위령선 추출물이 Human Neuroblastoma 세포주에서 CoCl 2 에 의해 유도된 세포사멸에 미치는 보호효과
박정우⋅임형호
가천대학교 한의과대학 한방재활의학과교실
Protective Effect of Clematidis Radix Extract on CoCl 2 -induced Apoptosis in Human Neuroblastoma Cells
Jung-Woo Park, K.M.D., Hyung-Ho Lim, K.M.D.
Department of Korean Medicine Rehabilitation, College of Korean Medicine, Gachon University
이 연구는 2013년도 가천대학교 지원에 의한 결과임.
RECEIVED March 18, 2014 REVISED March 28, 2014 ACCEPTED March 28, 2014
CORRESPONDING TO
Hyung-Ho Lim, Department of Korean Medicine Rehabilitation, College of Korean Medicine, Gachon University, 1200-1, Guwol-dong, Namdong-gu, Incheon 405-835, Korea
TEL (070) 7120-5011 FAX (032) 468-4033 E-mail [email protected]
Copyright © 2014 The Society of Korean Medicine Rehabilitation
Objectives The purpose of this study was to evaluate the effects of Clematidis radix ex- tract on CoCl
2-induced apoptosis in SH-SY5Y human neuroblastoma cells.
Methods In order to investigate the protective effect of Clematidis radix on CoCl
2-induced cytotoxicity in neuronal cells, MTT(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro- mide) assay, DAPI(4,6-diamidino-2-phenylindoleI) staining, TUNEL (terminal deoxynucleo- tidyl transferase-mediated dUTP nick end-labeling) assay, DNA fragmentation assay and western blotting were performed on SH-SY5Y human neuroblastoma cells.
Results Cells treated with CoCl
2exhibited several apoptotic features, while cells pre-treat- ed with Clematidis radix prior to CoCl
2exposure showed a decrease in the occurrence of apoptotic features. CoCl
2increased HIF-1α expression, in contrast, Clematidis radix treat- ment decreased CoCl
2-induced HIF-1α expression. Pre-treatment with the extract of Clematidis radix suppressed Bax, cytochrome c, and caspase-3 expressions, and also in- creased Bcl-2 expression in SH-SY5Y human neuroblastoma cells.
Conclusions These results suggest that Clematidis radix may exert a protective effect on CoCl
2-induced apoptosis in SH-SY5Y human neuroblastoma cells. (J Korean Med Rehab 2014;24(2):41-50)
Key words Clematidis radix , CoCl
2, Hypoxia, Apoptosis, SH-SY5Y human neuro- blastoma cells
서론»»»
위령선(
Clematidis radix
)은 전통적으로 신경통, 근육 통, 편두통과 부종의 치료제로 사용되어온 약물로, 성분 으로는 saponin, clematoside A, anemonol 등이 동정되 어 있다1). 위령선의 선행 연구에서는 관절염에서의 소염, 진통, 해열 작용2), 그리고 면역증강 작용3), 류마티스성 관 절염 억제 작용4), 통증 조절 작용5), 골다공증에서 파골세포 억제 작용6) 등이 보고되고 있다.
저산소증은 세포나 조직 내 산소 농도가 무산소증과 정상 수준 사이에 있는 대사 상태로, 뇌졸중 및 퇴행성 뇌질환의 신경손상을 관찰하고 그 치료법을 연구하는 과 정에 있어 중요한 요소이다7,8). 최근 연구에 의하면 CoCl2
(Cobalt chloride) 처치가 저산소 상태를 유도할 뿐만 아 니라 DNA 손상을 유도하고 세포의 DNA 손상 반응을 활 성화시킨다고 보고되었다9).
세포의 저산소 상태가 장시간 지속되거나 그 정도가 매우 극심한 경우, 세포는 사멸하게 된다. 이로 인하여 HIF-1 (hypoxia-inducible factor-1) 등의 단백질 발현이 증가되어 세포사를 유도할 수 있다7). HIF-1은 120 kDa의 HIF-1α와 91, 93, 94 kDa의 HIF-1β의 두 subunits으로 구성되어 있는데, 이중 HIF-1α가 저산소 상태에 의해 조 절되는 주요한 subunit이라 한다10). HIF-1α는 저산소 상 태에서는 외부 산소농도 변화에 적절히 반응하기 위해 세 포내의 항상성을 유지시켜주지만 지나친 HIF-1α의 발현 은 세포사를 유도하는 신호 체계를 활성화 시킨다10).
세포사멸에 대한 보호효과에 관한 연구로는 시호11), 오 가피12), 정공피13), 인삼14), 봉약침15) 등을 이용한 연구가 보고되어 있다. 그러나 저산소성
in vitro
모델에서 위령 선의 세포사멸에 대한 보호효과에 관한 연구는 미흡하다.이에 본 연구에서는 위령선의 세포 사멸에 대한 보호효과 에 대하여 알아보기 위하여, CoCl2에 의해 유발된 저산소 성
in vitro
모델에서 위령선 추출물 처치가 인간 신경아 세포종 SH-SY5Y의 HIF-1α와 세포사멸에 미치는 영향을 조사하였다.재료 및 방법»»»
1. 재료
1) 세포배양
인간 신경아세포종(SH-SY5Y)는 한국 세포주은행(KCLB, Seoul, Korea)으로부터 구입하였다. SH-SY5Y를 10% fe- tal bovine serum (FBS, Gibco-BRL, Gaithersburg, MD, USA)을 넣은 Dulbecco’s Modified Eagle Medium (DMEM, Welgene, Daegu, Korea) 배양액으로 5% CO2, 95% O2가 공급되는 배양기에서 37oC 조건으로 배양하였으며, 배지 교체는 2일마다 하였다.
2) 시료준비
본 실험에 사용할 위령선은 위령선 50 g을 1 L의 3차 증류수에 넣어 90oC에서 2시간 동안 추출하였고, rotary evaporator (Eyela, Tokyo, Japan)를 사용하여 농축하였 다. 그리고 농축된 위령선을 12시간 동안 동결건조 (Eyela)하여 6.94 g (수율 14%)의 시료를 얻었다.
3) 실험군 분류
농도에 따른 CoCl2의 세포독성을 확인하기 위해 세포 생존율 측정시에는 100μM, 200μM, 300μM, 400μM, 500μM의 CoCl2를 이용하였으며, 나머지 실험에서는 MTT 분석을 통해 결정된 100μM의 CoCl2를 선택하여 사용하였다.
농도에 따른 위령선 추출물의 효과를 알아보기 위해 0.1μg/ml, 1μg/ml, 10μg/ml, 100μg/ml, 1,000μg/ml 의 농도로 처치군을 나누어 세포에 처치를 하고 100μM 의 CoCl2를 처치하였으며, 대조군으로는 아무런 처치를 하지 않은 군을 사용하였다.
2. 실험
1) MTT 분석
MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazo- lium bromide) cytotoxicity assay는 세포배양에서 생존하 는 세포의 수를 측정하는 방법으로 살아있는 세포의 mi- tochondrial dehydrogenase가 기질 MTT를 검푸른 색깔 의 formazan으로 변환시키는 작용을 이용한 것이다. 흡 광도(OD; optical density)의 값은 살아있는 세포의 수를 반영하므로 세포생존 지수는 다음의 산출식에 따라 %로 나타내었다.
Cell viability (%)=
Optical density of treated group Optical density of control group×100
세포를 10% FBS가 함유된 DMEM 용액에서 5%
CO2-95% O2, 37oC 온도 조건이 유지되는 세포 배양기에 서 배양하였다. 그리고 96 well plate에 well당 2.0×104 의 세포수가 되도록 분주하고 배양액 100μl를 첨가하였 다.
농도에 따른 CoCl2의 세포독성을 결정하기 위하여 세 포에 각각 100μM, 200μM, 300μM, 400μM, 500μM 의 CoCl2 처치를 하여 12시간 동안 배양하였다. 별도로 위령선 추출물의 농도에 따른 세포 생존율의 변화를 확인 하기 위해 세포에 각각 0.1μg/ml, 1μg/ml, 10μg/ml, 100μg/ml의 위령선 추출물 처치를 하고 1시간 후에 100 μM CoCl2 처치를 하여 12시간 동안 배양하였다.
각각에 MTT 용액을 50μl 가한 뒤 알루미늄 호일로 덮어 빛에 노출되지 않게 하여 4시간 동안 배양하였다.
Solubilization solution 100μl를 첨가하여 overnight re- action 후 enzyme-linked immunosorbent assay (ELISA) reader (Bio-Rad, Hercules, CA, USA)로 595 nm와 690 nm에서 흡광도를 측정하였다.
2) DAPI 염색
DAPI(4ʹ,6-diamidino-2-phenylindole) 염색은 핵 DNA 의 dsDNA에 특이적으로 결합하여 핵분절, 핵응축 등 세 포사멸에 의한 핵의 형태적인 변화를 관찰하는 데에 사용 된다. 위령선 추출물과 100μM CoCl2 처치 후 4% paraf- ormaldehyde로 30분간 고정화시켰다. 이어 phosphate buffer saline (PBS)에 wash 후 1μg/ml DAPI (Sigma- Aldrich Co., St. Louis, MO, USA) 용액과 37oC에서 30분 간 반응시켰다. DAPI 염색 시약 처리가 끝난 후 PBS로 30분간 마지막 wash를 하고 slide glass 위에 mounting solution을 이용하여 cover-slip을 붙인 후 형광현미경 (Nikon Eclipse 50i, Nikon Inc., Melville, NY, USA)으로 관찰하였다.
3) TUNEL 염색
세포사멸의 대표적 현상인 DNA 분절화의 발생은 in situ cell death detection kit (Roche Diagonostics GmbH, Mannheim, Germany)를 TUNEL (Terminal de- oxynucleotidyl transferase-mediated dUTP nick end -labeling) 염색을 이용하여 분석하였다. SH-SY5Y 세포주 를 Lab-Tek chamber slide(Nalge Nunc International, Rochester, NY, USA)에 분주하고 37oC, 5% CO2 배양기 에서 overnight reaction 후 100μM CoCl2 처치 1시간 전 에 위령선 추출물 농도를 각각 10μg/ml, 100μg/ml로 처치하고 12시간 동안 배양하였다. 4% paraformalde- hyde (pH 7.4)를 첨가하여 상온에서 20분간 고정한 후, PBS로 2회 세척하고, 0.1% Triton X-100, 0.1% sodium citrate가 포함된 permeabilization 용액으로 4oC에서 2분 간 처치하였다.
PBS로 다시 2회 세척한 후, terminal deoxynucleotidyl transferase (TdT)와 nucleotide mixture를 첨가하고 37oC 에서 빛을 차단한 후 60분간 배양하였다. PBS로 3회 세 척하고 난 후 현미경(Olympus, Tokyo, Japan)으로 세포
사멸을 관찰하였다.
4) PI 염색
SH-SY5Y 세포를 Lab-Tek chamber slide (Nalge Nunc International)에 분주하고 37oC, 5% CO2 배양기에서 배 양한 후 100μM CoCl2 처치 1시간 전에 위령선 추출물 농도에서 각각 10μg/ml, 100μg/ml로 처치하고 12시간 동안 배양하였다. 처치 종료 30분 전에 5 mg/ml PI (pro- pidium iodide; Sigma-Aldrich Co., St. Louis, MO, USA) 로 37oC에서 빛을 차단한 후 배양하였다. 배양 후 4%
paraformaldehyde (pH 7.4)를 첨가하여 상온에서 20분간 고정한 후, PBS로 3회 세척하고 난 후, 형광현미경(Nikon Inc.)으로 관찰하였다.
5) DNA fragmentation 분석
세포사멸 DNA cleavage를 조사하기 위한 DNA frag- mentation 분석은 ApopLadder ExTM DNA fragmentation assay kit (Takara, Shiga, Japan)를 사용하였다. 100μM CoCl2 처치 1시간 전에 위령선 추출물을 각각 100μg/ml, 1,000μg/ml로 처치하고 12시간 동안 배양하였다. 100μl 의 lysis buffer를 첨가한 후 10μl enzyme A를 포함한 10% SDS 10μl를 가지고 1시간 동안 56oC에서 배양한 후 10μl enzyme B를 추가하여 37oC에서 1시간 동안 배 양하였다. 그리고 침강제 70μl를 추가한 후 pellet를 얻 어 2% agarose gel에 전기영동 하였다.
6) Caspase-3 활성도 측정
100μM CoCl2 처치 1시간 전에 위령선 추출물 농도를 각각 100μg/ml, 1,000μg/ml로 처치하고 12시간 동안 배양한 후 세포를 모아서 용해시킨 후 용해한 세포를 얼 음에 10분 동안 두었다가 12,000 rpm으로 10분간 4oC에 서 원심 분리하여 상층액을 이용하여 정량하였다. 정량값 을 동일하게 맞춘 시료에 2X reaction buffer와 1 mM di- thiothreitol (DTT)을 첨가하여 얼음에서 30분 동안 반응 시키고, 1 mM caspase-3 substrate (DEVD-pNA)를 5μl 첨가하여 37oC 수조에서 1시간 동안 반응시킨 후 405 nm에서 흡광도를 측정하였다.
7) Western blot 분석
100μM CoCl2 처치 1시간 전에 위령선 추출물 농도를
각각 100μg/ml, 1,000μg/ml로 처치하고 12시간 동안 배양한 후 SH-SY5Y 세포를 PBS로 washing 후 extraction buffer (50 mM HEPES, pH 7.5, 150 mM NaCl, 10%
glycerol, 1% Triton X-100, 1.5 mM magnesium chloride hexahydrate, 1 mM ethyleneglycol-bis-(β-aminoethyl ether)-N,N'-tetraacetic acid, 1 mM phenylmethyl sulfon- yl fluoride, 2μg/ml leupeptin, 1μg/ml pepstatin, 1 mM sodium ortho vanadate, and 100 mM sodium flo- ride)로 세포를 lysis 시켰다. 10분간 ice에 방치 후 4oC에 서 14,000 rpm으로 15분간 원심분리 하였다. 상층액을 새로운 tube에 옮겨 단백질을 정량 후 사용하였다. 단백 질의 정량은 Bio-Rad protein assay kit (Bio-Rad)를 사용 하였다. SH-SY5Y 세포주에 lysis buffer를 넣고 4oC에서 30분간 얼음에 방치하고 12,000 rpm으로 15분간 원심 분 리하여, 그 상층액을 실험에 사용하였다. 추출된 단백질 30μg을 loading buffer와 혼합하여 10분간 끓이고, 10%
SDS-polyacrylamide gel에서 전압 70 V로 전기영동 시켰 다. 전기영동 후 단백질을 transfer buffer를 이용하여 260 mA로 4oC에서 90분 동안 nitrocellulose membrane (Whatman, Clifton, NJ, USA)에 옮긴 후, 5% skim milk (Becton Dickinson, Sparks, MD., USA)으로 4oC에서 overnight reaction 시키고 Tris buffered saline with Tween-20으로 세척하였다. 1차 항체를 TBS-T buffer에 희석하여 실온에서 2시간 반응시키고, tris-buffered saline tween-20 (TBST) buffer로 3회 세척하고, horseradish peroxidase 결합 2차 항체(Vector Laboratories, Burlinga- me, CA, USA)를 희석하여 실온에서 1시간 반응시켜 en- hanced chemiluminescence (ECL) kit (Santa Cruz Bio- technology Inc, Santa cruz CA, USA)를 이용하여 필름에 현상하였다. 1차 항체는 세포사멸과 관련된 단백질들에 대한 항체로서 anti-HIF-1α antibody (Novus Biologicals, Littleton, CO, USA), anti-Bcl-2 antibody (Santa Cruz Biotechnology Inc.), anti-Bax antibody (Santa Cruz Bio technology Inc.), cytochrome c (Santa Cruz Biotechnol- ogy Inc.)를 각각 사용하였다. 그리고 단백질 발현정도는 Image-ProⓇPlus software (Media Cybernetics Inc., Silver Spring, MD, USA)를 사용하여 정량화하였다.
3. 통계처리
통계처리는 SPSS-statistical software version 21.0 (SPSS Inc., Chicago, Illinois, USA)을 사용하였다. 측정치는 mean±standard error로 표시하였다. 그룹 간 비교를 위 해 one-way ANOVA 분석과 사후검증으로 Duncan's multiple range test를 사용하였으며, 모든 통계적 유의 수준은 p<0.05로 설정하였다.
결과»»»
1. 세포 생존율 측정
CoCl2를 100μM, 200μM, 300μM, 400μM, 500μM 농도에서 12시간 동한 처치 한 결과 세포 생존율은 각각 대조군과 비교했을 때 54.86±2.49%, 48.05±1.26%, 46.39±2.05%, 40.99±1.29%, 39.21±1.61%로 농도 의존 적으로 감소하였다(Fig. 1-1).
본 실험에서 선택된 100μM CoCl2 처치를 한 경우 세 포 생존율은 54.86±2.49%로 나타났고, 여기에 위령선 추출물 0.1μg/ml, 1μg/ml, 10μg/ml, 100μg/ml, 1,000 μg/ml을 처치하였을 때 세포 생존율은 각각 48.37±3.63%, 55.55±4.38%, 54.93±3.46%, 67.03±3.37%, 70.18±5.25%
로 농도 의존적으로 증가하였다(Fig. 1-2).
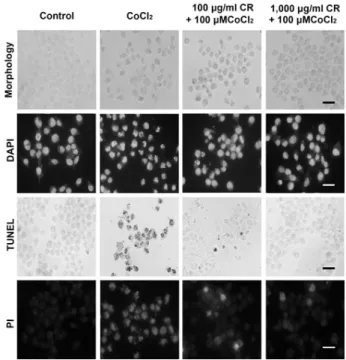
2. 세포의 형태학적 변화에 미치는 영향
세포의 형태학적 변화를 관찰한 결과 세포에 대한 100 μM CoCl2 처치 후 응축된 모양의 세포체가 나타났으며, 위령선 추출물로 처치에 의해서 응축된 세포체의 수가 줄 어드는 것으로 나타났다. DAPI 염색에서 밝은 푸른색을 나타내는 사멸된 핵은 염색체 응축과 사멸체(apoptotic body)에 의해 쉽게 구별할 수 있다. 100μM CoCl2 처치 후 핵의 염색체 응축이 나타났으며, 위령선 처치군에서는 100μM CoCl2 처치에 의해 유도된 염색체 응축이 억제되 어 나타났다. TUNEL 양성세포수는 100μM CoCl2 처치 군에서 증가하였으며, 위령선을 처치한 세포에서는 증가 가 억제되는 양상을 보였다. PI 염색에서 100μM CoCl2
처치에 의해 핵조각과 응축된 형태의 핵이 증가하였으며,
Fig. 1-1. CoCl
2-induced cytotoxicity.
A: Control group, B: 100μM CoCl
2-treated group, C: 200μM CoCl
2-treated group, D: 300μM CoCl
2-treated group, E: 400μ M CoCl
2-treated group, F: 500μM CoCl
2-treated group.
Results are presented as mean±standard error of the mean (S.E.M.).
*represents p<0.05 compared to the control group.
†
†
Fig. 1-2. Protective effect of Clematidis Radix on CoCl
2-induced cytotoxicity.
A: Control group, B: 100μM CoCl
2-treated group, C: 0.1μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, D: 1μg/ml Clematidis Radix -pretreated and 100μM CoCl
2- treated group, E: 10μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, F: 100μg/ml Clematidis Radix - pretreated and 100μM CoCl
2-treated group, G: 1,000μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group.
Results are represented as the mean±standard error of the mean (S.E.M.).
*represents p<0.05 compared to the control group.
†repre- sents p<0.05 compared to the 100μM CoCl
2-treated group.
Fig. 2. Morphological observations of cells treated with CoCl
2and Clematidis Radix (CR).
The scale bar represents 100μm.
위령선 처치에 의해 핵 조각의 증가가 억제되고 응축되는 핵의 형태학적 변화의 개선이 관찰되었다(Fig. 2).
3. DNA 분절화에 미치는 영향
세포의 DNA 분절화를 유도하는지 확인하기 위하여 100μM CoCl2 처치 1시간 전에 위령선 추출물 농도를 각 각 10μg/ml, 100μg/ml로 처치하고 12시간 동안 배양한 뒤 DNA를 추출하여 전기영동을 수행하였다. 실험 결과 CoCl2 처치 후 DNA 분절현상이 나타났으나 위령선 추출 물 처치에 의해 DNA laddering의 강도가 감소되었다 (Fig. 3).
4. HIF-1α 발현에 미치는 영향
HIF-1α 발현은 대조군에서의 HIF-1α(105 kDa) 발현 을 1.00으로 하였을 때, 100μM CoCl2 처치군에서 2.68±0.09로 나타났다. 위령선 추출물을 100μg/ml, 1,000μg/ml로 전처치 하였을 때의 HIF-1α 발현은 각각 2.08±0.28, 1.81±0.19로 나타났다. 100μM CoCl2 처치 군은 대조군과 비교해 HIF-1α 발현이 현저히 증가하였 으나 위령선 추출물 처치에 의하여 그 증가가 유의하게 억제되었다(Fig. 4).
†
†
†
†
† †
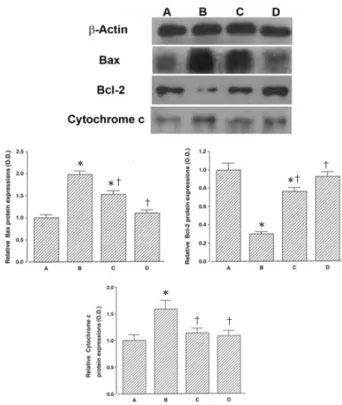
Fig. 5. Effects of Clematidis Radix on the expression of Bax, Bcl-2, cytochrome c proteins.
Upper: Photomicrographs of Bax, Bcl-2, and cytochrome c proteins. Middle left: Relative Bax protein expression (O.D.).
Middle right: Relative Bcl-2 protein expression (O.D.). Lower:
Relative cytochrome c protein expression (O.D.). A: Control group, B: 100μM CoCl
2-treated group, C: 100μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, D: 1,000μg/
ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group.
Results are represented as the mean±standard error of the mean (S.E.M.).
*represents p<0.05 compared to the control group.
†repre- sents p<0.05 compared to the 100μM CoCl
2-treated group.
Fig. 3. Electrophoretic examination of the genomic DNA of SH-SY5Y human neuroblastoma cells treated with CoCl
2and Clematidis Radix .
A: Control group, B: 100μM CoCl
2-treated group, C: 100 μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, D: 1,000μg/ml Clematidis Radix -pretreated and 100 μM CoCl
2-treated group.
†
†
Fig. 4. Effects of Clematidis Radix on the expression of HIF-1α protein.
A: Control group, B: 100μM CoCl
2-treated group, C: 100μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, D: 1,000μg/ml Clematidis Radix -pretreated and 100μM CoCl
2- treated group.
Results are represented as the mean±standard error of the mean (S.E.M.)
*represents p<0.05 compared to the control group.
†repre- sents p<0.05 compared to the 100μM CoCl
2-treated group.
5. Bax, Bcl-2, cytochrome c 발현에 미치는 영향
Bax (24 kDa)는 대조군에서의 발현을 1.00으로 하였 을 때, 100μM CoCl2 처치군의 발현이 1.98±0.07로 나 타났다. 위령선 추출물을 100μg/ml, 1,000μg/ml로 전 처치 하였을 때의 Bax 발현은 각각 1.53±0.08, 1.10±0.07로 감소하였다. 100μM CoCl2 처치군은 대조 군에 비해 Bax 발현이 현저히 증가하였으나 위령선 추출 물 처치에 의하여 그 증가가 유의하게 억제된 것으로 나 타났다(Fig. 5).
Bcl-2 (26 kDa)는 대조군에서의 발현을 1.00으로 하였 을 때, 100μM CoCl2 처치군의 발현이 0.30±0.02, 100 μg/ml로 나타났다. 위령선 추출물을 100μg/ml, 1,000
†
†
†
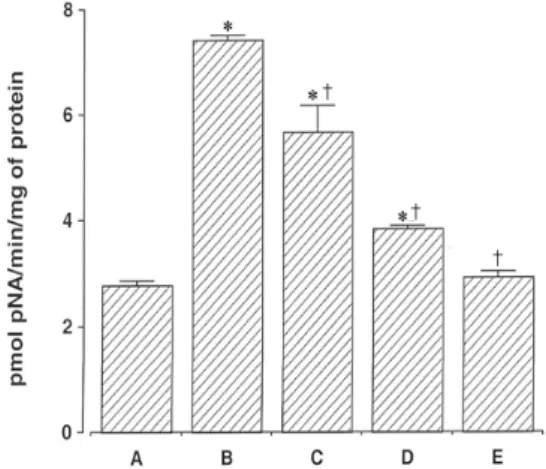
Fig. 6. Inhibitory effect of Clematidis Radix extract on CoCl
2-induced caspase-3 enzyme activity.
The rate of DEVD- p NA cleavage was measured at a wave- length of 405 nm.
A: Control group, B: 100μM CoCl
2-treated group, C: 100μg/ml Clematidis Radix -pretreated and 100μM CoCl
2-treated group, D: 1,000μg/ml Clematidis Radix -pretreated and 100μM CoCl
2- treated group, E: 100μM CoCl
2-treated group with DEVD-FMK added.
Results are represented as the mean±standard error of the mean (S.E.M.).
*represents p<0.05 compared to the control group.
†repre- sents p<0.05 compared to the 100μM CoCl
2-treated group.
μg/ml로 전처치 하였을 때의 Bcl-2 발현은 각각 0.76±
0.05, 1,000μg/ml, 0.92±0.04로 증가하였다. 100μM CoCl2 처치군은 대조군에 비해 Bcl-2 발현이 현저히 감소 하였으나 위령선 추출물 처치에 의하여 그 감소가 유의하 게 억제된 것으로 나타났다(Fig. 5).
Cytochrome c (15 kDa)는 대조군에서의 발현을 1.00 으로 하였을 때, 100μM CoCl2 처치군의 발현이 1.59±
0.16로 나타났다. 위령선 추출물을 100μg/ml, 1,000 μg/ml로 전처치 하였을 때의 cytochrome c 발현은 각각 1.13±0.09, 1,000μg/ml, 1.08±0.09로 감소하였다. 100 μM CoCl2 처치군은 대조군에 비해 cytochrome c 발현 이 현저히 증가하였으나 위령선 추출물 처치에 의하여 그 증가가 유의하게 억제된 것으로 나타났다(Fig. 5).
6. Caspase-3 활성에 미치는 영향
Caspase-3 활성은 DEVD-peptide-nitroanilide (pNA)를 사용하여 측정된다. Caspase-3 활성은 100μM CoCl2 노 출 12시간 후 대조군에 비해 2.78±0.08 pmol에서 7.42±
0.09 pmol로 증가하였다. 반면 100μg/ml과 1,000
μg/ml의 위령선 처치군은 각각 45.66±0.52 pmol과 3.82±0.07 pmol로 각각 감소하였다. 그리고 caspase-3 억제제인 1μg DEVD-FMK를 처치했을 때는 2.81±0.10 pmol로 감소된 것이 확인되었다. 이는 100μM CoCl2 처 치가 caspase-3 활성을 증가시키며, 위령선 추출물 처치 에 의해 caspase-3 활성을 억제시킬 수 있다는 것을 나타 낸다(Fig. 6).
고찰»»»
위령선은 모랑과(미나리아재비과;Ranunculaceae)에 속 한 다년생 낙엽성 만경식물인 으아리속(
Clematis man- shurica
Rupr.)와 위령선(C. chinensis
Osbeck.) 및 棉團 鐵線蓮(C. hexapetala
Pall.)의 뿌리 및 줄기를 건조한 것 이다. 性은 溫하고 無毒하며, 味은 辛鹹하고, 膀胱經에 歸 入하는 약물로, 肌表에 작용하여 祛風除濕하고 通經活絡 하여 止痛하게 하므로 風濕痺痛, 筋脈拘攣, 關節屈伸不利 등의 증후를 치료하는 要藥으로 알려져 있다16). 약리작용 으로는 항염작용, 항균작용, 항뇨작용 및 진통작용에 대 한 연구가 보고되고 있다17).저산소증은 세포나 조직 내 산소 농도가 무산소증과 정상 수준 사이에 있는 대사 상태로, 뇌졸중 및 퇴행성 뇌질환의 신경손상을 관찰하고 그 치료법을 연구하는 과 정에 있어 중요한 기전 중의 하나이다7).
CoCl2는 화학적으로 저산소 상태를 유도하는 물질로, 세포들에 활성산소종의 생성과 더불어 분자생물학적 변 화를 일으키고 DNA 손상과 함께 세포사를 초래한다9). CoCl2의 cobalt 이온은 철이온의 길항제로서 heme내 철 이온을 치환하여 heme protein O2 sensor 내에 구조적 변화를 일으킴으로써 저산소증을 유도한다고 알려져 있 다18).
세포의 저산소 상태가 장시간 지속되거나 그 정도가 매우 극심한 경우, 세포는 사멸하게 된다. 세포사멸이 일 어나는 세포에서는 미토콘드리아로부터 세포질로 cyto- chrome c가 방출되고, 방출된 cytochrome c와 Apaf-1이 caspase에 작용하여 이를 활성화시킨다19). 세포사멸을 방 지하는 단백질인 Bcl-2는 미토콘드리아로부터 cytochrome c의 방출을 억제하여 caspase 활성을 저해시키고20), Bax 는 세포질에서 미토콘드리아로 이동하여 cytochrome c의
유출을 유도한다21). 활성화된 caspase는 직접 또는 간접 적으로 actin 등의 구조단백질에서 세포막의 수포 생성을 유발하고, DNase를 활성화시켜 DNA 분절을 초래하는 등 세포사멸의 특징적인 형태학적 변화를 초래하는 것으로 보고되었다22). 본 연구에서는 CoCl2에 의해 유발된 저산 소성
in vitro
모델에서 위령선 추출물 처치가 인간 신경 아세포종 SH-SY5Y의 세포사멸에 미치는 보호효과에 대 하여 조사하였다.CoCl2에 의한 세포 생존율의 변화를 확인하고 위령선 추출물이 CoCl2 처치에 의해 유도된 세포 생존율에 미치 는 효과가 있는지를 알아보기 위하여 MTT assay를 실시 하였다. 그 결과 CoCl2 농도에 따라 의존적으로 세포 생 존율이 감소되었고, 위령선 추출물 처치에 의해 고농도에 서 세포 생존율 감소가 억제되는 것으로 나타났다.
위령선의 보호효과를 형태학적으로 분석하기 위하여 핵의 세포사멸 특징을 확인할 수 있는 DAPI 염색, DNA 분절을 확인하는 TUNEL 방법, 세포의 사멸을 형태학적으 로 관찰하는 PI 염색방법을 실시하였다. 세포사멸이 일어 나는 세포의 형태적 특징은 우선 핵 내의 염색질이 응집 하며 세포전체가 위축되면서 DNA의 분절이 발생하여 세 포사멸체를 형성한다23). CoCl2 처치는 DAPI로 염색하여 관찰하였는데, CoCl2를 처리한 세포에서 정상세포 핵과는 다르게 핵의 응축이 일어나 세포 조각으로 나눠지려는 초 기 단계의 세포사멸체가 형성되는 것을 관찰할 수 있었으 며, PI 발현 양상도 증가하였다. 반면 위령선 추출물 처치 는 100μM CoCl2 처치에 의해 유도된 핵 조각과 응축된 핵의 형태학적 변화를 억제시켰다. TUNEL 양성세포는 DNA가닥이 조각날 때 free 3'-OH 그룹을 감지하는 것으 로 세포사멸 과정 중에 발생하며, 이러한 DNA 분절 절단 은 TUNEL 분석을 통하여 관찰할 수 있다24). TUNEL 양 성세포수는 100μM CoCl2 처치에서 증가하였으며, 위령 선 추출물을 처치한 세포에서는 증가가 억제되는 양상을 보였다. 이러한 결과는 CoCl2 처치에 의해 세포사멸의 형 태학적 변화가 초래되었다는 것을 의미하며, 위령선 처치 에 의해 세포사멸의 형태학적 특징을 감소시키는 효과가 있다는 것을 의미한다.
DNA 분절화 현상은 세포사멸 현상을 알아보는 중요한 변인으로, 세포사멸의 초기 단계에 보편적이고 필수적 역 할을 한다25). 본 연구에서 100μM CoCl2 처치는 DNA 분 절을 확인 할 수 있었다. 반면, 위령선 추출물은 CoCl2 처
치에 의해 유도된 DNA 분절의 강도를 감소시켰다. 이러 한 결과는 위령선 추출물이 CoCl2에 유도된 DNA 손상과 세포사멸을 감소시킨다는 것을 의미한다.
HIF-1α은 저산소증에 대한 대사과정에서의 전사인자 의 일종으로 저산소증상태에서 특이적으로 발현된다. 정 상적인 상태에서 HIF-1α은 빠른 속도로 분해되나 저산 소 상태에서는 안정화되어 전사인자(transcriptional fac- tor)로 작용하여 세포의 생존에 필요한 여러 가지 물질의 발현을 유도한다26). 반면 지나친 HIF-1α의 발현은 세포 사멸을 유도하는 신호 체계를 활성화 시킨다27). 본 연구 에서 위령선 추출물이 저산소증의 지표인 HIF-1α의 발 현에 미치는 영향을 관찰한 결과, 100μM CoCl2 노출에 의해 HIF-1α 발현이 증가하였으며, 위령선 추출물은 농 도 의존적으로 HIF-1α 발현을 감소시켰다.
Bax와 Bcl-2는 세포사멸과 밀접한 관계를 가지고 있는 단백질로, Bax는 세포사멸의 억제에 관여하는 Bcl-2의 활 성을 감소시키며, Bax의 과발현은 세포사멸로 인한 세포 사를 촉진시킨다28). 본 연구에서 Bax 발현은 100μM CoCl2 처치에 의해 증가하였는데 이러한 Bax의 과발현은 세포사멸의 증가를 의미한다29). 반면 위령선 추출물 처치 는 CoCl2에 의해 유도된 Bax 발현을 감소시켰다. Bcl-2 발현은 100μM CoCl2 처치에 의해 감소하였는데 이러한 Bcl-2의 억제는 세포사멸의 증가를 의미한다30). 그러나 위령선 추출물 처치는 CoCl2에 의해 감소된 Bcl-2 발현을 증가시켰다. 이러한 HIF-1α, Bax 발현의 감소와 Bcl-2 발현의 증가는 위령선 추출물이 항 세포사멸 효과가 있다 는 것을 의미한다.
Cytochrome c는 apoptosis 유도를 활성화 시키는 Bcl-2 family의 신호전달을 통해 활성화 되어 미토콘드리 아에서 유리된다31). 또한 cytochrome c의 유리는 세포사 멸을 유발하는 단백질인 Bax에 의해 유도될 수 있는 것 으로 알려지고 있다32). 본 연구에서는 CoCl2 처치는 cyto- chrome c의 발현량을 증가시켰으며, 위령선 추출물의 처 치는 증가된 cytochrome c의 발현량을 억제시켰다. 따라 서 위령선 추출물은 CoCl2에 의해 유도된 세포사멸에서 Bcl-family의 발현변화를 통하여 cytochrome c의 유출을 억제하는 것으로 생각된다.
Caspase-3는 세포사멸에 필수적인 역할을 하는 단백질 분해 효소로서 DNA 분절화 현상과 핵의 응축을 유도하 면서 세포사멸을 일으킨다33). 저산소증은 caspase-3 발현
에 중요한 인자로, 신경아세포종에서 caspase-3 활성을 증가한다고 하였다34). 본 연구에서 caspase-3 활성은 100 μM CoCl2 처치에 의해 증가하였으며, 반면 위령선 추출 물 처치는 CoCl2에 의해 증가된 caspase-3 활성을 억제시 켰다. 이러한 결과는 CoCl2 처치에 의해 세포사멸의 실행 자 역할을 하는 caspase-3 발현을 증가시키며, 위령선 추 출물이 세포사멸을 억제한다는 것을 의미한다.
본 연구 결과를 종합하여 볼 때, 위령선 추출물이 CoCl2에 의해 유도된 저산소 상태에 의해 발현이 증가되 는 HIF-1α와 Bax 발현의 발현을 억제시켰으며, Bcl-2 발 현을 증가시켰다. 또한 세포사멸의 특징적인 형태학적 변 화에서도 개선효과가 나타났다. 이것으로 미루어 보아, 위령선 추출물이 저산소증의 상태를 개선시키고 저산소 증으로 인해 나타날 수 있는 세포사멸의 감소에 도움을 준다는 것을 알 수 있다.
결론»»»
CoCl2에 의해 유발된 저산소성
in vitro
모델에서 위령 선 추출물 처치가 인간 신경아세포종 SH-SY5Y의 HIF-1α 발현과 세포사멸에 미치는 효과에 대하여 실험하여 다음 과 같은 결과를 얻었다.1. 세포 생존율이 CoCl2 처치로 인해 농도 의존적으로 감소되었으나, 위령선 추출물 처치에 의해 세포 생존율의 감소가 농도 의존적으로 억제되었다.
2. 세포사멸의 특징인 형태학적 변화가 CoCl2 처치로 인해 증가되었으나, 위령선 추출물 처치에 의해 그 변화 가 억제되었다.
3. 세포사멸에서 특징적으로 나타나는 DNA 분절화가 CoCl2 처치에 의해 유발되었지만, 위령선 추출물 처치에 의해 DNA 분절화의 강도가 감소되었다.
4. 저산소증의 지표인 HIF-1α 발현이 CoCl2 처치에 의해 증가하였으나, 위령선 추출물 처치에 의해 농도 의 존적으로 HIF-1α 발현이 억제되었다.
5. Bax, cytochrome c의 발현과 caspase-3의 활성은 CoCl2에 의해 증가하였으나, 위령선 추출물 처치에 의해 그 증가가 억제되었다. 그리고 Bcl-2 발현은 CoCl2 처치 에 의해 감소하였으나, 위령선 추출물 처치에 의해 Bcl-2 발현 증가가 억제되었다.
이상의 결과를 종합하여 볼 때, 위령선 추출물이 저산 소증의 상태를 개선시키고 저산소증으로 인해 나타날 수 있는 세포사멸의 감소에 도움을 준다는 것을 알 수 있다.
참고문헌»»»