생체 내 골 발생과정에서 고아핵수용체 Small Heterodimer Partner의 역할: 미세단층촬영기를 이용한 SHP 유전자 결손 생쥐의 골 분석
전남대학교 치의학전문대학원 치의학연구소 및 BK21 사업단, 광주보건대학 물리치료학과1, 남부대학교 방사선학과2
배인호․손혜주․김종상․정병철․백윤웅1․이준행2․고정태
Roles of Orphan Nuclear Receptor Small Heterodimer Partner in Bone Development: Microcomputed Tomographic Analysis of Bone Microarchitecture in
SHP Knockout Mice
In-Ho Bae, Hye-Ju Son, Jong-Sang Kim, Byung-Chul Jeong, Yun-Woong Paek1, Jun-Haeng Lee2, Jeong-Tae Koh
Dental Science Research Institute and BK 21, School of Dentistry, Chonnam National University,
Department of Physical Therapy2, Kwangju Health College, Department of Radiology3, Nambu University, Gwangju, Korea
Objectives: Orphan nuclear receptor small heterodimer partner (SHP) is involved in osteoblastic differentiation. This study was undertaken to demonstrate a role of SHP in in vivo bone development using microcomputed tomographic (microCT) analysis of SHP knockout (KO) mice.
Material & Methods: Tibia bones were harvested from 1-, 4-, 8- and 20-week-old wild type (WT) and SHP KO mice. The microarchitecture of tibial bone was analyzed using a microCT (Skyscan 1172; Skyscan, Kontich, Belgium). Samples were scanned at a resolution of 17 μm (isotropic). The X-ray was operated with 50 kV, 200 μA of energy, 1.2 sec of exposure time, and a 0.5 mm thick aluminum filter. Projections were acquired over an angular range of 180°. For quantification of the bone mineral density (BMD), the microCT was calibrated using 2 standard phantoms with densities of 0.25 and 0.75 g/cm3. The image slices were reconstructed and analyzed using CT analyzer software (CTan, Skyscan).
Results: The CT values of tibial trabecular bone were significantly decreased in SHP KO compared to WT at 20-week-old mice determined by microCT; (bone volume / tissue volume [BV/TV, 40%], BMD [80%], and trabecular number [Tb.N, 50%]). However, the CT values were not significantly different between WT and SHP KO in cortical bone. Furthermore, the qualitative indices of trabecular bone such as the structure model index (SMI) and the polar moment inertia (PMI) did not differ between WT and SHP KO mice.
Conclusion: These microCT results supports that SHP may act as a positive regulator of trabecular bone formation.
Key Words: Bone formation, Microarchitecture, Microcomputed tomography, Small heterodimer partner, Trabecular bone
Received: February 8, 2010 Revised: March 13, 2010 Accepted: April 6, 2010
Corresponding Author: Jeong-Tae Koh, Dental Science Research Institute and BK 21, School of Dentistry, Chonnam National University, 671, Jebongro, Dong-gu, Gwangju 501-757, Korea
Tel: +82-62-220-4433, Fax: +82-62-227-4008 E-mail: [email protected]
* 본 논문은 전남대학교병원 임상의학연구소 학술연구비(CRI09004-1) 및 2007년도 정부재원(교육인적자원부 학술연구조성사업비)으로 한국학술진흥재단의 지원을 받아 연구되었음(KRF-2007-313-E00469).
뼈 조직은 신체의 지지, 칼슘과 인의 저장, 조혈기 능 및 뇌, 심장 등의 중요기관을 보호하는 역할을 하 며, 일생 동안 지속적인 형성(modeling)과 개조(re- modeling)의 과정을 거치면서 항상성을 유지하고 있 다.1,2 뼈는 형태학적으로 크게 피질골(cortical bone) 과 지주골(trabecular bone)로 나누어진다. 피질골은 외피를 덮고 있는 조직으로서 매우 치밀하게 배열된 콜라겐 섬유로 구성되어 있으며, 기계적인 보호기능 을 수행한다. 지주골은 뼈 내부에 채워지는 조직으 로 엉성한 다공성 구조로 되어 있으며 대사 물질의 공급 역할을 담당한다. 이와 같은 뼈의 성장, 발달 그리고 유지는 매우 조절이 잘된 과정이며, 뼈를 형 성하는 골모세포(osteoblast)와 뼈를 흡수하는 파골세 포(osteoclast), 그리고 뼈의 기질 속에서 그 평형 유 지에 관여하는 골세포(osteocyte)에 의해 조절된다.3,4 파골세포의 활성이 비정상적으로 증가하면 뼈가 쉽 게 부러지는 골다공증이 초래되며, 골모세포의 활성 이 증가하게 되면 골밀도(bone mineral density; BMD) 의 증가와 함께 골석화증이 일어나게 된다.5 골다공증 치료를 위한 기존의 연구는 주로 파골세 포 분화를 억제하는 신호전달 경로를 밝혀서, 해당 경로를 차단하는 약물을 찾는 데 초점을 맞추어 왔 다.6-8 최근에는 골수(bone marrow)에 많이 존재하는 줄기세포로부터 골모세포의 분화를 촉진하는 인자 를 찾아 골다공증의 치료에 적용하려는 시도가 이루 어지고 있다.9-10
골모세포 분화를 촉진하는 전사인자로는 Runx2, Osterix, Dlx 등이 알려져 있으며, VitD3 수용체 (VDR), 에스트로겐 수용체(ER), 레티노이드 X 수용 체(RXR) 등과 같은 핵 수용체의 관여도 알려지고
있다.11-15 최근에는 리간드가 밝혀지지 않은 고아 핵
수용체(orphan nuclear receptor)도 골모세포의 분화에 중요한 역할을 하는 것으로 알려지고 있다. 그 예로 에스트로겐 수용체와 구조적으로 관련이 있는 에스 트로겐 α 관련 수용체(ERRα)는 골모세포 분화과 정 중에 현저한 발현 증가를 보이며, 골조직 기질 중 의 하나인 osteopontin의 발현을 조절한다.16 ERRr는 BMP-2에 의한 골모세포의 분화과정 중에 발현이 증 가하며, Runx2의 전사활성을 억제하여 골 생성을 저 해한다.17 최근 본 연구진은 또 다른 고아 핵 수용체
인 small heterodimer partner (SHP)가 HDAC4와 경쟁 적으로 작용하여 골모세포 분화의 중요한 전사인자 인 Runx2 전사활성을 증가시킴으로써 골형성 촉진 에 관여한다는 내용을 보고하였다.18
생체 내 골구조에 대한 전통적인 조직학적 평가는 동물을 희생시킨 후 탈회, 조직처리과정, 염색과정 등을 거치게 되는데, 이러한 과정 중에 장기간의 탈 회에 의한 시편의 손상이 나타날 수 있으며, 조직학 적 평가에 사용된 시편에서의 물성 평가가 어렵다는 단점이 있다. 그러나 1980년대에 미세단층촬영기 (microcomputed tomography, microCT)가 개발되면서 골의 2차원적 이미지에 대한 분석 및 3차원적 골 미 세구조(bone microarchitecture)에 대한 비파괴적 분석 이 가능하게 되었다.19 MicroCT의 사용으로 인해 지 주골에 대한 체적, 두께, 간격 등에 대한 정량적 분 석(quantitative analysis)과 더불어 structure model index (SMI), polar moment inertia (PMI) 등을 통한 정 성적 분석(qualitative analysis)이 가능하게 됨으로써 생체 내 유전 인자에 의한 골 조직의 변화를 평가하 는 데 microCT의 활용 빈도가 점차 증가하고 있다.
본 연구는 생체 내 골형성 과정에 고아 핵 수용체 SHP가 어떠한 영향을 나타내는지를 확인하기 위해 microCT를 이용하여 SHP 유전자가 결여된 생쥐 (SHP knockout mice; SHP KO)에서 주령에 따른 경골 미세구조의 변화를 정상쥐(wild type; WT)와 비교하 고자 하였다.
대상 및 방법
1. 실험 동물
본 연구에 사용된 congenic C57BL/6 SHP KO 생쥐 는 C57BL/6와 129/Sv 교배 후 10세대 역교배를 통하 여 만들어졌으며, 분당 서울대병원에서 분양받아 사 용하였다.20 주령에 따른 골 미세구조의 변화를 관찰 하기 위하여 각각 4마리의 생후 1, 4, 8, 20주 WT와 SHP KO 생쥐를 이산화탄소 가스로 희생시켰다. 경 골(tibia)만을 추출하여 10% 포르말린에 하루 동안 고정시킨 후, microCT를 이용하여 경골의 미세구조 를 분석하였다. 본 연구는 전남대학교 동물 보호 및 사용에 관한 가이드라인과 법규에 따라 수행되었다.
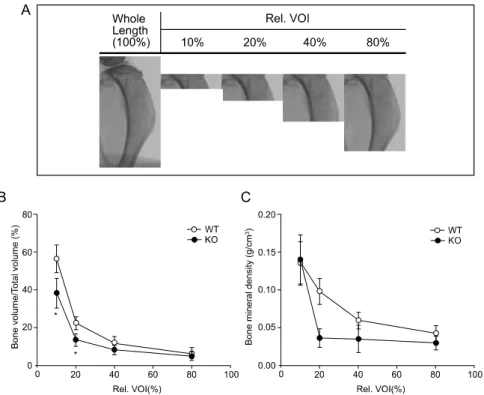
Fig. 1. Illustration of the VOI at proximal tibial metaphysis (A), and BV/TV (B) and BMD (C) values analyzed according to designated volume of interest. Total volume was set from epiphyseal plate to upper limit of lower junction between fibular and tibial bones. Data are expressed as mean ±SD. n=4, *P<0.05
2. 미세단층촬영 및 단면 재구성
시편의 단층 영상은 microCT (Skyscan1172; Skyscan, Kontich, Belgium)를 이용하여 다음과 같은 조건하에 채득하였다. 원추형 빔(cone beam) 방식의 X-ray source 를 이용하여 50 kV, 200 μA 하에서 시편에 방사선을 조사하였고, 이때 해상도(spatial resolution)는 17 μm 로 조절하였다. 시편을 0.4도씩 회전시키면서 매회 1.2 초간 방사선을 조사하였다. 촬영은 포커스 스폿(focus spot)에서 방사된 빔이 시편을 투과하여 charge-coupled devices (CCD) 카메라에 영상이 맺히는 원리를 이용하 였다. 채득된 영상은 NRecon 소프트웨어(Skyscan, Belgium)를 이용하여 150개의 2차원 횡단면으로 재구 성되었고, 각 단면간의 거리는 80 μm로 설정하였다.21
3. 구조 분석
경골의 미세구조는 2차원적으로 채득된 영상의 골 영역만을 적용하여 CTan 소프트웨어(Skyscan, Belgium) 로 분석하였다. 지주골의 분석은 성장판으로부터
400μm를 제외한 나머지 부분을 0.2 mm 간격으로 나누어 총 4 mm를 측정 대상으로 하였다.22,23 재구성 된 2차원 이미지로부터 피질골과 지주골을 구분하여 관심영역(region of interest, ROI)을 설정한 후, 각 부 분의 bone volume/tissue volume (BV/TV), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp), trabecular number (Tb.N) 등의 양적 분석과 polar moment inertia (PMI), structure model index (SMI) 등 의 구조적 파라미터(structural parameters) 값을 측정
하였다.24,25 BMD는 0.25와 0.75 g/cm3의 hydroxya-
patite standard phantom에 대한 Hounsfield unit (HU) 값을 얻은 후, 측정하고자 하는 시편의 HU 값을 구 하여 계산식에 대응시켜 구하였다.26,27
4. 통계분석
WT와 SHP KO 생쥐 간의 주령에 따른 골 미세구 조의 통계학적 차이는 Mann-Whitney와 Kruskal- Wallis test (SPSS software Version 17.0, SAS Institute, Cary, NC)로 비교하였다. P<0.05를 통계적으로 유의
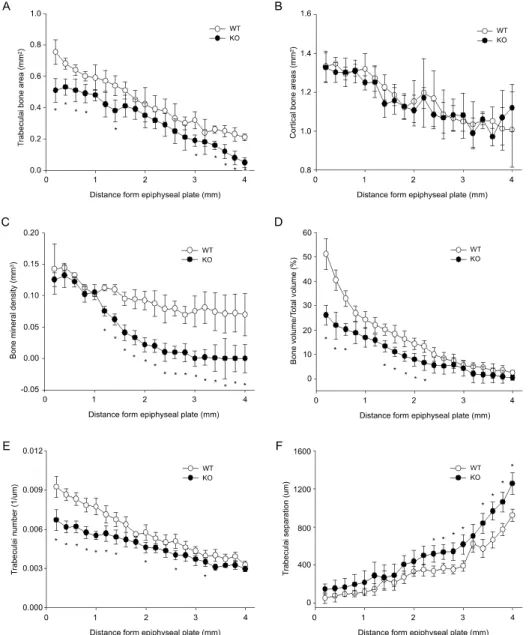
Fig. 2. Curve-fit analysis of trabecular a), cortical bone area b) and various trabecular bone microarchitecture (c~f) were represented. The analysis in WT and SHP KO mice showing changes in tibial metaphyseal trabecular bone parameters at every 0.2 mm. Four millimeters of distance from epiphyseal plate were set as analysis area. Data are expressed as mean ±SD. n=4, *P<0.05.
하다고 간주하였다.
결 과
1. 분석영역의 설정
대사과정이나 유전적 변이에 따른 지주골 차이를 분석하고자 할 경우, 일반적으로 proximal metaphy- ses 부분이 분석 영역으로 적용된다.28,29 해부학적으
로 지주골의 경우 성장판으로부터 멀어질수록 지주 골의 분포가 줄어드는 것으로 알려져 있으므로,30 분 석하고자 하는 영역(volume of interest; VOI)에 따라 분석 결과가 달라질 수 있다. 따라서 본 연구에서는 적절한 VOI 설정을 위하여 위치에 따른 지주골의 분포 변화를 먼저 확인하였다(n=4). 경골의 성장판 을 시작점으로 하여 비골과 경골의 하측 연결 부위 까지를 전체 부피(total volume)로 설정한 후, 전체 부
A B
WT
KO
WT
KO
1 4 8 20 1 4 8 20
After birth (weeks) After birth (weeks)
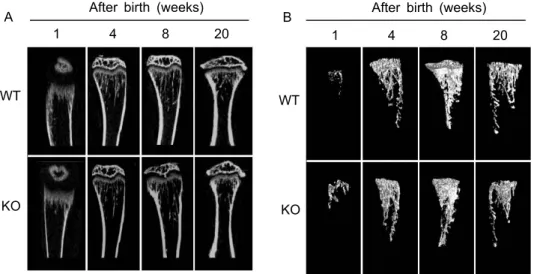
Fig. 3. Comparison of morphologies of vertical-sectional (A) and 3D trabecular images (B) between WT and SHP KO mice at 1, 4, 8, 20 weeks aged. The vertical section site was right angle center of Y and Z axis.
피의 20, 40, 60, 80%에 해당하는 부위의 미세 골 변 화를 관찰하였다(Fig. 1A). 생후 8주 생쥐의 경우 경 골 전체의 길이는 20 mm 정도였으며, BV/TV과 BMD 의 경우, 측정영역을 전체 길이의 20% (약 4 mm)로 설정하였을 때 WT와 SHP KO 간의 의미 있는 차이 가 관찰되었다(Fig. 1B, 1C). 40~80%의 설정 범위에 서는 지주골의 분포가 감소된 영역 설정이 증가하여 WT와 SHP KO 간의 골 평가 지표의 차이가 줄어드 는 경향을 보였다. 따라서 본 연구에서는 시편 전체 길이의 20% 이내에서 같은 크기의 측정 영역을 설정 하여 골 미세구조의 변화를 평가하였다.
2. 측정 부위에 따른 2차원적 골 분석
골조직 부위에 따라 SHP의 작용이 다르게 나타날 수 있으므로 경골 전체의 20% 이내에서 피질골과 지주골에 대한 영역을 각각 설정하여 성장판으로부 터의 거리에 따른 골량의 변화를 SHP KO 군과 WT 간에 비교하였다. 지주골의 경우 성장판으로부터 거 리가 멀어짐에 따라 골량이 감소하였으며, SHP KO 군의 골량이 WT에 비해 더 적었다(Fig. 2A). 피질골 의 경우도 성장판으로부터 거리가 멀어짐에 따라 골 량이 감소하였으나, SHP KO 군과 WT 간의 유의한 차이는 관찰되지 않았다(Fig. 2B). SHP가 지주골 형 성에 관여한다는 결과에 따라 전체 경골 길이의 20% 영역 내에서 0.2 mm 간격으로 단층 촬영한 후
각 지주골의 단편에 대한 다양한 CT 측정값을 비교 하였다. 성장판에서 멀어질수록 WT에 비해 SHP KO 군에서 현저한 BMD의 감소가 관찰되었으며 (Fig. 2C), BV/TV와 Tb.N 값도 SHP KO 군에서 WT 에 비해 낮은 값을 보이지만 성장판에서 거리가 멀 어질수록 그 차이는 감소하였다(Fig. 2D, 2E). Tb.Sp 값은 SHP KO군에서 WT에 비해 높은 값을 보였다 (Fig. 2F).
3. 주령에 따른 골 미세구조의 변화
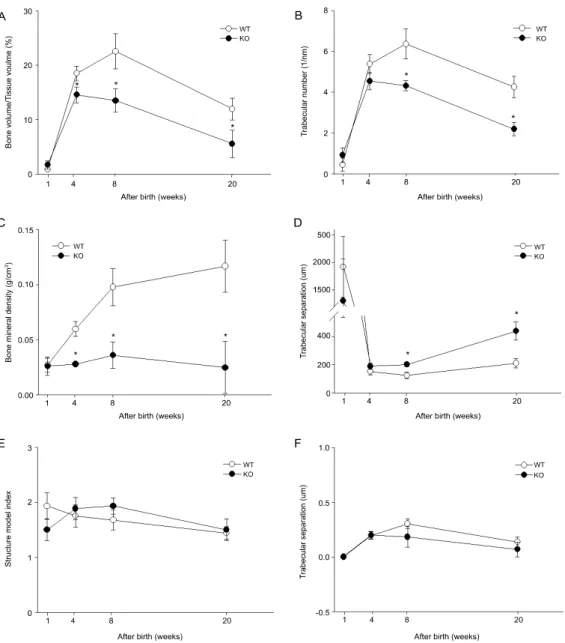
시기에 따른 골 발생과정에서의 SHP 역할을 알아 보기 위하여 4마리의 생후 1, 4, 8, 20주 SHP KO와 WT 생쥐의 경골을 위에서 설정한 관심영역 내에서 3차원으로 재구성한 후, 골 미세구조 변화를 비교 관찰하였다. Fig. 3은 각 시기에서의 경골의 2차원 종단면 이미지와 지주골을 3차원으로 재구성한 이 미지를 보여주고 있다. 정량적 수치를 확인하기 위 해 CTan을 이용하여 분석한 결과 생후 1주에는 WT 와 SHP KO 간의 뚜렷한 골 미세구조의 변화가 관찰 되지 않았으며, 4, 8주에서는 SHP KO 군에서 WT에 비해 지주골의 양이 유의하게 감소되어 있었다(Fig.
3, 4A). 생후 20주에는 8주에 비해 지주골 양이 감소 하였으며(약 40%), WT와 SHP KO 간의 차이 역시 8 주에 비해 감소하였다(Fig. 3, 4A). Tb.N과 BMD 역 시 생후 8주에 SHP KO에서 각각 약 33%, 65%의 감
Fig. 4. Differences of trabecular microarchitectures with aging in WT and SHP KO mice were represented as follows;
BV/TV (A), Tb.N (B), BMD (C), Tb.Sp (D), SMI (E), PMI (F). Analysis was performed at 20% of VOI of total tibial volume at 1, 4, 8, 20 weeks aged mice. Data are expressed as mean ±SD. n=4, *P<0.05.
소를 나타냈으며, 20주에는 각각 50%, 80%로 그 차 이가 증가하였다(Fig. 4B, 4C). 이와 같은 골 형성과 관련된 CT측정값의 차이는 전체적으로 8주까지는 지속적으로 증가하였지만, 20주에는 감소하는 경향 을 나타냈다. 골 형성 관련 CT 측정값의 증가와 더 불어 공간지표 수치인 Tb.Sp 값도 시간이 지남에 따 라 증가하였으며, WT와 SHP KO 간의 차이도 증가 하였다(Fig. 4D). 그러나 형태학적 구조와 관련된
SMI나 뒤틀림에 대한 골의 저항값을 나타내는 PMI 값은 WT와 SHP KO 간의 유의한 차이가 관찰되지 않았다(Fig. 4E, 4F).
고 찰
핵 수용체는 다양한 조직에서 발현되며, 다양한 전사인자의 상호작용에 의해 표적유전자들의 발현
을 조절하는 것으로 알려져 있다. 본 연구진은 최근 고아 핵 수용체인 SHP가 골모세포 분화에 과정에 관여한다는 결과를 보고하였으며, 또한 생체 내 골 발생 과정에서의 SHP의 역할에 관한 연구 필요성을 제시하였다.18
생체 내 골발생의 평가는 분석 방법 등의 다양한 요소에 의해 그 결과가 달라질 수 있다. 최근에는 기 존의 해부학적 방법에 비해 조작이 간편하고 실험자 의 요구에 따라 다양한 분석을 수행할 수 있는 장점 을 지닌 microCT 분석 방법이 소개되어 많은 연구에 적용되고 있다. 본 연구에서도 microCT를 이용하여 시기에 따른 SHP KO 생쥐의 경골 미세구조 형성과 정의 변화를 관찰하여, 생체 골 발생에서의 SHP의 역 할을 알아보고자 하였다. 대사과정이나 유전적 변이 에 따른 지주골 차이를 분석하고자 할 경우, 일반적 으로 proximal metaphyses 부분이 분석 영역으로 적용 된다. MicroCT를 이용해 골 미세구조를 분석하고자 할 때 어느 영역을 얼마만큼 VOI로 설정하였는지에 따라 측정결과와 해석이 달라질 수 있다. MicroCT를 이용한 골 구조를 분석한 대부분의 보고들은 우선 설 정된 VOI에 대한 정보를 제시한다.28,29 하지만, 이들 은 결과 도출에 사용된 VOI만을 제시하여 분석의 결 과가 VOI에 제한되어 있다. 이는 VOI을 달리하였을 때 결과는 달라질 수 있음을 시사한다. 따라서 본 연 구에서는 결과 도출을 위해 사용된 VOI를 설정하는 데 객관적인 근거를 제시하고자 VOI에 따른 결과의 상이함을 설명하고자 하였다. VOI를 성장판으로부터 경골 전체 부피의 20% (4 mm)로 설정하였을 때, WT 와 SHP KO 간의 골 미세구조의 평가가 적절함을 확 인하였다(Fig. 1). 또한 본 연구에서는 ROI를 지주골 과 피질골로 분리 설정하여 CT 측정값을 비교함으로 부위에 따른 SHP의 작용이 다르게 나타나는지를 알 아보았다. 지주골 분석에서 WT와 SHP KO 군 간에는 CT 측정값의 유의한 차이가 관찰되었지만, 피질골 분석에서는 유의한 차이가 없었다. 이는 SHP가 피질 골에 비해 상대적으로 세포의 성분이 많이 차지하는 지주골에서 더 큰 작용을 나타내고 있음을 시사한다 (Fig. 2A, 2B). 2차원 지주골 분석에서 WT와 SHP KO 군 간의 차이가 유의하더라도(Fig. 1), 좀 더 정확하고 객관적인 평가를 위해서는 3차원적 영역설정(VOI)
내에서 BV/TV, BMD, Tb.N, Tb.Sp 등의 경향을 확인 할 필요가 있다. SHP KO 생쥐의 3차원 영역에서의 분석 결과, 성장판에서 멀어질수록 BMD, BV/TV, Tb.N 등 골 관련 CT 값이 줄어드는 것을 확인할 수 있었다(Fig. 2C-2E). BMD의 경우 WT와 SHP KO 군 간의 차이가 성장판에서 멀어질수록 증가하였지만 (Fig. 2C), BV/TV와 Tb.N의 차이는 감소하였다(Fig.
2D, 2E). 이는 SHP의 작용이 연골화 골화과정을 겪고 있는 성장판에 가까운 부분보다는 성장판에서 조금 떨어져 있는 성숙된 지주골에서 더 크다는 것으로 해 석될 수 있다. 20주령 생쥐의 BV/ TV가 감소하는 것 은(Fig. 3B, Fig. 4A) 실험동물의 노화에 따른 파골세 포 활성화 등으로 인한 골 소실로 이해할 수 있을 것 이다. 하지만 20주령 WT군의 골 부피 감소에도 불구 하고 BMD는 감소하지 않았다(Fig. 4C). 이는 BMD가 파골세포보다는 골내 칼슘 등의 미네랄 비율에 영향 을 받기 때문인 것으로 판단된다.
MicroCT는 골 미세구조의 양적 분석(BV/TV, Th.N. Tb.Sp 등)은 물론 질적 평가(PMI, SMI 등)도 예측할 수 있는 정보를 제공한다. 본 연구 결과, WT 와 SHP KO 군 간의 PMI, SMI 등의 유의한 질적 차 이는 관찰되지 않았다(Fig. 4E, 4F). 이는 SHP 유전 자 결여만으로는 지주골의 형태나 외부 힘에 대한 저항 정도에는 큰 영향을 미치지 않는다는 것을 의 미한다. SMI는 지주골의 형태학적 특성을 나타내는 CT 값으로 그 값이 0에 가까울수록 판구조를 의미 하며, 3에 가까울수록 막대구조를 의미한다. 기존의 연구 결과, 골다공증이 유도된 조건에서는 SMI 값이 감소되는 것으로 보고된 바 있다.31,32 본 연구에서 20 주까지의 연령 변화에도 SHP KO 생쥐의 SMI 값의 변화가 없는 것으로 보아 골다공증 발생에 SHP가 크게 관여하지는 않는 것으로 판단된다. 좀 더 정확 한 평가를 위해서는 난소제거에 의한 골다공증 유발 동물 모형 등에서의 추가적인 연구가 필요하다. 또 한 결과를 도출하는데 사용된 샘플의 수가 적다는 제한점이 있으며 좀 더 많은 수의 샘플의 분석이 필 요할 것이다.
본 연구 결과를 요약해 보면, 생후 4, 8주령에서 SHP KO 군이 WT에 비해 골형성이 저해되어 있으 며, 20주의 생쥐에서는 그 차이가 감소되는 경향을
보인다. 이러한 결과는 SHP가 골 형성에 미치는 영 향은 생리적으로 노화된 시기(20주)보다는 골 형성 이 활발한 시기(4, 8주)에 더 크게 작용하며, 주로 지 주골의 형성에 관여하는 인자임을 시사한다.
감사의 글
본 연구에 사용된 SHP KO 생쥐를 제공해 주신 분 당 서울대병원 박영주 교수님께 감사드립니다.
참 고 문 헌
1. Olsen BR, Reginato AM, Wang W. Bone develop- ment. Annu Rev Cell Dev Biol 2000;16:191-220.
2. Suda T, Nakamura I, Jimi E, Takahashi N.
Regulation of osteoclast function. J Bone Miner Res 1997;12:869-79.
3. Hofbauer LC, Khosla S, Dunstan CR, Lacey DL, Boyle WJ, Riggs BL. The roles of osteoprotegerin and osteoprotegerin ligand in the paracrine regula- tion of bone resorption. J Bone Miner Res 2000;
15:2-12.
4. Yu X, Huang Y, Collin-Osdoby P, Osdoby P.
CCR1 chemokines promote the chemotactic recruit- ment, RANKL development, and motility of osteo- clasts and are induced by inflammatory cytokines in osteoblasts. J Bone Miner Res 2004;19:2065-77.
5. Lam DK, Sándor GK, Holmes HI, Carmichael RP, Clokie CM. Marble bone disease: a review of osteopetrosis and its oral health implications for dentists. J Can Dent Assoc 2007;73:839-43.
6. Anderson WF. Human gene therapy. Science 1992;
256:808-13.
7. Mulligan RC. The basic science of gene therapy.
Science 1993;260:926-32.
8. Tsuda E, Goto M, Mochizuki S, Yano K, Kobayashi F, Morinaga T, et al. Isolation of a novel cytokine from human fibroblasts that specifi- cally inhibits osteoclastogenesis. Biochem Biophys Res Commun 1997;234:137-42.
9. Sampath TK, Maliakal JC, Hauschka PV, Jones WK, Sasak H, Tucker RF, et al. Recombinant human osteogenic protein-1 (hOP-1) induces new bone formation in vivo with a specific activity comparable with natural bovine osteogenic protein and stimulates osteoblast proliferation and differen- tiation in vitro. J Biol Chem 1992;267:20352-62.
10. Wang EA, Rosen V, D'Alessandro JS, Bauduy M, Cordes P, Harada T, et al. Recombinant human bone morphogenetic protein induces bone forma- tion. Proc Natl Acad Sci USA 1990;87:2220-4.
11. Franceschi RT, Ge C, Xiao G, Roca H, Jiang D.
Transcriptional regulation of osteoblasts. Cells Tissues Organs 2009;189:144-52.
12. Kobayashi T, Kronenberg H. Minireview: transcrip- tional regulation in development of bone. Endocri- nology 2005;146:1012-7.
13. Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997;89:755-64.
14. St-Arnaud R. The direct role of vitamin D on bone homeostasis. Arch Biochem Biophys 2008;473:225- 30.
15. Marie PJ. Transcription factors controlling osteo- blastogenesis. Arch Biochem Biophys 2008;473:98- 105.
16. Zirngibl RA, Chan JS, Aubin JE. Estrogen receptor-related receptor alpha (ERRalpha) regulates osteopontin expression through a non-canonical ERRalpha response element in a cell context- dependent manner. J Mol Endocrinol 2008;40:61- 73.
17. Jeong BC, Lee YS, Park YY, Bae IH, Kim DK, Koo SH, et al. The orphan nuclear receptor estro- gen receptor-related receptor gamma negatively regulates BMP2-induced osteoblast differentiation and bone formation. J Biol Chem 2009;284:14211- 8.
18. Jeong BC, Lee YS, Bae IH, Lee CH, Shin HI, Ha HJ, et al. The Orphan Nuclear Receptor SHP is a Positive Regulator of Osteoblastic Bone Formation.
J Bone Miner Res 2009;25:262-74.
19. Pan S, Liou W, Shih A, Park MS, Wang G, Newberry SP, et al. Experimental System for X-ray Cone-Beam Microtomography. Microsc Microanal 1998;4:56-62.
20. Park YJ, Qatanani M, Chua SS, LaRey JL, Johnson SA, Watanabe M, et al. Loss of orphan receptor small heterodimer partner sensitizes mice to liver injury from obstructive cholestasis. Hepatology 2008;47:1578-86.
21. Gabet Y, Kohavi D, Kohler T, Baras M, Müller R, Bab I. Trabecular bone gradient in rat long bone metaphyses: mathematical modeling and application to morphometric measurements and correction of implant positioning. J Bone Miner Res 2008;23:48- 57.
22. Bouxsein ML, Myers KS, Shultz KL, Donahue LR, Rosen CJ, Beamer WG. Ovariectomy-induced bone loss varies among inbred strains of mice. J Bone Miner Res 2005;20:1085-92.
23. Ackert-Bicknell CL, Salisbury JL, Horowitz M, DeMambro VE, Horton LG, Shultz KL, et al. A chromosomal inversion within a quantitative trait locus has a major effect on adipogenesis and osteo- blastogenesis. Ann N Y Acad Sci 2007;1116:291- 305.
24. Odgaard A. Three-dimensional methods for quanti- fication of cancellous bone architecture. Bone 1997;
20:315-28.
25. Hildebrand T, Rüegsegger P. Quantification of Bone Microarchitecture with the Structure Model Index. Comput Methods Biomech Biomed Engin
1997;1:15-23.
26. Hui SL, Gao S, Zhou XH, Johnston CC, Jr., Lu Y, Glüer CC, et al. Universal standardization of bone density measurements: a method with optimal properties for calibration among several instru- ments. J Bone Miner Res 1997;12:1463-70.
27. Lu Y, Fuerst T, Hui S, Genant HK. Standardization of bone mineral density at femoral neck, trochanter and Ward's triangle. Osteoporos Int 2001;12:438- 44.
28. Gabet Y, Kohavi D, Müller R, Chorev M, Bab I.
Intermittently administered parathyroid hormone 1-34 reverses bone loss and structural impairment in orchiectomized adult rats. Osteoporos Int 2005;
16:1436-43.
29. Bonnet N, Benhamou CL, Brunet-Imbault B, Arlettaz A, Horcajada MN, Richard O, et al.
Severe bone alterations under beta2 agonist treat- ments: bone mass, microarchitecture and strength analyses in female rats. Bone 2005;37:622-33.
30. Weber MH, Sharp JC, Hassard TH, Orr FW.
Normal murine bone morphometry: a comparison of magnetic resonance microscopy with micro X- ray and histology. Skeletal Radiol 2002;31:282-91.
31. Dai R, Ma Y, Sheng Z, Jin Y, Zhang Y, Fang L, et al. Effects of genistein on vertebral trabecular bone microstructure, bone mineral density, micro- cracks, osteocyte density, and bone strength in ovariectomized rats. J Bone Miner Metab 2008;26:
342-9.
32. Zhang ZM, Li ZC, Jiang LS, Jiang SD, Dai LY.
Micro-CT and mechanical evaluation of subchond- ral trabecular bone structure between postmeno- pausal women with osteoarthritis and osteoporosis.
Osteoporos Int 2010;21:1383-90.