https://doi.org/10.20307/nps.2016.22.4.231
231
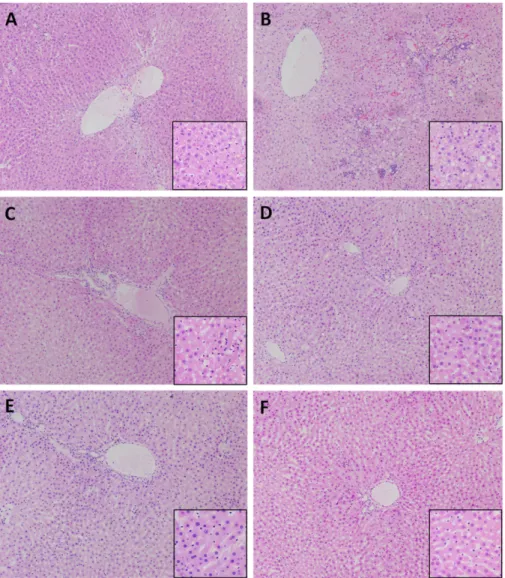
A Fruit Extract of Paeonia anomala Attenuates Chronic Alcohol-induced Liver Damage in Rats
Sarangerel Oidovsambuu
1, Ji Ho Yun
1,4, Kyungsu Kang
3, Batsuren Dulamjav
5, Jigjidsuren Tunsag
5, Eui Jeong Nam
1,6, and Chu Won Nho
1,2*
1
Natural Products Research Center, Korea Institute of Science and Technology (KIST) Gangneung Institute, Gangneung, Korea
2
Convergence Research Center for Smart Farm Solution, Korea Institute of Science and Technology (KIST) Gangneung Institute, Gangneung, Korea
3
Systems Biotechnology Research Center, Korea Institute of Science and Technology (KIST) Gangneung Institute, Gangneung, Korea
4
Department of Life Science, Sogang University, Seoul, Korea
5
Institute of Chemistry and Chemical Technology, Ulaanbaatar, Mongolia
6