheilognathus signifer, sexual maturation, spawning, reproductive cycle, sex ratio
nathinae)는 전 세계에 약 40여종이 분포하고 있는데, 본 아과의 대부분의 종들은 우리나라, 중국, 일본, 대만, 베트남 등의 동아시아에 주로 서식하고, 유럽에는 납줄 개, Rhodeus sericeus 1종만이 분포하며(Holcik, 1962;
Holcik and Duyvene, 1962; Schimidt, 1982; 송, 1994), 우 리나라의 남한에는 3속 14종이 분포하는 것으로 알려져 있다(김과 박, 2002). 본 아과의 암컷은 길게 신장된 산 란관(ovipositor)을 이용하여 조개의 출수공을 통해 조 개의 새강(gill cavity)에 산란하고(Wiepkema, 1964), 조 개는 산란하기 위해 가까이 접근하는 본 아과 어류의 아가미나 지느러미 등에 유생(glochidium)을 부착시켜, 유패를 멀리 분산시키는 특이한 공생관계에 있어(권, 1981), 흥미 있는 생태학적 의의를 갖는다.
묵납자루, Acheilognathus signifer는 한국 고유종으로, 한강수계를 포함한 이북지역인 임진강, 대동강, 압록강, 성천 및 회양 등에 분포하며(內 , 1939), 같은 아과 (subfamily)의 다른 종보다 비교적 상류에 서식하고, 작 은말조개, Unio douglasiae sinuolatus에 산란한다. 납자 루아과의 산란주기에 관한 연구는 줄납자루, Acheilog- nathus yamatsutae의 생식주기(송과 권, 1989, 1994)와 각시붕어, Rhodeus uyekii의 산란주기에 관한 연구(안, 1995)가 있으며, 본 종에 관한 연구로는 內 (1939)가 성어의 난소내 완숙란의 상태와 후기자어의 출현상황에 의하여, 산란기를 추정한 단편적인 기록 이외에 산란시 기 및 생식주기에 관한 연구는 찾아 볼 수 없었다.
본 연구는A. signifer의 생식 생태학적 연구의 일환으 로, 성적 성숙 및 산란시기를 밝히고, 최근 난 개발과 오 염에 의해 개체수가 급격히 감소하고 있는 본 종의 보 호와 보전을 위한 생식 생태학적 기초 자료를 확보하고 자 하였다.
재료 및 방법
본 연구는 2002년 3월부터 10월까지 강원도 홍천군 의 홍천강 지류인 내촌천에서 매월 1회 채집을 실시하 였다(Fig. 1). 어류의 채집은 투망(망목 5×5 mm), 족대 (망목 2×2 mm)와 유인망(25×25×70 cm, 망목 3×3 mm)을 이용하였으며, 채집된 표본은 현장에서 10% for- malin에 고정한 후 실험실로 운반하였다. 채집된 표본은 성별로 구분하고, 체장(body length : BL)은 어체측정판 을 사용하였고, 산란관의 길이(ovipositor length : OL)는 1/20 mm vernier caliper를 이용하여 0.1 mm 까지 측정 하고, 산란관 길이지수(ovipositor index (%) : OI = OL/BL
×100)를 구하였으며, 체중(body weight: W)은 전자저울
을 이용하여 0.01 g까지 측정하였다. 생식소중량지수 (gonadosomatic index (%) : GSI = Gw/W×100)는, 암컷 201개체, 수컷 154개체를 대상으로 복부를 절개하여 생 식소를 분리하고 무게(Gw)를 측정하여 월별로 산출하 였다.
생식소의 조직학적인 변화를 조사하기 위해 적출한 생 식소를10% 중성formalin에 고정한 후 paraffin으로 포 매하여 5~6µm의 두께로 연속 절편하고, haematoxylin
-eosin으로 이중염색하여 광학현미경 하에서 관찰하였
다. 포란수는 해부현미경하에서 계수하고, 난경은 광학현 미경하에서 마이크로메타로 장경을 계측하였다.
통계학적 처리는 systat v. 10을 이용하여 분산분석 및 회귀분석을 하였고, 회귀분석에서 체장과 포란수는 로그 변수로 변환하여 분석하였다.
결 과
1
.월별 생식소중량지수
(GSI)변화
생식소중량지수(GSI)의 월별 변화 양상을 보면, 3~5 월에 걸쳐 암컷은6 이상, 수컷은 2 이상의 높은 값을 유 지하였다. 4월에 암컷(GSI = 6.95±0.83)과 수컷(GSI = 2.84±0.55) 모두 최대치를 나타내었고, 5월부터 서서히 감소하기 시작하여 6월에는 급격히 하강하였다. 그 이후 에 지속적으로 감소하여 암컷은 8월(GSI = 2.02±0.91), 수컷은 9월(GSI = 0.54±0.50)에 최저값을 나타냈으며, 10월부터 다시 GSI가 상승하였다. 수온 및 일장, GSI의 변화를 연계해 보면, 3월에 7.5�C, 12.1 L, 4월에 14.3�C, 13.2 L로 GSI는 수온이 상승하고, 일장이 길어짐에 따라 GSI가 높아졌고, 5월부터 GSI는 낮아지게 되는데 이때 Fig. 1. Map showing the sampling sites at Naechoncheon in Hongcheon-gun, Kangwon-do.
Kangwon-do 37�50′
37�49′
37�48′
128�03′
0 500m 2km
128�04′ 128�05′ 128�06′ 128�07′
Hongcheon-gun
Naechon-myeon
KOREA Hongcheon-
gun
일장은 14.2 L, 20.4�C이었다. 6월에 이르면 일장이 최대 로 길어져 14.3 L, 수온은 26�C까지 상승하지만 GSI는 암컷, 3.77±1.23, 수컷은 0.77±0.38로 낮은 값을 나타 내었다. 그 후에 일장, 수온 및 GSI는 계속 낮아지다가 일장이 11L, 수온이 13�C 이하가 되는 10월부터 암컷과 수컷의GSI는 다시 높아지기 시작하였다(Figs. 2, 3).
2
.산란관의 길이변화
A. signifer가 산란을 하기 위해서는 산란관의 신장이
필수적이며, 산란 성기일 때 최대로 신장된다. 월별 산란 관길이 지수(OI)의 변화를 조사한 결과, 3월부터 증가하 기 시작하여 5월에 이르면 산란관이 최대로 신장(26.53
±8.34%)된다. 6월부터 산란관의 길이는 축소되기 시작 하여 8월에 이르러 가장 축소(5.77±1.37%)되었고, 그 후에는 서서히 증가하기 시작하였다. 따라서 본 종의 산
란은, GSI가 4월에 가장 높은 값을 나타낸 후 5월부터 감소하기 시작하고, 산란관이 최대로 신장되는 시기가5 월인 것으로 보아, 이 시기가 산란성기인 것으로 추정된 다(Fig. 4).
3
.연령과 체장에 따른 성숙도 및 성비
A. signifer의 주 산란기로 추정되는 4~5월에 채집된 개체를 대상으로 체장별 GSI의 관계를 살펴 본 결과, 암컷은 체장 35 mm 전 후에서 성숙하기 시작하여 체장
45 mm 이상에 이르면 모든 개체에서 높은 성숙도를 보
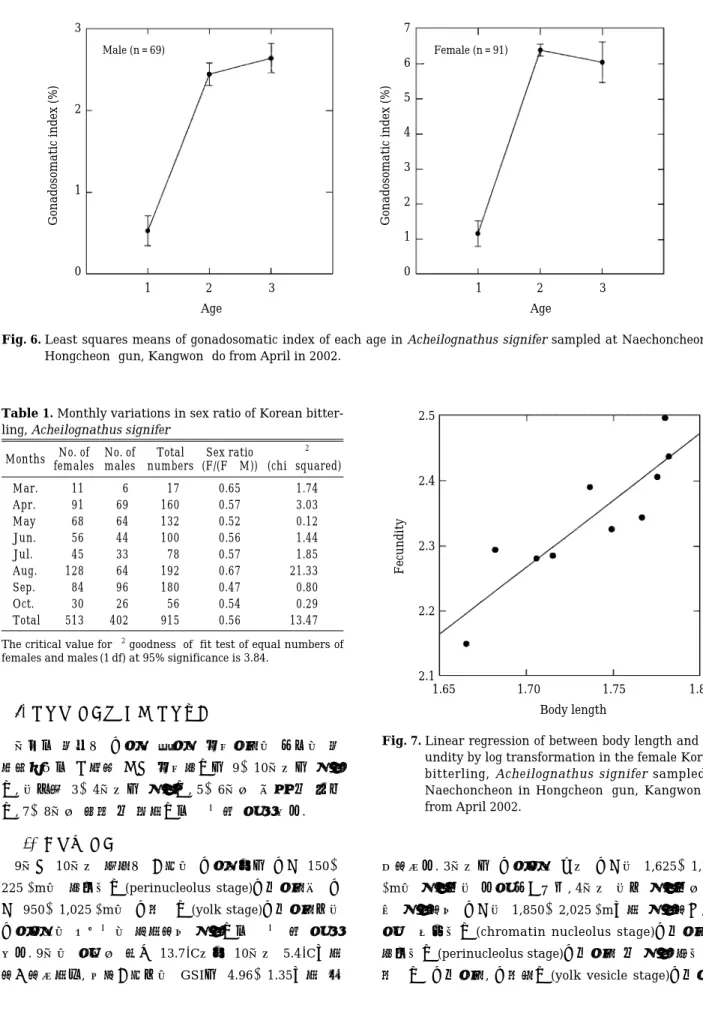
여 대부분의 개체가 생식이 가능 할 것으로 추정된다. 따라서 45 mm 이하의 개체는 미성숙군, 45 mm 이상의 개체들은 성숙군으로 구분할 수 있었으며, 산란이 가능 한 최저 체장은 약 35 mm 전 후로 추정되었다. 수컷도 암컷과 비슷한 양상으로 체장 30 mm에 이르러 생식소 성숙도가 증가하기 시작하여 40 mm에 달하면 생식이 가능한 것으로 추정되는 바, 체장 40 mm 이하의 개체는 미성숙군 그 이상의 개체들은 성숙군으로 구분 할 수 있었으며, 생식소성숙도지수가 2 이상이면 생식이 가능 할 것으로 추정된다(Fig. 5). 백 등(2002)은 체장40 mm 전후는 만 1년생, 40~60 mm 는 만 2년생, 60 mm 이상 의 크기는 만 3년생 이상으로 구분 한 바 있는데, 산란 이 가능 할 것으로 생각되는 높은 생식소성숙도지수를 나타내는 개체들은 2년생의 비율이 가장 높게 나타났다 (암컷: 75.8%, 수컷: 39.1%). 그러나 3년생의 비율은 수컷 (27.5%)이 암컷(6.6%)보다 높게 나타나 주목되었으며, 2 년생 이상에서 암수 모두 연령군에 따른 성숙도의 차이 는 없는 것으로 나타났다(Fig. 6, 암컷; F = 86.1, P⁄
0.001, 수컷; F = 44.0, P⁄0.001).
Fig. 2. Monthly changes of water temperature and day length at Naechoncheon in Hongcheon-gun, Kang- won-do from March to November, 2002.
Fig. 3. Monthly variations of the gonadosomatic index of the female (○) and male (●) Korean bitterling, Acheilognathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from March to October, 2002.
Fig. 4. Monthly variations of the ovipositor index in the female Korean bitterling, Acheilognathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from March to October, 2002.
M A M J J A S O N
Month
Water temp.
Day length 30
25 20 15 10 5 0
16
14
12
10
8
Temperature (�C) Day length (hr)
M A M J J A S O
Month 10
8 6 4 2 0
Gonadosomatic index (%)
M A M J J A S O
Month 40
30
20
10
0
Ovipositor index (%)
한편 생식의 참여 형태가 암컷과 수컷에서 다르게 나 타났는데, 암컷의 경우 성숙한 개체들은 전장의 크기에 큰 영향을 받지 않고, 작더라도 산란관의 길이가 10 AU 이상이면 수컷의 선택을 받아 산란에 참여하게 되어, 산 란관의 길이가 산란 참여에 중요한 요인으로 작용한다. 그러나 수컷은 작은말조개를 중심으로 세력권을 형성하 고, 성숙한 암컷을 조개로 이끌어 산란을 하게 되는데, 만 1년생군은 2년생 이상의 군보다 크기가 작기 때문에
독자적인 세력권 형성이 어렵다. 따라서 만 1년생 수컷 은 주로 다른 쌍이 산란행동을 할 때 산란 직전 또는 직후에 끼어들어 방정하는 sneaker형태로 산란에 참여 하는 것으로 관찰되었다.
또한 외형적으로 암, 수의 구별이 가능한 913개체를 대상으로 성비를 조사한 결과 암컷이 513개체, 수컷이 402개체로, 성비에서 암컷이 수컷보다 우세하게 나타나 암수의 성비가 1 : 1이 아니었고, 암수의 성비는 유의한 차이를 보였다(x2 = 13.5, P⁄0.05).
4
.포란수
A. signifer의 난은 비점착성으로 미성숙난은 구형이며
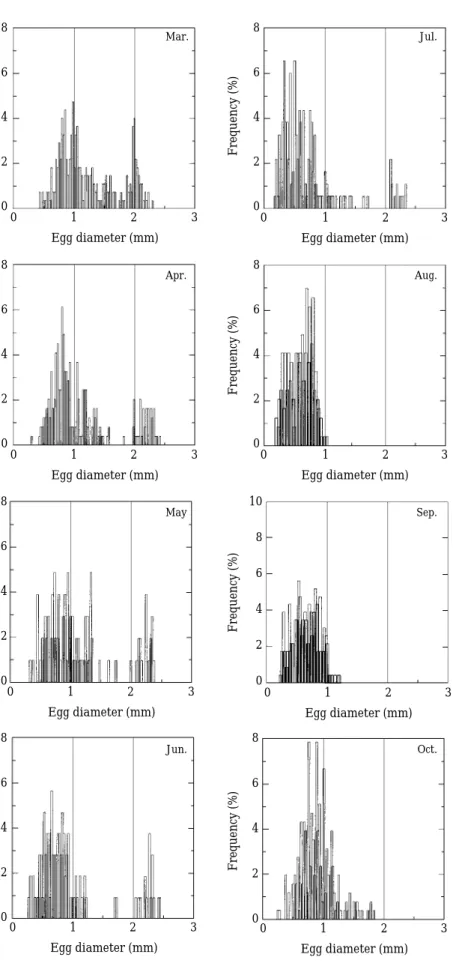
크기가 작으나, 성숙난은 동물극쪽이 볼록하게 부푼 모 양이며, 진한 황색을 띠고, 난의 크기가 2 mm 이상으로 미성숙난과 잘 구별된다. 체장 50 mm 이상의 암컷에서 성숙난의 크기를 측정한 결과 장경은 2.19±0.114 mm, 단경은 1.85±0.080 mm이었다. 또한 3~4월에 GSI값이 6 이상인 10개체를 대상으로 포란수를 조사한 결과 평 균 225개의 난을 가지고 있었다(Table 1). 체장과 포란 수를 로그 변수 변환하여 회귀분석한 결과 Fecundity = 10-1.245±102.066log*Body Length,
(r = 0.89)에 회귀되었고(Fig.
7), 체장이 증가할수록 포란수도 증가하였다(F = 28.9, P
= 0.001). 월별로 체장 50 mm 이상의 암컷을 무작위로 추출하여 포란수를 조사한 결과3~4월에는 194±35~ 209±22개로 나타나, 포란수가 많았고, 5월부터 포란수 가 감소하기 시작하여, 7월에 가장 적었다. 그러나 포란 수는 9월부터 급격히 증가하기 시작하여 10월에는 190
±10개까지 증가하였다(Fig. 8). 이러한 포란수의 월변화 양상은 GSI 및 산란관의 길이 변화와 비슷한 양상으로 나타났다. 송과 권(1989)은 줄납자루의 경우 3~5월에 최소289개, 최대 514개로 평균 302개의 난을 가진다고 보고하였는데, 묵납자루는 최대314개의 난을 가지고 있 었고, 평균225개 내외로 이보다 적었다(Table 2).
5
.난경조성의 월변화
난소내 난들의 월별 크기 변화를 측정한 결과, 3~6월 에는 전체 포란수에서 성숙난(난경 2 mm 이상)이 차지 하는 비율이 12.7~17.5%이었다. 7월에도 2 mm 이상의 성숙난들이 존재하지만, 8~9월에 이르면 성숙난은 모 두 방출, 흡수 및 퇴화되어 난경 1 mm 이하의 미성숙난 이 대부분이다. 10월에는 난이 성장하기 시작하여 1 mm 내외까지 성장하고, 일부는 대략 2 mm 이내로 성장하였 다(Fig. 9).
Fig. 5. Gonadosomatic indices for individual females and males of Korean bitterling, Acheilognathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from April in 2002 (○‹1 year old,
● = 2 years old, ★›3 years old, confidence kernel
= 0.95).
20 30 40 50 60 70 80
Body length (mm) Male (n = 69)
Female (n = 91) 5
4
3
2
1
0
10
8
6
4
2
0
Gonadosomatic index (%)Gonadosomatic index (%)
20 30 40 50 60 70 80
Body length (mm)
6
.생식소 발달에 따른 생식주기
월별로 조사된 난소와 정소의 생식세포의 발달을 조 직학적으로 관찰한 결과 생식주기는 9~10월에는 성장 기, 이듬해 3~4월에는 성숙기, 5~6월은 완숙 및 산란 기, 7~8월은 퇴화 및 휴지기로 구분할 수 있었다.
1)
난소의 발달9월과 10월에 채집된 개체의 난소에서는 난경 150~ 225µm의 주변인기(perinucleolus stage)난모세포와 난 경 950~1,025µm의 난황구기(yolk stage)난모세포들이 난소소엽의 대부분을 차지하여 성장기로 구분 할 수 있 었다. 9월의 수온은 평균 13.7�C에서 10월에 5.4�C까지 하강하였지만, 암컷개체들의 GSI는 4.96±1.35까지 상
승하였다. 3월에는 난소소엽 내에 난경이 1,625~1,700 µm의 성숙란이 다수 발견되며, 4월에 이들 성숙란은 더 욱 성장하여 난경이 1,850~2,025µm까지 성장하고, 소 수의 염색인기(chromatin nucleolus stage)난모세포와 주변인기(perinucleolus stage)난모세포 및 성장 중인 난 황구기 난모세포, 난황포기(yolk vesicle stage)난모세포 Fig. 6. Least squares means of gonadosomatic index of each age in Acheilognathus signifer sampled at Naechoncheon in
Hongcheon-gun, Kangwon-do from April in 2002.
Table 1. Monthly variations in sex ratio of Korean bitter- ling, Acheilognathus signifer
Months No. of No. of Total Sex ratio x2 females males numbers (F/(F±M)) (chi-squared)
Mar. 11 6 17 0.65 1.74
Apr. 91 69 160 0.57 3.03
May 68 64 132 0.52 0.12
Jun. 56 44 100 0.56 1.44
Jul. 45 33 78 0.57 1.85
Aug. 128 64 192 0.67 21.33
Sep. 84 96 180 0.47 0.80
Oct. 30 26 56 0.54 0.29
Total 513 402 915 0.56 13.47
The critical value for x2goodness-of-fit test of equal numbers of females and males (1 df) at 95% significance is 3.84.
Fig. 7. Linear regression of between body length and fec- undity by log transformation in the female Korean bitterling, Acheilognathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from April 2002.
1 2 3
Age 3
2
1
0
7
6
5 4
3
2
1
0
Gonadosomatic index (%) Gonadosomatic index (%)
1 2 3
Age
1.65 1.70 1.75 1.80
Body length 2.5
2.4
2.3
2.2
2.1
Fecundity
Male (n = 69) Female (n = 91)
가 관찰되어 다회산란종에서 흔히 나타나는 특징인 모든 단계의 난모세포가 관찰되었다. 이시기의 암컷의 GSI는 4.96±1.35으로 연중 최대치로 나타났다. 5~6월은 본 종의 산란성기로 난소 내에는 난경이 2,000µm 이상의 완숙란과 성숙란이 함께 관찰되며, 잔존여포의 출현도 확인할 수 있다. 이 들 성숙란은 완숙란의 산란 후 일정 기간동안 성장 후 완숙란으로 방란되어 다회산란을 하 게 된다. 이 시기는 완숙 및 산란기로 구분할 수 있으며, 5월에 GSI가 6.22±1.71에서 6월에 4.30±1.18로 낮아 져, 이 시기에 산란이 집중적으로 이루어지는 것을 알 수 있다. 7~8월에는 산란기 직후의 시기로, 난소는 수 축되어있고, 퇴화 중인 난모세포가 관찰되어 퇴화 및 휴 지기로 구분할 수 있는데, 8월의 GSI는 2.02±0.91로 연중 최저치로 나타났다(Fig. 10).
2)
정소의 발달9월과 이듬해 10월에 채집된 개체의 정소에서는 정원
세포(spermatogonium)와 정모세포(spermatocyte)가 출 현하여 성장기로 구분 할 수 있는데, 10월에GSI는0.97
±0.42로 상승하기 시작하였다. 3~4월에 채집된 개체의 GSI는 상승하여 4월에 2.84±0.55에 이르며, 산란 전 성숙기로서 정소 내에는 다수의 정세포의 cyst와 일부 변태 된 정자가 출현하였다. 5~6월에 정소는 완숙 및 방정기를 나타내었는데, 정소의 조직상을 살펴보면 정소 소엽은 정자로 가득 차고, 방정이 일어난 cyst의 일부가 비어 나타나는데, 5월에 GSI가 2.72±0.99에서 6월에 1.12±0.58로 낮아진다. 7~8월은 소엽간질조직이 결합 조직으로 바뀌면서 확장되어 정세관이 수축되고, 정소소 엽 내에 정자들이 방정된 후, 소엽 내에 잔존하는 정자 들과 정세포들이 퇴화∙흡수되며, 8월의 GSI는 0.71± 0.38로 낮았다(Fig. 11).
고 찰
담수 경골어류의 생식소의 성숙, 산란 개시와 종료는 생식 관련 내분비 조절에 의해 결정되지만, 이러한 내분 비의 활성에 외부조건이 작용하며, 수온과 광주기는 주 된 환경적 외부조건이다(de Vlaming, 1972; Billard and Breton, 1978; Crim, 1982). 수온과 광주기는 산란이 일 어나는 계절에 따라 작용이 틀려지게 되는데, Shimizu and Hanyu (1993)의 연구 결과에서 춘계 산란종은 수온 상승이 산란 개시의 주된 요인으로 작용하고, 광주기는 촉진적 보상작용을 한다고 보고하였다. 이러한 결과를 토대로 본 종을 살펴보면 3월부터 높은 GSI값을 나타 내지만 3월에는 10�C 이하의 낮은 수온과 12시간 정도 의 장일 조건에서 산란은 이루어지지 않고, 온도가 20
�C, 장일 조건이 14 hr 이상으로 상승하는 5월에 주로 산란하는 것으로 추정된다. 또한 고수온은 생식소 활성 을 억제한다고 하였는데, 본 종도 마찬가지로7월과8월 의 고수온 시기에는 생식소 활성이 저해되는 것을 알 수 있었다.
A. signifer의 산란성기는 5월로 추정되고, 4월 하순부 터 6월 상순까지 산란하는 것으로 나타나 內 (1939) 의 결과와 거의 일치하였으나, 그 해의 기온, 강수량 등 에 따른 수온 변화에 따라 산란시기는 다소간의 차이가 있을 것으로 생각된다.
어류의 연간 생식 리듬의 유형을 수온과 광주기 등의 환경요인과 관련하여 봄 산란형(spring spawner), 봄-여 름 산란형(summer spawner), 여름 산란형(summer spawner), 봄-가을 산란형(spring-autumn spawner), 가 을산란형(autumn spawner), 겨울산란형(winter spawn- Table 2. The number of eggs from the ovary of Korean
bitterling, Acheilognathus signifer
No Body length Weight Gonad weight GSI Number
(mm) (g) (g) (%) of eggs
1 46.3 2.16 0.14 6.48 141
2 48.1 3.10 0.24 7.74 197
3 50.8 3.41 0.28 8.21 191
4 51.9 3.55 0.27 7.61 193
5 54.5 5.55 0.42 7.57 246
6 56.1 4.88 0.41 8.40 212
7 58.4 5.20 0.39 7.50 221
8 59.6 6.77 0.45 6.65 255
9 60.2 6.10 0.38 6.23 314
10 60.5 6.04 0.41 6.79 274
Average 225
Fig. 8. Monthly variations of fecundity in the female Kor- ean bitterling, Acheilognathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from March to October, 2002.
M A M J J A S O
Month 300
200
100
0
Number of eggs
n = 80
Fig. 9. Monthly variations of frequency distribution of egg diameter (the long axis) in the female Korean bitterling, Acheilog- nathus signifer sampled at Naechoncheon in Hongcheon-gun, Kangwon-do from March to October, 2002.
8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
10
8
6
4
2
0
8
6
4
2
0
0 1 2 3
Egg diameter (mm)
Mar. Jul.
Apr. Aug.
May Sep.
Jun. Oct.
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
0 1 2 3
Egg diameter (mm)
Frequency (%)Frequency (%)Frequency (%)Frequency (%) Frequency (%)Frequency (%)Frequency (%)Frequency (%)
Fig. 10. Photomicrographs of ovarian developmental phases of the Korean bitterling, Acheilognathus signifer. A-B: sec- tion of ovarian lobules in growing stage (September to October). C-D: section of ovarian lobules in mature stage (March to April). E-F: section of ovarian lobules in ripe and spent stage (May to June). G-H: section of ovarian lobules in degenerative and resting stage (July to August). Abbreviations: DO, degenerating oocyte; N, nucleus;
MO, mature oocyte; OCN, oocyte in the chromatin nucleolus stage; OPN, oocyte in the preinucleolus stage; OY, oocyte in the yolk stage; RO, ripe oocyte; YV, yolk vesicle. Scale bars = 25µm.
A B
D
F
H C
E
G
Fig. 11. Photomicrographs of testicular developmental phases of the Korean bitterling, Acheilognathus signifer. A: section of testicular lobules in growing stage (September to October). B-D: section of testicular lobules in mature stage (March to April). E-F: section of testicular lobules in ripe and spent stage (May to June). G-H: section of testicu- lar lobules in degenerative and resting stage (July to August). Abbreviations: ISC, interstitial cell; SC, spermato- cyte; ST, spermatid; SZ, spermatozoon. USZ, undischarged spermatozoon. Scale bars = 125µm.
A B
D
F
H C
E
G
er)으로 나눌 수 있는데(Aida, 1991), 본 종은 5월이 산 란성기인 것으로 추정 할 수 있어, 봄 산란형에 해당한 다. 한편 본 아과에 속하는 어종 중A. yamatsutae (송과 권, 1989, 1994), R. uyekii (안, 1995) 일본산 A. tabira (Shimizu and Hanyu, 1981), R. amarus (Verhoeven and Oordt, 1955)등이 봄산란형, 일본산 R. occellatus (Nishi and Takano, 1979)는 봄-여름 산란형, 일본산 A. rhom- beus (Shimizu et al., 1985)와 Pseudoperilampus typus (Shimizu and Hanyu, 1983)는 가을산란형 등으로 구분 할 수 있었다.
어류 난모세포의 발달양식에 따라 동시발달형(total synchronism), 난군동시발달형(group synchronism), 비 동시발달형(asynchronism)의 세가지로 분류하여 볼 때 (Marza, 1938), A. signifer는 산란기 암컷의 난소 내에 성장기의 난모세포로부터 성숙된 난에 이르기까지 여러 단계가 존재하여 다회산란종에서 흔히 나타나는 비동기 발달형에 속하며, 완숙한 난이 산란되면 성장중인 난이 완숙되어 또 다시 산란되는 것으로 보인다. 이와 더불어 산란관이 신장된 암컷의 복부를 압박 할 때 배출되는 성숙난이 30~50개정도 정도로 나타나, 산란을 시작하 면 1회에 약 30~50개의 알을 낳는 것으로 추산되며, 납자루, A. lanceolatus가 3회 이내의 산란횟수를 가진다 고 보고된 것으로 미루어 볼 때 산란횟수는 이와 유사 할 것으로 사료된다(Suzuki and Jeon, 1990a).
Suzuki and Jeon은 웅천천 수계의 A. lanceolatus를 인공 수정시키면서, 각각 85개와 106개의 성숙난을 얻 어, Acheilognathus속 어류 중에서 한번의 산란수가 많 은 편이라 하였고(suzuki and Jeon, 1990b), 큰납지리, Acanthorhodeus macropterus는 한 개체가 최소 5회 이 상 산란하며, 1회에 평균적으로 195개의 난을 산란하고 (Suzuki and Jeon, 1989), 가시납지리, Acan. gracilis의 경우에도 산란기에 최소 5회 이상 산란하며, 1회에 200
~300개의 난을 산란하는 것으로 추정하였는데(Suzuki and Jeon, 1990), A. signifer의 1회 산란수는 이 보다 적 은 점에서 주목된다.
한편11~2월까지는 낮은 수온과 결빙으로 인해 채집 하지 못하였으나, 안(1995)의 결과로 미루어 볼 때 A.
signifer의 생식주기 역시 이와 유사하여 12월에서 2월 에는 정체기 및 재 성장기를 거칠 것으로 추정되나 추 가 연구가 필요할 것으로 사료된다.
적 요
묵납자루, Acheilognathus signifer의 성적성숙과 산란
시기를 연구하기 위하여 2002년 3월부터 동년 10월까 지 북한강 수계의 홍천강 상류인 내촌천에서 조사, 연구 하였다.
생식소성숙도지수(GSI)는 4월에 암컷(GSI = 6.95± 0.83)과 수컷(GSI = 2.84±0.55) 모두 최대치로 나타났 고, 암컷은 8월(GSI = 2.02±0.91), 수컷은 9월에(GSI = 0.54±0.50)최저치를 보였으며, 10월부터 GSI가 상승하 였다. 산란관길이 지수(OI)는 5월에 최대로 신장(26.53
±8.34%)되었으며, 8월에 가장 축소(5.77±1.37%)되었 다. 암컷은 체장 45 mm 이상, 수컷은 체장40 mm 이상부 터 생식 활동이 가능 할 것으로 추정되었다. 암컷과 수 컷에서 산란에 참여 비율이 높은 연령군은 2년생이었으 며, 수컷은 암컷보다 3년생의 참여 비율이 높았다. 생식 주기는 3~4월의 성숙기 5~6월의 완숙 및 산란기, 7~ 8월의 퇴화 및 휴지기, 9~10월의 성장기로 구분되었다. 포란수는 141~314 (평균 225)개였으며, 성비에서 암컷 이 수컷보다 우세하게 나타나 암수의 성비가 1 : 1이 아 니었고, 암수의 성비는 유의한 차이를 보였다(x2 = 13.5, P⁄0.05). 본 종의 산란 성기는GSI가 높은 값을 나타내 고, 산란관이 최대로 신장되며, 난소와 정소가 최대로 성 숙하는5월로 추정되었다.
인 용 문 헌
Aida, K. 1991. Environmental regulation of reproductive rhythms in teleostei. Bull. Inst. Zool., Academia Sinsca, Monograph., 16 : 173~187.
Billard, R. and B. Breton. 1978. Rhythms of reproduction in teleost fish. In: Rhythmic Activity of Fishes. J. E.
Thorpe, ed. Academic Press Inc., London. pp. 31~53.
Crim, L.W. 1982. Environmental modulation of annual and daily rhythms associated with reproduction in teleost fish. Can. J. Fish. Aquat. Sci., 39 : 17~21.
de Vlaming, V.L. 1972. The effects of temperature and photoperiod on reproductive cycling in the estuarine gobiid fish Gillichthys mirabilis. NOAAfish. bull., 70:
1137~1152.
Holcik, J. 1962. Redescription of Acanthorhodeus asmussi (Dybowski) 1872 and description of Acanthorhodeus asmussi amurensis ssp. n. from the Amur river USSR.
Japan. J. Ichthyol., 9 : 153~162.
Holcik, J. and J.J. Duyvene DeWit. 1962. The taxonomic charateristics of hybrid Rhodeus. Copeian, 4 : 777~788.
Lagler, K.F., J.E. Bardach. and R.R. Miller. 1962. Ichthyol- ogy: the study of fishes. Jonh Wiley and Sons, Inc., New York London, pp. 289~290.
Marza, V.D. 1938. Histophysiologie de l’ovogenese. Paris, Hermanu & Cie Editeurs. 81pp.
Nishi, K. and K. Takano. 1979. Effects of photoperiod and temperature on the ovary of the bitterling, Rhodeus ocellatus ocellatus. Bull. Fac. Fish. Hokkaido Univ., 30(1) : 63~73.
Schimidt, R. E., 1982. Biology of the European Bitterling Rhodeus sericeus (Pisces : Cyprinidae) in the Bronix river. New York, USA : An apparently benign exotic species. Biology conservation, 24 : 157~162.
Shimizu, A. and I. Hanyu. 1981. Annual reproductive cycle of a spring-spawning bitterling, Acheilognathus tabira.
Bull. Japan. Soc. Sci. Fish., 48 : 1563~1568.
Shimizu, A. and I. Hanyu. 1983. Environmental regulation of spawning-period in autumn-spawning bitterling, Pseudoperilampus typus. Bull. Jap. Soc. Sci. Fish., 49 : 895~900.
Shimizu, A., K. Aida and I. Hanyu. 1985. Endocrine pro- files during the short reproductive cycle of an autumn- spawning bitterling, Acheilognathus rhombeus. Gen.
Comp. Endocrinol., 60 : 361~371.
Shimizu, A. and I. Hanyu. 1993. Factors involved in the development of and decline in photoperiodism as it relates to the gonadal activity of spring-spawning bit- terling, Acheilognathus tabira. J. Exp. Zool., 265 : 134
~143.
Suzuki, N. and S.R. Jeon. 1989. Development of the bitter- ling, Acanthorhodeus asmussi (Cyprinidae) with note on minute tubercles on the skin surface. Kor. J. Ichthy- ol., 1 : 73~82.
Suzuki, N. and S.R. Jeon. 1990a. Development of the bit- terling, Acheilognathus lanceolatus from Ungchon river, Korea. Kor. J. Ichthyol., 2 : 77~87.
Suzuki, N. and S.R. Jeon. 1990b. Development of the bit-
terling, Acanthorhodeus (= Acheilognathus) gracilis (Cyprinidae), with a note on minute tubercles on the skin surface. Kor. J. Ichthyol., 2 : 169~181.
Verhoeven, B and G.J. Van Oordt. 1955. The influence of light and temperature on the sexual cycle of the bitter- ling, Rhodeus amarus. Zool., pp. 628~632.
Wiepkema, P.R. 1961. An ethological analysis of the repro- ductive behavior of the bitterling (Rhodeus amarus Bloch). Arch. Neel. Zool., 14 : 103~199.
권오길. 1981. 의암호의 패류에 관한 연구(1) : Anodonta fukudai유생의 어체부착에 관하여. 육수지, 14 : 21~25.
김익수∙박종영, 2002. 한국의 민물고기. 교학사, 서울, pp. 61
~93.
박두신 최신석. 1998. 송사리와 대륙송사리의 산란, 난발생 및 자어의 성장. 육수지, 31 : 109~112.
백현민 송호복 권오길. 2002. 홍천강 상류에 서식하는 묵납자 루, Acheilognathus signifer의 연령과 성장. 한어지, 14 : 254~261.
송호복. 1994. 줄납자루, Acheilognathus yamatsutae Mori (잉어과)의 생태학적 연구. 강원대학교 박사학위논문. 182pp.
송호복∙권오길. 1989. 의암호에 서식하는 줄납자루(Acheilog- nathus yamatsutae Mori)의 산란 및 발생 특성에 관한 연구. 육수지, 22 : 51~70.
송호복∙권오길. 1994. 줄납자루, Acheilognathus yamatsutae (Cyprinidae)의 생식생태, 성적 성숙과 산란시기. 강원대 학교 과학기술논문집, 33 : 76~84.
송호복 최신석. 2000. 버들붕어, Macropodus chinensis의 생 식생태와 초기생활사. 육수지, 33 : 282~286.
안철민. 1995. 각시붕어, Rhodeus uyekii의 생식주기. 한어지, 7 : 33~42.
內 . 1939. , , , 內 .
, pp. 133~140.
Received : August 29, 2003 Accepted : November 15, 2003