Correspondence to: Choong-Yeol Lee, Department of Physiology, College of Oriental Medicine, Kyungwon University, 65, Bogjeong- dong, Sujeong-gu, Seongnam 461-701, Korea
Tel: +82-31-750-5419, E-mail: [email protected] Received August 15, 2009, Revised August 31, 2009
Accepted September 5, 2009
Effect of N-acetylcysteine on Hydrogen Peroxide-Induced Apoptosis in Mouse C2C12 Myoblast Cells
*Department of Physiology, College of Oriental Medicine, Kyungwon University, Seongnam,
†
Department of Physiology, College of Medicine, Kyung Hee University, Seoul,
‡
Department of Exercise Physiology, School of Sport Science, Sungkyunkwan University, Suwon, Korea
Ji-Eun Koh*, Ye-Jin Kim
†, Mi-Hyeon Jang
†, Mal-Soon Shin
†, Chang-Ju Kim
†, Tae-Woon Kim
‡, Choong-Yeol Lee*
Reactive oxygen species (ROS) plays an important role as a mediator of skeletal muscle damage after strenuous exercise and during aging. In the present study, the protective effect of the antioxidant N-acetylcysteine (NAC) against hydrogen peroxide (H
2O
2)-induced cytotoxicity in myoblast C2C12 cells was investigated. Through morphological and biochemical analysis, it was demonstrated that C2C12 cells treated with H
2O
2exhibited several features of apoptosis. The apoptotic characteristic was decreased in cells treated with NAC prior to H
2O
2exposure. In addition, it was shown that H
2O
2increased the expression of bax but decreased the expression of bcl-2. Pre-treatment with NAC intervened H
2O
2-induced increase in the bax expression level and alleviated H
2O
2-induced suppression of bcl-2 expression. Our data show that NAC protects myoblast C2C12 cells against H
2O
2-induced apoptosis through the regulation of the expression of bax and bcl-2. (Korean J Str Res 2009;17:313∼321)
Key Words: N-acetylcysteine, Reactive oxygen species, Hydrogen peroxide, Skeletal muscle cells, Apoptosis
INTRODUCTION
Reactive oxygen species (ROS) have been implicated in the pathogenesis of various diseases. The key ROS responsible for such toxic effects are hydrogen peroxide (H
2O
2), superoxide anions (O
2-), and hydroxyl radicals (OH). H
2O
2is considered to be the main precursor of highly reactive free radicals that triggers apoptotic cell death in the brain, the lung, and the liver (Ratan
et al., 1994; Kanno et al., 1999; Liu et al., 2001). In addition, ROS is known to play an important role as the mediator of skeletal muscle damage after strenuous exercise and in aging (Jackson et al., 1993; Pansarasa et al., 1999) In muscle injury during extreme exercise, the generation of ROS and its toxic action has been attributed to the necrotic degeneration of muscle fibers (Davies et al., 1982). Animals deficient in the antioxidant vitamin E develop the necrotizing myopathy (Pillai et al., 1994).
Apoptosis, also known as programmed cell death, is a form of cell death that occurs during several pathological situations in multicellular organisms. In normal physiological condition, apoptosis contributes to cell replacement, tissue remodeling, and removal of damaged cells (Thompson, 1995; DeLong, 1998).
Numerous studies reported that the process of apoptosis is
regulated by the expression of several proteins that are classified into two functionally distinct groups: anti-apoptotic proteins and pro-apoptotic proteins. The anti-apoptotic protein bcl-2 is known to regulate the apoptotic pathway and to inhibit the progression of the apoptotic pathway, while the pro-apoptotic protein bax is expressed abundantly and selectively during apoptosis and pro- motes cell death (Oltvai et al.,1993; Korsmeyer, 1999).
N-acetylcysteine (NAC) is a small molecule containing a thiol group with the antioxidant property. NAC freely translocates in intracellular compartments (Holodines, 1991; De Vries et al., 1993). The diversity of pharmacological applications of NAC is due to the chemical property of the cysteinyl thiol group of its molecule. The reduced thiol group scavenges oxygen free radicals (Aruoma et al., 1989; Cuzzocrea et al., 2000).
NAC is well known antioxidant. However it has not been clearly established whether NAC exerts protective effects on ROS- induced apoptosis in muscles. Here, we examined the protective effect of NAC on the H2O
2-induced apoptosis in mouse C2C12 myoblast cells by applying 3-(4,5-dimethylthiazol-2-yl)-2,5-diphe- nyltetrazolium bromide (MTT) assay, 4,6-diamidino-2-pheny- lindole (DAPI) staining, terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling (TUNEL) assay, DNA fragmentation assay, flow cytometry, ROS detection assay, reverse transcription-polymerase chain reaction (RT-PCR), and Western blotting.
MATERIALS AND METHODS
1. Cell culture
C2C12 mouse myoblasts were cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco BRL, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco BRL) at 37°C in 5% CO
2, 95% O
2in a humidified cell incubator. The medium was changed every 2 days.
2. MTT cytotoxicity assay
Cell viability was determined using the MTT assay kit according to the manufacturer’s protocol (Boehringer Mannheim GmbH, Mannheim, Germany). Briefly, cells were treated with H
2O
2, added 10 μl MTT labeling reagent to each well, and incubated the plates for 4 h. Subsequently, 100 μl solubilization
solution was added to each well, incubated the plates for another 12 h, and the absorbance at the test wavelength 595 nm and the reference wavelength 690 nm was then measured with a microtiter plate reader (Bio-Tek, Winooski, VT, USA). The optical density (O.D.) was calculated as the difference between the absorbance at the reference wavelength and that at the test wavelength. The percent viability was calculated as follows: The percent viability=(O.D. of drug-treated sample/control O.D.)×100.
3. Morphological change
After treatment with H
2O
2, cells were washed three times in phosphate-buffered saline (PBS) and fixed with 100% methanol at
−20°C for 10 min. The cells were observed with a phase-cont- rast microscope (Olympus, Tokyo, Japan).
4. TUNEL assay
For in situ detection of apoptotic cells, TUNEL assay was performed using ApoTag
Ⓡperoxidase in situ apoptosis detection kit (Boehringer Mannheim GmbH) (Kim BS et al., 2001). Cells (2× 10
4cells/chamber) were cultured on 4-chamber slides (Nalge Nunc International, Naperville, IL, USA), washed with PBS, and fixed in 4% paraformaldehyde (PFA) for 10 min at 4°C. The fixed cells were incubated with digoxigenin-conjugated dUTP in a terminal deoxynucleotidyl transferase-catalyzed reaction for 1 h at 37°C in a humidified atmosphere, incubated with stop/wash buffer for 10 min at room temperature, and incubated with anti-digoxigenin antibody conjugated with peroxidase for 30 min.
The DNA fragments were stained using 3,3’-diaminobenzidine (Sigma Chemical Co., St. Louis, MO, USA) as the substrate for the peroxidase.
5. DAPI staining
To see whether H
2O
2induces apoptosis, DAPI staining was
performed as previously described (Jang MH et al., 2003). Briefly,
cells were cultured on 4-chamber slides, washed twice with PBS,
fixed by incubating with 4% PFA for 30 min, washed with PBS,
incubated with 1 μg/ml DAPI for 30 min in the dark, and
analyzed with a fluorescence microscope (Zeiss, Oberköchen, Ger-
many).
6. DNA fragmentation
DNA fragmentation assay was performed using ApopLadder EX
TMDNA fragmentation assay kit (TaKaRa, Shiga, Japan). Cells were treated with H
2O
2, lysed with 100 μl of lysis buffer, in- cubated with 10 μl of 10% sodium dodecyl sulfate (SDS) solu- tion containing 10 μl of Enzyme A at 56°C for 1 h, and then incubated with 10 μl of Enzyme B at 37°C for 1 h. This mixture was added with 70 μl of precipitant and 500 μl of ethanol and centrifuged for 15 min. DNA was extracted by washing the pellet in ethanol and resuspending it in Tris-EDTA (TE) buffer. DNA fragmentation was visualized by 2% agarose gel electrophoresis and staining with ethidium bromide.
7. Flow cytometric analysis
Flow cytometric analysis was performed as described (Kim EH et al., 2003). Cells were collected, fixed by incubating with 75%
ethanol in PBS at −20°C for 1 h, incubated with 100 μg/ml RNase and 20 μg/ml propidium iodide in PBS for 30 min at 37°C, and analyzed using FACScan (Becton Dickinson, San Jose, CA, USA).
8. Measurement of ROS generation
2’-7’-Dichlorofluorescin diacetate (DCFH-DA) allows to mea- sure intracellular ROS production by flow cytometry. Briefly, cells were suspended in PBS, incubated with 10 μM DCFH-DA at 37°C for 30 min, washed with ice-cold PBS, detached by trypsinization, resuspended in PBS, and analyzed more than 10,000 viable cells. To localize ROS, cells were cultured on 4-chamber slides, washed twice with PBS, incubated with 10 μM DCFH-DA at 37°C for 30 min, and analyzed through a fluo- rescence microscope (Zeiss).
9. RNA isolation and RT-PCR
Total RNA was isolated from C2C12 cells using easy-BLUE
TMtotal RNA extraction kit according to the manufacturer’s instruction (iNtRON, INC., Seoul, Korea). Two μg of RNA and 2 μl of random hexamers (Promega, Madison, WI, USA) were added together and the mixture was heated at 65°C for 10 min.
To the mixture, 1 μl of AMV reverse transcriptase (Promega), 5 μl of 10 mM dNTP (Promega), 1 μl of RNasin (Promega),
and 5 μl of 10×AMV RT buffer (Promega) were added and the final volume was adjusted to 50 μl with dimethyl pyrocarbonate (DEPC)-treated water. The reaction mixture was incubated at 42
°C for 1 h.
PCR amplification was performed in the reaction volume 40 μl containing 1 μl of the appropriate cDNA, 1 μl of each set of primers at the concentration 10 pM, 4 μl of 10×reaction buffer, 1 μl of 2.5 mM dNTP, and 2 units Taq DNA polymerase (TaKaRa). The primer sequences for bax were 5’-AGATG AACT GGATAGCAATATGGA-3’ (a 24-mer sense oligonucleotide star- ting at position 201) and 5’-CCACCCTGGTCTTGGATCCAG ACA-3’ (a 24-mer anti-sense oligonucleotide starting at position 446). The primer sequences for bcl-2 were 5’-TCCGTG CCTGACTTTAGCAAGCTG-3’ (a 24-mer sense oligonucleotide starting at position 741) and 5’-GGAATCCCAACCAGAGATCT CAA-3’ (a 24-mer anti-sense oligonucleotide starting at position 1051). The primer sequences for the internal control cyclophilin were 5’-ACCCCACCGTGTTCTTCGAC-3’ (a 20-mer sense oli- gonucleotide starting at position 52) and 5’-CATTTGCCATGGA CAAGATG-3’ (a 20-mer anti-sense oligonucleotide starting at position 332). The expected sizes of the PCR products were 270 bp for bax, 333 bp for bcl-2, and 291 bp for cyclophilin.
PCR procedure was carried out using a GeneAmp 9600 PCR system (Perkin Elmer, Norwalk, CT, USA) under the following conditions: initial denaturation at 94°C for 5 min, followed by 30 amplification cycles, each consisting of denaturation at 94°C for 30 sec, annealing at 58°C for 30 sec, and extension at 72°C for 30 sec, with an additional extension step at the end of the procedure at 72°C for 5 min. For cyclophilin, the PCR procedure was carried out under identical conditions except that 25 amplification cycles were performed. The final amount of RT-PCR product was calculated densitometrically using Molecular Analyst
TMversion 1.4.1 (Bio-Rad, Hercules, CA, USA).
10. Western blot analysis
Cells were lysed in the lysis buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.5% deoxycholic acid, 1%
nonidet-P40 (NP40), 0.1% SDS, 1 mM phenylmethylsulfonyl
fluoride (PMSF), and 100 μg/ml leupeptin. Protein concentration
was measured using a Bio-Rad colorimetric protein assay kit
(Bio-Rad). Protein of 50 μg was separated on SDS-polyacryla-
Fig. 1. Protectivie effect of N-acetylcysteine (NAC) on H2O2-induced cytotoxicity. Left: Cytotoxic effect of H2O2. ○: Incubation for 12 h. ●: Incubation for 24 h. Right: Protective effect of NAC. (A) Control cells, (B) 100 μM H2O2-treated cells, (C) 5 μM NAC followed by 100 μM H2O2-treated cells, (D) 10 μM NAC followed by 100 μM H2O2-treated cells, (E) 50 μM NAC followed by 100 μM H2O2-treated cells, (F) 100 μM NAC followed by 100 μM H2O2-treated cells. ap<0.05 compared to control cells, bp<0.05 compared to 100 μM H2O2-treated cells.
mide gels and transferred onto a nitrocellulose membrane (Schleicher & Schuell GmbH, Dassel, Germany). Mouse bax antibody (1:1,000; Santa Cruz Biotech, Santa Cruz, CA, USA), and rabbit bcl-2 antibody (1:1,000; Santa Cruz Biotech) were used as primary antibody. Horseradish peroxidase-conjugated anti-mouse antibody for bax (Amersham Pharmacia Biothech GmbH, Freiburg, Germany) and anti-rabbit antibody for bcl-2 (Amersham Pharmacia Biothech GmbH) were used as secondary antibody. Band detection was performed using the enhanced chemiluminescence (ECL) detection system (Amersham Pharmacia Biothech GmbH).
11. Statistical analyses
Results are expressed as the mean±standard error of the mean (S.E.M.). The data were analyzed by one-way ANOVA followed by Duncan’s post-hoc test using SPSS. Difference was considered statistically significant at p<0.05.
RESULTS
1. Effect of H
2O
2on the viability of C2C12 cells As shown in Fig. 1, the viability of cells incubated with H
2O
2for 12 h at the concentrations 10 μM, 50 μM, 100 μM, 500 μM, 1 mM, 5 mM, and 10 mM was 99.74±5.19%, 103.62±
6.20%, 78.24±5.87%, 67.94±2.42%, 38.67±0.93%, 9.51± 0.68%, and 6.19±5.24%, respectively. The viability of cells incubated with
H
2O
2for 24 h at the concentrations 10 μM, 50 μM, 100 μ M, 500 μM, 1 mM, 5 mM, and 10 mM was 88.96±6.52%, 73.84±4.44%, 53.84±3.21%, 41.64±2.34%, 27.40±0.11%, 5.79±
0.28%, 6.62±5.24%, respectively. As H
2O
2increased, the viabili- ty was decreased.
The viability of cells exposed to 100 μM H
2O
2for 24 h was 53.84±3.21%. The viability of cells treated with NAC for 1 h at the concentrations 5 μM, 10 μM, 50 μM, and 100 μM prior to the H
2O
2exposure was increased to 57.85±2.01%, 64.75±
0.81%, 77.95±0.49%, and 88.57±3.67%, respectively. The data demonstrate that the viability of H
2O
2-treated cells was decreased and NAC exerted the protective effect against the H
2O
2-induced cytotoxicity.
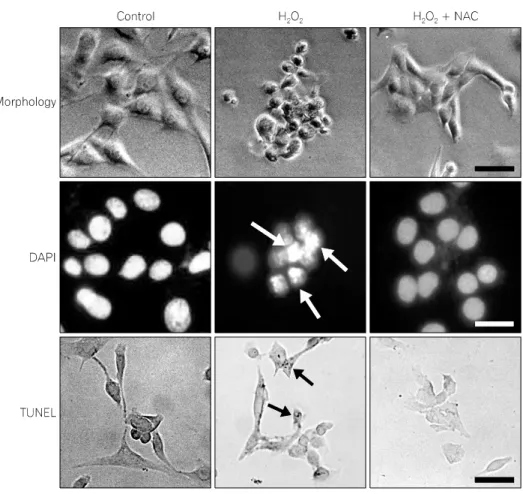
2. Morphological change induced by H
2O
2To characterize H
2O
2-induced change in cell morphology, cells were examined by phase-contrast microscopy. As shown in Fig. 2 (top), cells treated with 100 μM H
2O
2for 12 h detached from the culture dish, rounded, and became irregular shape with cytoplasmic blebbings. Cells treated with 100 μM NAC for 1 h prior to H
2O
2exposure were indistinguishable from normal cells.
In the DAPI assay, nuclear condensation, DNA fragmentation,
and perinuclear apoptotic bodies were detected in cells treated
with 100 μM H
2O
2for 12 h. Cells treated with 100 μM NAC
for 1 h prior to H
2O
2exposure were comparable to normal cells
(Fig. 2, middle).
Fig. 2. The effect of N-acetylcysteine (NAC) on H2O2-induced morphological change. Top: Phase-contrast photomic- rographs. Middle: C2C12 cells stained with DAPI. White arrows indicate condensed nuclei. Bottom: C2C12 cells analyzed by TUNEL assay. Black arrows show the labeling of condensed and marginated chromatin. The scale bar represents 100 μm.
Fig. 3. Electrophoretic examination of the genomic DNA of C2C12 cells.
(A) Control cells, (B) 100 μM H2O2-treated cells, (C) 100 μM N-ace- tylcysteine (NAC) followed by 100 μM H2O2-treated cells.
To confirm that H
2O
2induces apoptosis, cells treated with 100 μM H
2O
2were analyzed via TUNEL assay. As shown in Fig.
2 (bottom), TUNEL-positive cells were detected in cultures treated with H
2O
2. The number of TUNEL-positive cells in cultures treated with 100 μM NAC for 1 h prior to H
2O
2exposure was comparable to control cells without H
2O
2treat- ment.
3. Detection of apoptosis by examining DNA fragmentation
To further confirm that H
2O
2induces apoptosis, DNA fragmentation was analyzed. As shown in Fig. 3, DNA fragmentation was detected in cells treated with 100 μM H
2O
2for 12 h. In cells treated with 100 μM NAC for 1 h followed by H
2O
2, significantly decreased DNA laddering was detected.
4. Effect of H
2O
2on cell cycle
The population of cells in the sub-G1 phase in the cultures
Fig. 4. Flow cytometric analysis. The population of cells in the sub-G1 phase was increased in the cells treated with 100 μM H2O2 for 12 h and decreased by pre-treatment with 100 μM NAC for 1 h.
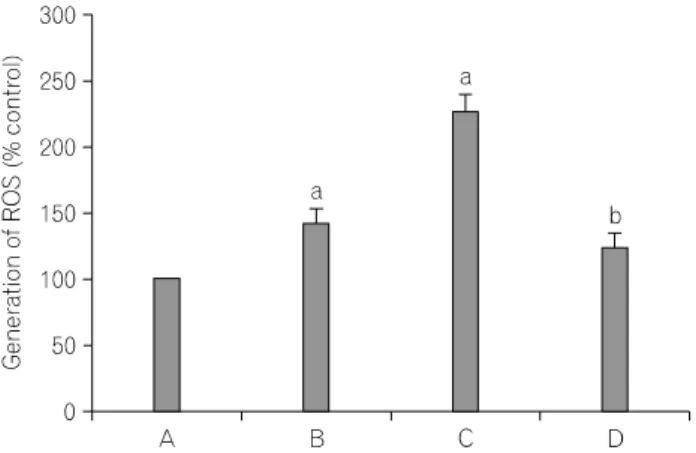
Fig. 5. Flow cytometric analysis of the generation of reactive oxygen species (ROS) in cells. Above: Fluorescence microscopic analysis of H2O2- induced ROS generation. Below: Protective effect of N-acetylcysteine (NAC) on ROS generation. (A) Control cells, (B) 50 μM H2O2-treated cells, (C) 100 μM H2O2-treated cells, (D) 100 μM NAC followed by 100 μM H2O2-treated cells, ap<0.05 compared to control cells, bp
<0.05 compared to 100 μM H2O2-treated cells.
treated with 100 μM H
2O
2for 12 h was increased from 23.27%
(control) to 85.48%, and decreased to 31.75% in cultures pre-treated with 100 μM NAC for 1 h (Fig. 4).
5. H
2O
2induces intracellular ROS generation To see whether H
2O
2induces the generation of ROS in cells, cells were treated with the 50 μM DCFH-DA for 30 min. The ROS level in cells incubated with H
2O
2for 12 h at the concentrations 50 μM and 100 μM was increased 142.52±
10.05% and 227.52±12.35%. The ROS level in cells treated with
100 μM NAC for 1 h followed by H
2O
2was decreased to 124.74±11.15% (Fig. 5).
6. Effect of H
2O
2on bax and bcl-2 mRNA level RT-PCR analysis of the mRNA levels of bax and bcl-2 was performed. Compared to the level of bax mRNA in control cells 1, in cells treated with H
2O
2at the concentrations 50 μM and 100 μM, the level of bax mRNA was markedly increased to 15.99±1.71 and 28.31±3.28, respectively. In cells treated with 100 μM NAC for 1 h followed by 100 μM H
2O
2, the level of bax mRNA was decreased to 3.63±1.08.
Compared to the level of bcl-2 mRNA in control cells 1, the level of bcl-2 mRNA was markedly decreased to 0.72±0.03 and 0.22±0.05 in cells treated with 50 μM and 100 μM H
2O
2, respectively. In cells pre-treated with 100 μM NAC and subsequently with 100 μM H
2O
2, the level of bcl-2 mRNA was increased to 0.73±0.05 (Fig. 6).
7. Western blot analysis of bax and bcl-2 pro- tein level
In cells treated with 100 μM H
2O
2, increased bax protein (21
kDa) and decreased bcl-2 protein (26 kDa) were detected. In cells
treated with 100 μM NAC for 1 h prior to H
2O
2exposure, the
expression of bax protein was decreased and bcl-2 protein was
increased (Fig. 7).
Fig. 6. Reverse transcription-polymerase chain reaction (RT-PCR) analysis of the mRNA levels of bax and bcl-2. As the internal control, cyclophilin was used. ap<0.05 compared to control cells, bp<0.05 compared to 100 μM H2O2-treated cells. (A) Control cells, (B) 50 μM H2O2-treated cells, (C) 100 μM H2O2-treated cells, (D) 100 μM N-acetylcysteine (NAC) followed by 100 μM H2O2-treated cells.
Fig. 7. Western blot analysis of bax and bcl-2. (A) Control cells, (B) 50 μM H2O2-treated group, (C) 100 μM H2O2-treated cells, (D) 100 μ M N-acetylcysteine (NAC) followed by 100 μM H2O2-treated cells.
DISCUSSION
The major ROS H
2O
2destroys cells by inducing apoptosis.
Such ROS may overwhelm cell’s natural antioxidant defense systems resulting in damaging all biological macromolecules such as nucleic acids, proteins, carbohydrates, and lipids. Such damage is implicated in several biological and pathological processes including aging, inflammation, and carcinogenesis (Pansarasa et al., 1999; Sandri et al., 1999). The increased level of H
2O
2was observed in skeletal muscles following excessive exercise or aging.
H
2O
2generation induces apoptosis in skeletal muscle cells (Dirks et al., 2002; Silveira et al., 2003). NAC may exert its antioxidant effect indirectly by facilitating the biosynthesis and supplying glutathione to glutathione peroxidase-catalysed reactions. NAC has been used for many disease states such as paracetamol toxicity, pulmonary oxygen toxicity, and human immune- deficiency virus infection, ischemic damage, the toxicity in the lung and liver, and experimental ischemic skin flaps (Moldeus et al., 1986; Knight et al., 1991; Sener et al., 2003). In addition, it was reported that antioxidant deficiency increases free- radical-induced tissue injury (Sacheck et al., 2001). Servais et al.(2003) reported that H
2O
2release was increased in subsar- colemmal and intermyofibrillar mitochodria after prolonged wheel exercise.
Our data confirmed that H
2O
2exerts cytotoxic effect in a
dose-dependent fashion and NAC protects cells from the cytotoxic
action of H
2O
2(Fig. 1). H
2O
2induced cell shrinkage, cytoplasmic
condensation, and the irregularity in shape (Fig. 2). Apoptotic
bodies, the morphological criteria of apoptosis, were detected in
H
2O
2-treated cells stained with DAPI. H
2O
2induced the changes
characteristic to apoptosis in C2C12 cells, and NAC attenuated
such change. In addition, TUNEL-positive cells, indicative of
apoptotic DNA strand breaks and it is known that nicks in the
DNA molecules can be detected via TUNEL assay (Jang MH et
al., 2002), were detected in cultures treated with H
2O
2. TUNEL-
positive cells were decreased in cultures pre-treated with NAC
(Fig. 2). The flow cytometric analysis of DNA content revealed
the increase of the fraction of cells in the sub-G1 phase in
cultures treated with H
2O
2. The increase of the sub-G1-phase
fraction was reduced by the pre-treatment with NAC (Fig. 3).
Apoptosis involves the activation of endonucleases resulting in the cleavage of genomic DNA into well-defined fragments that appear as the characteristic ladder pattern separated by agarose gel electrophoresis (Eastman et al., 1992). To provide evidence of the involvement of apoptosis in the H
2O
2-induced cytotoxicity and the protective action of NAC against the H
2O
2-induced apoptosis, DNA fragmentation assay was performed. In the present results, the distinctive characteristic of apoptosis DNA ladder pattern was detected in cells treated with H
2O
2. Cells pre-treated with NAC showed the noticeable decrease in the intensity of H
2O
2-induced DNA laddering (Fig. 4).
The bcl-2 family proteins are characterized by their ability to form a heterodimer with bax and a homodimer with itself (Reed, 1998). When the pro-apoptotic protein bax is overexpressed, apoptotic death in response to death signals is accelerated. When bcl-2 is overexpressed, it heterodimerizes with bax and cell death is repressed. The ratio of bax and bcl-2 may determine the susceptibility of cells to apoptosis (Korsmeyer, 1999). Our data showed that H
2O
2treatment resulted in the increase in the bax expression and the remarkable decrease in the bax level by NAC pre-treatment. In addition, we have shown the down-regulation of the bcl-2 level in cells exposed to H
2O
2. The remarkable increase in the bcl-2 level was observed in cells pre-treated with NAC.
Here, we show that H
2O
2induces apoptosis in myoblast C2C12 cells and NAC protects them from the H
2O
2-induced apoptosis through the regulation of the expression of bax and bcl-2. The present study suggests that NAC may offer a mean to treat the oxidative stress-induced skeletal muscle damage.
ACKNOWLEDGEMENTS
This research was supported by the Kyungwon University Research Fund in 2009.
REFERENCES
Aruoma OI, Halliwell B, Hoey BM et al. (1989) The antioxidant action of N-acetylcysteine: Its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic.
Biol. Med. 6:593-597.
Cuzzocrea S, Mazzon E, Costantino G et al. (2000) Beneficial effects of n-acetylcysteine on ischaemic brain injury. Br. J. Pharmacol.
130:1219-1226.
Davies KJ, Quintanilha AT, Brooks GA et al. (1982) Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res.
Commun. 107:1198-1205.
DeLong MJ (1998) Apoptosis: a modulator of cellular homeostasis and disease states. Ann. N. Y. Acad. Sci. 842:82-90.
De Vries N, De Flora S (1993) N-acetyl-l-cysteine. J. Cell. Biochem.
Suppl. 17F:270-277.
Dirks A, Leeuwenburgh C (2002) Apoptosis in skeletal muscle with aging. Am. J. Physiol. Regulatory Integrative Comp. Physiol. 282:
R519-R527.
Eastman A, Barry MA (1992) The origins of DNA breaks: a consequence of DNA damage, DNA repair, or apoptosis? Cancer Invest. 10:229-240.
Holodines MR (1991) Clinical pharmacokinetics of N-acetylcysteine Clin. Pharmacokinet. 20:123-134.
Jackson MJ, O’Farrell SO (1993) Free radicals and muscle damage Br. Med. Bull. 49:630-641.
Jang MH, Lee TH, Shin MC et al. (2002) Protective effect of Hypericum perforatum Linn (St. Johnn’s wort) against hydrogen peroxide-induced apoptosis on neuroblastoma cells. Neurosci. Lett.
329:177-180.
Jang MH, Shin MC, Lim S et al. (2003) Bee venom induces apoptosis and inhibits expression of cycooxygenase-2 mRNA in human lung cancer cell line NCI-H1299. J. Pharmacol. Sci.
91:95-104.
Kanno S, Ishikawa M, Takayanagi M et al. (1999) Exposure to hydrogen peroxide induces cell death via apoptosis in primary cultured mouse hepatocytes. Biol. Pharm. Bull. 22:1296-1300.
Kim BS, Kim SA, Kim MJ et al. (2001) Indomethacin induces apoptosis in NCI-H1299 human lung carinoma cells. Korean J.
Physiol. Pharmacol. 5:177-181.
Kim EH, Jang MH, Shin MC et al. (2003) Protective effect of aqueous extract of Ginseng radix against 1-methyl-4-phenylpyri- dinium-induced apoptosis on PC12 cells. Biol. Pharm. Bull. 26:
1668-1673.
Knight KR, Mac Phadyen K, Lepore DA et al. (1991) Enhancement of ischaemic rabbit skin flap survival with the antioxidant and free-radical scavenger N-acetylcysteine. Clin. Sci. 81:31-36.
Korsmeyer SJ (1999) bcl-2 gene family and the regulation of programmed cell death. Cancer Res. Suppl. 59:1693s-1700s.
Liu XL, Sato S, Dai W et al. (2001) The protective effect of hepatocyte growth-promoting factor (pHGF) against hydrogen peroxide-induced acute lung injury in rats. Med. Electron. Microsc.
34:92-102.
Moldeus P, Cotgreave IA, Berggren M (1986) Lung protection by a thiol-containing antioxidant: N-acetylcysteine. Respiration 50:31-42.
Oltvai ZN, Milliman CL, Korsmeyer SJ (1993) bcl-2 heterodimerizes in vivo with a conserved homolog, bax, that accelerates pro-
= 국문초록 =
반응성 산화물은 심한 운동이나 노화에 의한 골격근 손상에 중요한 역할을 한다. 본 실험에서는 항산화제인 N-acetylcysteine이 C2C12 근세포에서 과산화수소에 의한 세포독성에 대한 보호작용을 실험하였다. C2C12 근세포에서 과산화수소는 세포사멸을 유발하였고, N-acetylcysteine을 전처치 시 과산화수소에 의한 세포사멸이 감소되었다. 또한 과산화수소는 세포사멸 유전자인 bax를 증가시켰고, 세포보호 유전자인 bcl-2를 감소시켰다. N-acetylcysteine으로 전처 치 시 bax의 발현은 감소되었고 bcl-2의 발현은 증가되었다. 본 실험을 통하여 N-acetylcysteine은 근육세포에서 bax와 bcl-2의 발현을 조절함으로써 과산화수소에 의한 세포사멸에 보호작용을 나타냄을 알 수 있었다.
중심단어: N-acetylcysteine, 반응성 산화물, 과산화수소, 골격근 세포, 세포사멸
grammed cell death. Cell 74:609-619.Pansarasa O, Bertorelli L, Vecchiet J et al. (1999) Age-dependent changes of antioxidant activities and markers of free radical damage in human skeletal muscle. Free Radic. Biol. Med.
27:617-622.
Pillai SR, Traber MG, Kayden HJ et al. (1994) Concomitant brainstem axonal dystrophy and necrotizing myopathy in vitamin E-deficient rats. J. Neurol. Sci. 123:64-73.
Ratan RR, Murphy TH, Baraban JM (1994) Oxidative stress induces apoptosis in embryonic cortical neurons. J. Neurochem. 62:376- 379.
Reed JC (1998) bcl-2 family proteins. Oncogene 17:3225-3236.
Sacheck JM, Blumberg JB (2001) Role of vitamin E and oxidative stress in exercise. Nutrition 17:809-814.
Sandri M, Carraro U (1999) Apoptosis of skeletal muscles during development and disease. Int. J. Biochem. Cell Biol. 31:1373- 1390.
Sener G, Tosun O, Şsehirli Ö et al. (2003) Melatonin and N-acetyl- cysteine have beneficial effects during hepatic ischemia and reperfusion. Life Sci. 72:2707-2718
Servais S, Couturier K, Koubi H et al. (2003) Effect of voluntary exercise on H2O2 release by subsarcolemmal and intermyofibrillar mitochondria. Free Radic. Biol. Med. 35:24-32.
Silveira LR, Pereira-Da-Silva L, Juel C et al. (2003) Formation of hydrogen peroxide and nitric oxide in rat skeletal muscle cells during contractions. Free Radic. Biol. Med. 35:455-464.
Thompson CB (1995) Apoptosis in the pathogenesis and treatment of disease. Science 267:1456-1462.