*Corresponding author E-mail: [email protected]
서해 고군산군도 서측 해역 표층 퇴적물에서의 규편모류 산출 특성
고영구
전남대학교 지구과학교육과
요 약: 고군산군도 서측해역의 17개 정점의 표층퇴적물에서 4속 16종 2품종의 규편모류가 산출되었다. 이중,
Di. messanensis forma messanensis와 Di. messanensis forma spinosa가 매우 우세한 분류군으로 나타났다.연구해역의 규편모류 골격에서는 어두운 색과 비정형개체들이 다산되며 비정형 개체들은 5 유형으로 분류되었
다. 규편모류 군집조성에 의하면, 연구해역은 온난수괴의 영향을 받는 저염분의 연근해역에 해당하며 해저지형 과 관련된 다양한 조류환경이 조성되고 있는 것으로 생각된다. 추가로, 연구해역의 이런 환경적 조건들은 불규 칙한 기저환을 포함하는 비정형 생성에 주로 영향을 미치는 것으로 여겨진다.
주요어: 어두운 색, 비정형, 연근해, 저염분, 온난수괴
서 론
우리나라 서해의 변산반도 북서해역에 분포하는 고군산군도는 선유도, 무녀도, 방축도, 관리도, 신시도 등 을 비롯한 여러 작은 섬들로 구성된다. 고군산군도 안쪽의 연안 해역은 해저지형 상, 반 폐쇄적(semi-closed) 인 환경을 이루고 있으며 이 군도의 방축도, 선유도, 관리도 등으로 둘러싸인 서측 연안역은 동측 연안역에 비해 비교적 깊은 해저곡(submarine valley)들이 형성되어 있어 다양한 심도변화를 하는 복잡한 퇴적환경을 이루고 있다(이희준 외, 2008; 해양수산부, 2008). 이 연안역의 표층퇴적물은 주로 사질 실트(sandy silt), 실 트질 사(silty sand), 니질 사(muddy sand) 등으로 이루어지며 해역 내의 시료 채취 정점들에 따라 입도 변화 폭이 큰 것으로 알려진다(이희준 외, 2008).
서해를 포함한 우리나라 주변 해역의 퇴적층들에서는 유공충이나 규조를 비롯한 석회질과 규질의 다양한 미생물 유해군집과 미화석들이 풍부히 산출되고 있어 미고생물학적 연구를 통한 해양 퇴적환경 및 층서적 연 구들이 이루어져 왔다(정혜경 외, 1992; 이유대 외, 1996; 김정무 외, 1999; 박영숙 외, 2010; 최정민 외, 2010; 박영숙, 2012). 이들 미생물 유해와 미화석 분류군들 중, 규편모류(silicoflagellate)는 분류군의 조성과 산출비의 특성이 해양환경의 좋은 지시자 역할을 하는 한편, 다른 미생물 유해 분류군들이 드문 한랭한 고위 도 해역 퇴적층의 층서적 연구에 큰 역할을 하고 있다(Shitanaka et al., 1970; Ling, 1973; Poelchau, 1976;
Ciesielski and Weaver, 1974; Henriksen et al., 1993; Barron et al., 2004; Onodera and Takahashi, 2005;
Takahashi et al., 2009). 특히 규편모류는 각 분류군 내에서 전형적이지 못한 비전형적 형(atypical form)이나 기하학적으로 비정상적(abnormal)인 비정형 개체들(aberrant individuals)의 출현이 빈번한 편이다. 이러한 비정형 개체들의 산출특성은 Frenguelli (1935)에 의해 아르헨티나 해역에서 연구된 이래, 주로 환경적 영향 과 관련이 있는 것으로 알려지고 있다(Shitanaka, 1983; Osawa et al., 2005). 또한 규편모류 개체들의 상당수 는 투명한 골격을 보유하지 않고 갈색에서 흑색으로 착색된 골격을 지니는 어두운 색 개체(dark individual) 들의 산출이 자주 나타난다. 특히 매우 어둡게 착색된 개체들은 해저 저탁류 등에 의한 재동(reworking)이나 재퇴적(redeposition)을 시사하는 것으로 보고된다(Poelchau, 1976).
한편, 고군산 군도 동부의 무녀도와 신시도 주변 해역의 표층퇴적물에서의 규편모류 산출특성을 보면, 어
두운 색 개체들과 비정형 개체들의 산출이 빈번하여 육지에 인접하고 영양염류가 풍부한 연안 환경적 특징을
Fig. 1. Map showing sampling stations in the study area (modified after Lee et al., 2008; ◆WG: sampling stations).
반영하는 것으로 시사된 바 있다(고영구 외, 2011). 따라서 이 연구에서는 고군산 군도 서측 연안 해역의 표층 퇴적물에서 산출되는 규편모류 분류군들의 산출상 조사와 아울러, 각 분류군 내의 비정형 개체들의 유형별 빈도 및 어두운 색 개체들의 산출빈도를 살펴 규편모류 각 분류군들의 산출특성과 해양환경과의 관련성을 파 악하기 위한 기초자료를 제공하려 하였다.
연구시료와 방법
연구시료
이 연구에 이용된 퇴적물 시료는 한국해양연구원에서 수행한 고군산군도 해역의 표층퇴적물 시료(이희준 외, 2008; 해양수산부, 2008) 중, 주로 군도 서측의 방축도, 선유도, 관리도 등 도서들의 연안 퇴적물 시료를 사용하였다. 이들 시료에서는 주로 니질이 풍부한 부분을 규편모류 연구에 사용하였다(Fig. 1).
연구방법
규편모류 개체의 추출
표층퇴적물 시료의 규편모류 개체들을 추출하기 위해 주로 규조류 연구에 사용되는 Schrader (1974)의 방 법을 사용하였다. 오븐에서 충분히 건조한 퇴적물 시료를 비커에 넣고 염산(HCl), 과산화수소수(H
2O
2), 증류 수를 혼합한 용액을 가하여 가열판 위에서 70℃ 정도로 반응시켰다. 그 후, 비커에 증류수를 부어 부유물을 완전히 가라앉혀 윗물을 제거하는 것을 2-3회 시행하였다. 탄산염의 제거와 확산이 끝난 시료는 규편모류 개 체들을 집중시키기 위해 실크 스크린으로 체질하였다(Ling and Kim, 1983). 선별된 시료는 스포이드로 흡입 하여 슬라이드 글라스 위에서 잘 분산시켜 건조하여 각 시료 당 2개의 스미어 슬라이드를 제작하였다. 규편 모류 개체들은 광학현미경 하에서 ×100 ~ ×500의 배율로 관찰하고 사진촬영을 하였다.
규편모류 분류군별 빈도 산정
현미경 하에서 규편모류 개체들의 관찰은 개체수 차이에 따른 편차를 최소화하기 위해, Mandra (1968)의
방법을 적용하여 2개의 슬라이드 내에서 200개체들을 관찰해서 각 분류군 별 빈도를 산출하였다. 200개의 규편모류 개체를 관찰한 후에 나타나는 분류군은 ‘tr’로 기입하였다. 규편모류 개체 빈도를 산출할 때는, 골격 이 전체 크기의 2/3 이상 보존되어 분류가 가능한 개체들만을 그 대상으로 하였다. 각 분류군 별 개체들을 산 정할 때는 어두운 색 개체들과 각 분류군에 속하는 비정형 개체들의 빈도를 유형별로 함께 기입하였다.
규편모류의 분류는 연구자에 따라 기저환(basal ring), 정부구조(apical apparatus), 침(spine) 등의 기하학적 배열 특징에 따른 분류군 설정에 다양한 관점의 차이들을 보이고 있다. 이 연구에서는 규편모류의 구분에 Loeblich et al. (1968), Poelchau (1976), Takahashi (1991), Desikachary and Prema (1996), Parkinson (2002) 등의 기준을 고려하였으며, Dictyocha 속과 Distephanus 속은 속명의 축약이 중복되므로 Di. 와 Ds. 로 구분하였다. 연구해역에서 산출되는 다수의 규편모류는 종 수준의 분류를 하였으나 Dictyocha 속의 Di. mes-
sanensis는 기저환의 크기와 형태, 침의 배열, 정부구조의 특성이 분명히 구분되어 품종 수준의 분류를 행하였다(Poelchau, 1976). 그러나 Poelchau (1976)와 Takahashi (1991)의 연구들에서는 기저환의 전개나 정부구조 및 침들의 광범위한 변이 형태들을 Di. messanensis에 속하는 분류군들에 포함시키고 있다. 따라서 이 연구에 서는 Poelchau (1976)와 Takahashi (1991) 등에 의해 Di. messanensis 분류군들에 포함시킨 일부 규편모류 분 류군들을 기저환과 정부구조의 특성에 근거하여 종 수준의 분류를 하였다(Bukry and Foster, 1973).
한편, 전술한 것처럼 연구해역에서 산출되는 규편모류들은 매우 다양한 형태의 비정형 개체들을 포함하고 있다. 이 비정형 개체들은 Takahashi et al. (2009)의 연구에서 제시된 비정형의 유형을 고려하여 5 유형(type) 으로 구분하여 규편모류의 분류군별로 빈도수를 각각 산정하였다.
규편모류 군집
고군산군도 서측 방축도, 선유도 및 관리도 연안 해역의 17개 정점의 표층퇴적물 시료들에서는 4속 16종 2 품종의 규편모류들이 산출되었다. 이들 중, 형태적으로 분명하게 구분되는 Dictyocha messanensis는 Di.
messamensis forma messanensis와 Di. messanensis forma spinosa의 2 품종으로 분류하였다(Table 1).
규편모류의 분류군 별 산출
연구해역에서 산출되는 규편모류 속들의 분포를 보면, Dictyocha 속이 10종 2품종으로 분류군들의 다양도 가 높고 개체빈도도 매우 우세한 반면, Distephanus 속은 4속 4종, 그리고 Corbisema와 Octactis 속들에서 각각 1종이 관찰되어 Dictyocha 속을 제외한 나머지 속들의 산출량은 매우 저조한 편이었다. 다만, Distephanus
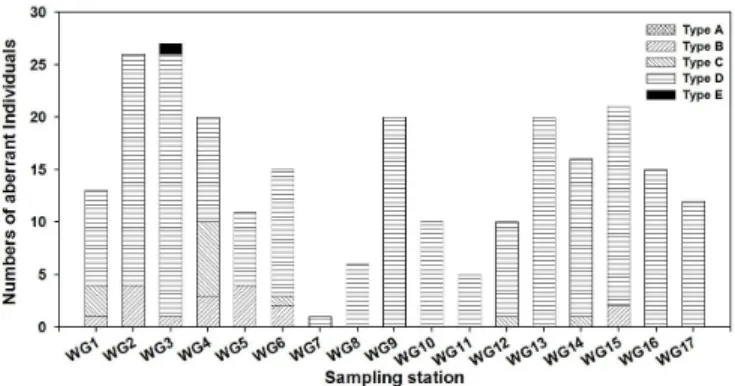
crux와 Octactis pulchra는 각 정점마다 지속적인 산출량을 보이고 O. pulchra는 정점에 따라서는 10여 개체이상이 관찰되기도 하였다. 이들 규편모류 개체들은 연구해역의 전 정점들에서 거의 대부분 60% 이상을 넘 는 어두운 색 개체들과 25% 이상의 비정형 개체들을 함유하는 것으로 나타난다(Table 1; Fig. 2).
연구해역에서 Dictyocha 속의 산출은 Di. messanensis forma messanensis와 Di. messanenesis forma spi-
nosa에 집중되며 그 중에서도 Di. messanenssis forma spinosa가 가장 우점 분류군으로 나타났다. Di. messa- nensis는 마름모꼴 기저환을 지니는 규편모류로, 전형적인 개체인 경우, 정부구조의 정부주가 약간 우측으로경사진다. 이 종의 Di. messanenesis forma messanensis는 소형 기저환을 가지는 반면, Di. messanensis forma
spinosa (Pl. 1, fig. 12)는 보다 큰 기저환, 정부침(apical spine), 기저환에서 안쪽으로 발출하는 지지침(supporting spine)들의 특성에 의해 Di. messanensis forma messanensis (Pl. 1, fig. 10)와 구별된다(Poelchau, 1976). Takahashi (1991)는 이들 두 품종들을 아종(subspecies) 수준의 Di. messanensis messanensis와 Di.
messanensis spinosa로 다루었으나 이 연구에서는 Poelchau (1976)의 제안에 따랐다. 그리고 Di. messanensis
forma spinosa는 기저환의 형태, 지지침의 배열, 정부구조의 형태 등에서 매우 다양한 범위의 변이형들을 보
유하는 것으로 보고된다(Marshall, 1934; Poelchau, 1976; Takahashi, 1991). 그러나 연구해역에서 산출되는
규편모류 개체들 중, 기저환이 5각형이나 6각형인 일부 개체들의 경우는 정부구조를 이루는 정부주와 측방주
(lateral bar)의 연결 방식이 마름모꼴 기저환을 형성하는 Di. messanensis forma spinosa와는 다르게 분명한 5
Station No.
Taxa WG
1 WG
2 WG
3 WG
4 WG
5 WG
6 WG
7 WG
8 WG
9 WG 10 WG
11 WG 12 WG
13 WG 14 WG
15 WG 16 WG
17
Corbisema triacantha tr tr
Dictyocha aculeata(?) 1 2 tr 2
(1) (2) (2)
Dictyocha aspera tr tr 1 tr 1 3 tr tr tr 1
(1) (1) (1)
Dictyocha calida tr tr
Dictyocha deflandrei(?) tr tr tr tr tr
Dictyocha fibula
5 12 7 6 5 6 9 8 2 4 tr 2 tr 1 2 8 5
(5) (12) (7) (4) (4) (5) (6) (8) (2) (4) (2) (1) (2) (7) (1)
*1 *1 *1
Dictyocha medusa 1 tr tr 1
(1) Dictyocha messanensis
forma messanensis
80 89 105 66 54 71 35 41 70 51 76 58 66 72 49 45 34
(72) (73) (84) (57) (48) (48) (31) (33) (55) (41) (60) (48) (56) (62) (42) (37) (30)
*13 *26 *27 *20 *11 *15 *1 *6 *20 *10 *5 *10 *20 *16 *21 *15 *12 Dictyocha
messanenesis forma spinosa
111 89 86 119 138 111 143 144 122 131 124 132 130 115 147 136 146 (95) (75) (71) (99) (117) (76) (120) (119) (96) (99) (82) (107) (108) (99) (125) (119) (92)
*50 *25 *25 *45 *64 *31 *34 *53 *32 *58 *42 *29 *31 *27 *68 *47 *50
Dictyocha pentagona
tr 1 3 tr 2 2 1 tr 2 tr 2 1 tr 1 2 2
(3) (1) (2) (1) (2) (1) (1) (1) (2)
*1 *2 *2 *1 *1 *2
Dictyocha perlaevis tr tr tr tr tr
Dictyocha pseudofibula
1 1 1 1 tr tr tr tr tr
(1) (1)
*1 *1 *1
Distephanus crux
1 1 tr 1 2 2 1 3 tr tr 2 tr 1 tr
(1) (1) (1) (2) (2) (1) (2) (1) (1)
*1 *1 *2 *1
Distephanus octangulatus
1 1 tr
(1) (1)
*1 *1
Distephanus quinquangellus
tr tr 1 tr
(1)
Distephanus speculum 1 tr 1
(1) (1)
Octactis pulchra 1 6 2 tr 2 9 9 5 7 tr 3 2 10 1 8 12
(5) (2) (6) (7) (4) (6) (2) (2) (10) (1) (7) (12)
Sum
200 200 200 200 200 200 200 200 200 200 200 200 200 200 200 200 200 (175) (167) (164) (169) (169) (137) (168) (165) (158) (155) (142) (162) (168) (173) (171) (172) (137)
*66 *53 *52 *68 *76 *50 *37 *63 *52 *68 *47 *39 *51 *43 *90 *65 *62
Table 1. Occurrences of silicoflagellates in the study area (Note: tr - silicoflagellate taxa observed after counting, ( )-dark individuals), *-aberrant individuals).
방향이나 6방향의 기하학적 분지형태(geometrical branched type)를 이루어서 이들은 Di. pentagona (Pl. 1, fig. 20)와 Di. pseudofibula (Pl. 1, fig. 23)로 구분하였다(Tsumura, 1963; Bukry and Foster, 1973).
연구해역에서 Di. messanensis forma messanensis와 Di. messanensis forma spinosa의 산출빈도는 산출
규편모류 개체들의 90%를 넘을 정도로 지배적이며 Di. messanensis forma spinosa의 산출이 Di. messa-
nensis forma messanensis보다 많은 편이다. Di. messanensis forma messanensis의 빈도는 연구해역의 방축도 북측 연안 해역에서는 Di. messanensis forma spinosa의 산출과 다소 유사한 개체수를 보이나 도서들로
둘러싸인 나머지 연안역들에서는
Di. messanensis forma spinosa의 산출빈도가 훨씬 증가한다. 이와 함께 도서들로 둘러싸인 이 해역에서는 Di. messanensis forma spinosa의 전체 개체수와 함께 이 분류군의 어두운
색 개체들의 빈도도 같이 증가한다. 이들 두 품종들은 형태적인 면에서도 다양한 비정형 개체들을 포함한다.
Fig. 2. Total, dark and aberrant individuals of silicoflagellates in the study area (WG: sampling stations).
특히, Di. messanensis forma spinosa는 비정형 개체들의 빈도가 Di. messanensis forma messanensis에 비해 많을 뿐만 아니라 그 변이도 광범위하게 나타난다.
산출되는
Dictyocha류 중에서 마름모꼴이나 다소 둥근 사각형 기저환을 보유하고 기저환의 종축(longitudinal axis)에 거의 나란한 정부주를 가지며 정부침이 없는 개체들은 Di. fibula (Pl. 1, fig. 7)로 분류 하였다. Di. fibula는 Dictyocha 속의 대표적인 종으로 매우 다양한 변이를 가지는 것으로 알려지나, 이 분류 군의 형태적 특성을 보다 명확히 하기 위해 앞에 기술한 골격특성을 가지는 규편모류로 다시 제안되고 있다 (Bukry and Foster, 1973). 이 연구에서도 Di. messanensis forma spinosa의 변이 또는 비정형 개체들과 구별 하기 위해 이러한 형태적 특성에 따른 분류를 하였다. 그러나 Di. messanensis forma spinosa의 비정형 개체 들 중, 기저환의 전개가 불완전한 비정형 개체는 Di. fibula와의 구별이 어려운 경우가 많은 편이다. 이 경우에 는 정부침의 부재와 함께 정부주와 연결주의 연결부가 단순하면서 둔각으로 연결되는 경우는 Di. fibula (Pl.
1, fig. 8)로 분류하였다. 연구해역에서 Di. fibula는 고군산 군도 북측의 외해 쪽 연안역에서는 정점 별로 200 개체 당, 5개체 이상의 산출을 하나 방축도와 관리도 사이의 도서들로 둘러싸인 일종의 내해역(inshore area) 에서는 감소하는 경향을 가진다. 이 분류군의 어두운 색 개체들은 산출 개체들의 거의 대부분을 차지하나 상 대적으로 비정형 개체들의 함량은 저조한 편이다.
연구해역에서 Dictyocha 속의 나머지 종들의 산출은 극히 저조하다. 전체적으로 사각형의 기저환에 작은 사각형이 사이에 엇갈리게 교차한 것 같은 준팔각형(suboctagon) 기저환을 지니는 Di. aculeata는 연구해역 에서 극소수가 거의 어두운 색 개체들로 산출된다. 이 종은 기저환의 외관이 다소 불규칙하나 준팔각형을 이 루는 점에서 Di. aculeata로 분류하였다. 작은 정부판(apical strut)과 연결주가 기저환 외부로 다소 연장되면 서 기저침으로 연결되는 골격특성을 지니는 규편모류는 그 특성상 Di. deflandrei (Pl. 1, fig. 6)로 생각된다.
이 종은 연구해역에서 극소수의 어두운 색 개체들만이 관찰될 뿐이다.
정부주가 전개되지 않고 십자형으로 연결주가 교차하는 Di. medusa (Pl. 1, fig. 9), 기저환과 연결주가 접 합되는 부위가 안쪽으로 약간 함입되는 Di. perlaevis (Pl. 1, fig. 22) 및 오각형 기저환을 지니는 Di. pentago-
na (Pl. 1, fig. 20) 및 종축방향으로 신장되는 육각형 기저환을 가진 Di. pseudofubula (Pl. 1, fig. 23) 등의 규편모류 개체들 역시 개체빈도가 매우 저조하다. 이들 분류군의 개체들 역시 비정형 개체나 변이형에 해당되 는 경우가 다수를 이룬다. 또한 연결주와 기저환의 접합부가 함입되고 종축 방향으로 신장된 기저환이 명확 한 좌우대칭을 이루는 Di. calida (Pl. 1, fig. 5)와 다른 분류군들과는 달리 정부주의 전개가 기저환의 종축방 향과 직각인 Di. aspera (Pl. 1, fig. 4)도 소수만이 출현할 뿐이다. 이들 두 분류군의 기저환의 크기는 다소 작 은 편이나 골격의 전개형태에서 전기한 분류군들에 해당시켰다.
연구해역의 Distephanus 속의 규편모류 개체들은 Ds. crux가 소수의 개체가 산출되지만 대부분의 정점에
서 확인되는데 반해 다른 분류군들은 극소수 개체들이 국한된 정점들에서만 관찰될 뿐이다. 이 속은 정부구
조에 다각형 또는 둥근 모양의 정부창(apical window)의 존재로 특징지어지는데, 창의 크기, 위치 및 기하학
적 규칙성 등을 바탕으로 Di. messanenesis forma spinosa, Di. pentagona 및 Di. pseudofibula 등의 비정형 개체들과 구분하였다. Ds. crux (Pl. 1, fig. 25)는 거의 대다수가 어두운 색 개체로 정부창이 둥근 사각형이고 정부창에 짧은 지지침이 돌출되는 비정형 개체들의 비가 높은 편이다. 이 외에 오각형 기저환을 가지는 Ds.
quinquangellus, 육각형 기저환을 보유하는 Ds. speculum, 정팔각형 기저환과 짧은 기저침을 가지는 Ds. oc- tangulatus 등은 극소수의 개체들만이 확인될 뿐이다. 그리고 Ds. octangulatus의 산출이 극소수이기는 하나
방축도 북측의 외해역에 국한되는 것도 특징적이다.
삼각형 기저환이 특징적인 Corbisema 속은 기저환의 전개 상태로 미루어 C. triacantha 로 생각되는 수개 체가 확인될 뿐이다(Desikachary and Prema, 1996). 다만 이 연구에서, C. triacantha의 삼각형 기저환 유형과
Di. messanensis forma spinosa 비정형 개체의 corbisemoid 유형은 기저침과 기저환의 전개 상태로 구분하였다. 특징적인 팔각형(octagon) 기저환과 매우 취약하고 가늘어 파손되기 쉬운 정부환을 보유하는 Octactis
pulchra는 기저환의 형태가 팔각형, 구각형(nonagon), 십각형(decagon)을 취할 수 있으며, 연구해역 일대에서는 구각형 기저환이 상대적으로 우세하고 규칙적인 형태를 보유하여 종의 특성에 대한 재검토가 요청됨을 지적한 바 있다(고영구 외, 2011). O. pulchra는 비교적 소수이긴 하지만 거의 대부분의 정점에서 지속적인 산출을 보이며, 구각형 기저환을 보유하는 개체들이 우세하고 대다수가 어두운 색 개체들로 이루어진다.
어두운 색 개체들의 산출
연구해역에서 어두운 색 개체들의 산출은 관찰된 전 규편모류 개체들의 60% 이상을 능가한다(Fig. 2). 특 히 Di. messanensis forma spinosa의 경우, 어두운 색 개체들의 빈도가 방축도와 관리도 내측의 내해역에서 더욱 증가한다(Tables 1; Fig. 1). 연구해역에서 어두운 색 개체들의 빈도는 매우 높지만 그 착색 정도와 착색 되는 골격의 부위에서 매우 다양한 범위에 걸친다. 먼저 착색 정도에서 골격이 거의 불투명한 정도(Pl. 1, fig.
8)에서 약간 어두운 정도(Pl. 1, fig. 17)에 이르기까지 넓은 착색농도 범위를 보인다. 이와 함께 골격의 착색 이 부위에 따라 선택적으로 진행되는 경우도 빈번히 관찰된다. 한 예로 기저환은 착색 정도가 강한데 비해 기 저침과 정부구조는 투명한 골격에 가까운 상태로 나타난다(Pl. 1, fig. 30). 일반적으로 골격이 거의 불투명할 정도로 강하게 착색된 개체들은 주로 대륙사면 등에서 저탁류나 등고선류 등에 의한 퇴적물의 재동 (reworking)이나 재퇴적(redeposition)에 기인하는 것으로 보고된다(Poelchau, 1976).
그러나 연구해역에서 어두운 색 규편모류 개체들의 산출은 착색된 정도와 착색 부위에서 매우 다양한 범 위를 지닌다. 그리고 분류군과 어두운 색을 띠는 개체빈도 간의 분명한 관계를 찾기는 어려웠으나 규편모류 골격의 특성 또는 환경적 요인에 따른 차별적인 착색현상이 관련될 가능성도 제기된다.
비정형 개체들의 산출
일반적으로 규편모류 개체들에서 비정형 개체들은 기하학적 형태에서 벗어난 비전형적(atypical) 또는 불 규칙한(irregular) 형태를 가진 개체들로 알려진다. 이러한 비정형 개체들에는 대체로 골격의 간극(gap)을 가 진 불완전한 기저환, 기저환의 각 우각(corner)에서 확장되는 기저침들의 비정상적인 크기, 기저환의 부재 및 정부환의 불균일한 크기나 비기하학적 배열 등을 보이는 개체들이 속한다(Takahashi et al., 2009),
이 연구에서는 연구해역 규편모류들의 산출과 Takahashi et al. (2009)의 제안을 고려해 5가지의 비정형 유 형을 설정하였다. 먼저 골격의 간극을 포함하여 불완전한 기저환을 지니는 유형 A (type A; Pl. 1, fig. 13), 둘 째로 지지침들의 배열이 비정상적인 유형 B (type B; Pl. 1, fig. 14), 셋째로 비정상적인 정부구조를 가지는 유형 C (type C; Pl. 1, fig. 20), 폐쇄된 형태이나 불규칙하게 전개되는 기저환을 가지는 유형 D (type D; Pl.
1, fig. 11) 그리고 기저환이 전혀 없거나 매우 불규칙한 골격 등 다른 비정형 특성을 나타내는 유형 E (type E;
Pl. 1, fig. 19) 등으로 구분하였다(Table 2). 이 구분에서 정부창의 비정상적인 배열과 정부주의 불규칙성은 유형 C에, 기저침들의 비정상적인 형태는 유형 D에 포함시켰다.
연구해역의 규편모류들에서 나타나는 비정형 개체들은 주로 개체빈도가 높은
Di. messanensis forma messanensis와 Di. messanensis forma spinosa에 집중적으로 출현하며 일부는 Di. pentagona, Di., pseudo-Station
Taxa AI WG1 WG
2 WG
3 WG
4 WG
5 WG
6 WG
7 WG
8 WG
9 WG
10 WG 11 WG
12 WG
13 WG
14 WG
15 WG
16 WG
17 A
Dictyocha messanensis forma messanensis
B 1 4 1 3 4 2 2
C 3 7 1 1 1
D 9 22 25 10 7 12 1 6 20 10 5 9 20 15 19 15 12
E 1
Sum of aberrant
individuals 13 26 27 20 11 15 1 6 20 10 5 10 20 16 21 15 12
Total
individuals 80 89 105 66 54 71 35 41 70 51 76 58 66 72 49 45 34
% of aberrant
individuals 16.3 29.2 25.7 30.3 20.4 21.1 2.9 14.6 28.6 19.6 6.6 17.2 30.3 22.2 42.9 33.3 35.3
A 3 1 1 1 5 1 2
Dictyocha messanenesis forma spinosa
B 29 16 18 28 54 12 30 40 27 51 35 25 27 23 60 41 47
C 7 2 1 4 2 9 1 2 2 3 2 4 3 4 1
D 11 6 5 13 8 4 1 6 2 5 4 2 1 3 2 3
E 1 5 2 1 1 1
Sum of aberrant
individuals 50 25 25 45 64 31 34 53 32 58 42 29 31 27 68 47 50
Total
individuals 111 89 86 119 138 111 143 144 122 131 124 132 130 115 147 136 146
% of aberrant
individuals 45.0 28.1 29.1 37.8 46.4 27.9 23.8 36.8 26.2 44.3 33.9 22.0 23.8 23.5 46.3 34.6 34.2
Table 2. Occurrences of aberrant individuals for Dictyocha messanensis forma messanensis and Di. messanensis forma
spinosa in the study area (A: aberrant type A with open basal ring, B: aberrant type B with abnormal supporting spines,C: aberrant type C with irregular apical apparatus, D: aberrant type D with irregular basal ring, E: Other irregular aberrant type E except above ones).
fibula, Ds. crux 등에서도 확인된다. 상대적으로 Di. fibula의 비정형 개체들의 빈도는 저조하며 예외적으로 O. pulchra에서는 비정형 개체들이 관찰되지 않았다(Tables 1, 2). 특히 Di. messanensis forma spinosa의 경
우 비정형 개체들의 빈도가 높고 그 유형도 다양한 것은 높은 산출빈도와 골격구조의 다양성이 관련된 것으 로 해석된다. 규편모류 비정형 개체들의 유형별 산출상을 비정형 개체들의 비가 높은 Di. messanensis forma
messanensis와 Di. messanensis forma spinosa들을 중심으로 살펴보면, Di. messanensis forma messanensis는 주로 유형 D에 집중되고 일부는 유형 B와 C에 해당되었다(Fig. 3). 이는 Di. messanensis forma messa-
nensis의 비정형 개체들(Pl. 1, fig. 11)이 주로 기저환의 비정상적인 형태에 집중되고 일부는 지지침과 정부구조의 불규칙한 배열이나 모습으로 출현한다는 것을 지시한다. 한편, Di. messanensis forma spinosa (Pl. 1, figs. 2, 14-17)는 유형 A에서 E에 이르는 비정형 개체들이 다 출현하나 주로 지지침의 불규칙성을 나타내는 유형 B에 집중되며 기저환의 비정상적인 형태인 유형 D가 다음으로 많고 정부구조의 불규칙한 배열 형태인 유형 C도 각 정점마다 지속적으로 출현한다(Fig. 4). 특히 삼각형의 corbisemoid형 개체의 경우는 4개의 기 저침과 기저환의 전개 형태에 의해 Di. messanensis forma spinosa의 비정형 개체로 분류하였으나 Corbisema 의 비정형 개체와의 구분은 기저침과 기저환의 기하학적 형태에 대해 보다 세부적인 검토를 필요로 한다.
Di. messanensis 분류군들을 제외한 다른 규편모류 분류군들의 경우에는 산출 개체수가 적어 비정형 개체
들의 유형별 산출을 판단하기는 어려웠지만 대체로 기저환과 정부구조의 불규칙한 모습이 많은 경향을 보인 다. 특히 Di. pentagona는 저조한 산출빈도에도 기저환과 정부구조의 비정형 개체들이 많이 출현하여 정부구 조에 미세한 정부창을 보유하는 개체들(Pl. 1, figs. 20, 21)이 관찰되기도 한다.
그리고 유형 E에서와 같이 정부구조만 보유하는 경우(Pl. 1, fig. 19), 그 골격이 뾰족하게 첨멸되는 경우
(Pl. 1, fig. 14)나 일부 지지침들이나 연결주들이 크게 전개되어 일종의 집적(accumulation) 상태를 보이는
경우(Pl. 1, fig. 17)는 골격형성 단계와 어떤 관련성을 시사할 가능성도 있다.
Fig. 4. Aberrant types of Dictyocha messanensis forma spinosa (Type A: aberrant type with open basal ring, Type B:
aberrant type with abnormal supporting spines, Type C: aberrant type with irregular apical apparatus, Type D: aberrant type with irregular basal ring, Type E: Other irregular aberrant type except above ones, WG: sampling stations).
Fig. 3. Aberrant types of Dictyocha messanensis forma messanensis (Type A: aberrant type with open basal ring, Type B: aberrant type with abnormal supporting spines, Type C: aberrant type with irregular apical apparatus, Type D: aberrant type with irregular basal ring, Type E: Other irregular aberrant type except above ones, WG: sampling stations).
토의 및 결론
고군산 군도 서측의 방축도, 관리도 및 선유도 등 도서들의 연안해역의 표층퇴적물에서 산출되는 규편모류는
Dictyocha 속에 주로 편중되고 그 중에서도 Di. messanensis forma messanensis와 Di. messaensis forma spino- sa의 두 분류군에 집중된다. 그리고 Dictyocha 속의 다른 종들이나 Distephanus를 비롯한 다른 속들의 산출빈도는 매우 낮다 . 종의 분산도도 Dictyocha 속이 상대적으로 다양하며 다른 속들은 제한적인 산출을 보일뿐이다. 또 한 연구해역에서 산출되는 규편모류 개체들의 60% 이상이 어두운 색 개체들이며 그 착색 정도도 다양하게 전개 되는 한편, 각 분류군 별로 골격의 비정형 유형에 차이가 있으나 비정형 개체들의 구성비가 높게 나타났다.
규편모류의 대표적인 두 속인 Dictyocha와 Distephanus 간의 산출비나 각 분류군들의 산출특성들은 해수 의 수온을 지시하는 좋은 도구로써 알려져 왔다(Gemeinhardt, 1934; Mandra, 1969; Ciesielski and Weaver, 1974; Bukry, 1982). 이러한 연구결과들에 의하면, 연구해역의 규편모류 군집조성에서 Dictyocha 속의 산출 비가 절대적으로 우세하고 한랭수괴(cold water mass)를 시사하는 대표적 분류군인 Ds. speculum과 Ds. oc-
tangulatus가 극히 제한적인 산출을 보이는 것은 연구해역이 온난수괴(warm water mass)의 강한 영향권 아래에 있음을 알려준다.
연구해역의 규편모류 군집에서 가장 우세한 분류군인 Di. messanensis forma messanensis와 Di. messa-
nensis forma spinosa는 세계의 해양에 널리 분포하며 북태평양 서부 해역에서 풍부한 산출이 보고된다. 특히 Di. messanensis forma messanensis는 북위 40° 이하의 저위도 해역에서 매우 빈번히 산출되는 분류군이며Di. messanensis forma spinosa는 Di. messanensis forma messanensis와 수반하여 북태평양의 서부에서 다
산되는 것으로 보고된다(Poelchau, 1976). 그러나 연구해역에서 쿠로시오(Kuroshio) 해류의 영향을 시사하 는 Di. aculeata 개체들이 극히 제한적인 산출을 하는 것은, 연구해역이 온난수괴(warm water mass)의 강한 영향을 받기는 하나 쿠로시오 해류가 직접적인 영향을 미치지는 않는 것으로 판단할 수 있다.
이와 함께, 규편모류의 분류군 별 산출비와 비정형 개체를 비롯한 골격 특성들은 해양환경의 중요한 지시 자가 되며, 특히 해수 중에서의 실리카의 공급, 염도, 영양염류 조건, 태양광 조건 등과 밀접하게 관련된다는 사실들이 알려지고 있다(Mandra, 1969; Lipps, 1970; Shitanaka, 1983; Henriksen et al., 1993; Onodera and Takahashi, 2005; Takahashi et al., 2009). 연구해역의 규편모류 개체들은 산출되는 분류군에 따라 차이를 보 이기는 하지만 비정형 개체들이 풍부하게 포함되며 일부 개체들에서는 분류군 간의 전이형으로 생각될 수 있 는 개체들이 출현한다. 이러한 경향은 Di. messanensis forma messanensis와 Di. messanensis forma spinosa 에서 잘 나타나며 특히 Di. messanensis forma spinosa에서 두드러지게 나타난다. 이러한 골격 특성들은 일 반적으로 연근해(neritic sea)에서 잘 나타나며 특히 육지에 근접한 해역의 저염도(low salinity)와 풍부한 영 양염류(nutrients)가 원인이 되는 것으로 보고된다(Mandra, 1969; van Valkenburg and Norris, 1970; Bukry and Foster, 1973; Shitanaka, 1983; Henriksen et al., 1993; Onodera and Takahashi, 2005). 따라서 연구해역 에서 산출되는 규편모류 개체들에서 비정형 개체들의 높은 빈도는 연구해역이 해안에 인접한 저염분 환경으 로 영양염류가 풍부하게 공급되는 환경임을 지시한다. 이러한 결과는 연구해역이 금강을 비롯한 하천들로부 터 영양염류를 풍부하게 공급받고 있으며 담수의 유입으로 인하여 저염분 환경이 조성되고 있다는 연구결과 를 지지하는 것으로 볼 수 있다(임동일 외, 2008). 이와 함께 규편모류 비정형 개체들의 유형별 산출빈도를
Di. messanensis를 중심으로 살펴보면, 지지침이 비정상적 배열을 하는 유형 B와 기저환이 불규칙한 유형 D의 빈도가 높은 편이나 다수의 규편모류 분류군에서는 유형 D가 비정형의 주류를 이루는 것을 알 수 있다. 이 러한 규편모류 비정형 개체들의 산출특성은 영양염류가 풍부한 저염분 환경에서는 규편모류 골격의 기저환 형성이 가장 많이 영향을 받을 가능성이 있는 것으로 생각된다.
한편, 연구해역의 규편모류 개체들에서는 어두운 색 개체들의 빈도가 매우 높게 나타나고 있다. 전술한 것 처럼 강하게 착색된 어두운 색 개체들의 산출은 저탁류나 등고선류에 의한 퇴적물의 재동이나 재배치에 기인 한 것으로 해석되고 있다(Poelchau, 1976). 그러나 연구해역에서 어두운 색 개체들의 산출은 착색의 정도가 다양하고 골격 부위에 따른 차별적 착색까지 출현하고 있을 정도로 매우 광범위하게 나타난다. 이러한 어두 운 색 개체들의 광범위한 다양성은 저탁류 등의 퇴적물 흐름과는 다른 여러 형태의 재배치작용의 영향을 받 은 결과로 생각된다. 연구해역은 도서들 사이의 해저곡이 전개되는 등 복잡한 해저지형에 따른 다양한 조류 가 이루어지는 해역으로 알려진다(김창수 외, 2006). 또한 연구해역에서 방축도, 관리도 및 선유도 사이의 내 해역에서는 Di. messanensis forma messanensis보다 기저환이 더 큰 Di. messanensis forma spinosa의 산출 이 많고 어두운 색 개체도 Di. messanensis forma spinosa의 빈도가 크게 증가한다. 이러한 크기에 따른 산출 상 차이는 해저지형에 따른 조류 등의 영향으로 인한 규편모류 개체에 대한 일종의 분급(sorting)으로 해석할 수 있다(Poelchau, 1976). 따라서 연구해역에서 어두운 색 개체들의 광범위한 산출과 개체들의 크기에 따른 집중 현상 등은 이 해역이 복잡한 해저지형과 이에 관련된 다양한 조류의 작용을 받고 있는 것으로 생각된다.
위의 연구 결과들을 종합하면, 연구해역은 육지에 근접한 해역으로 담수의 영향을 받는 온난수괴가 지배 적인 해역이며, 해저지형과 관계된 다양한 조류운동 등이 이루어지는 퇴적환경으로 생각된다. 그리고 이러한 환경적 요인들은 규편모류 비정형 개체들의 기저환 생성에 비교적 많은 영향을 미치는 것으로 추정된다.
감사의 글
이 연구에 사용된 고군산군도 해역의 표층퇴적물 시료를 제공해 주신 한국해양연구원에 감사드립니다. 특히
시료와 함께 연구에 여러 가지로 도움을 주신 한국해양연구원의 이희준 박사님께 깊이 감사드립니다. 그리고 논
문의 상세한 부분까지 검토해주시고 좋은 제안을 해주신 박영숙 박사님과 변현숙 박사님께도 깊이 감사드립니다.
CHARACTERISTICS OF SILICOFLAGELLATE
OCCURRENCES FROM THE SURFACE SEDIMENTS OF THE WESTERN AREA IN GOGUNSAN ISLANDS, WEST SEA
Yeong Koo Koh
Department of Earth Science Education, Chonnam National University, Gwangju 500-757, Korea
Abstract: From the 17 surface sediments in the western area of Gogunsan Islands, 16 silicoflagellate species and 2 formas belonging to 4 genera were identified. Of them, Di. messanensis forma messanensis and Di. messanensis forma spinosa were extremely dominate taxa. Among silicoflagellate skeletons in the study area, dark and aber- rant ones are frequent. Aberrant individuals of the silicoflagellates are classified into 5 types. On the basis of sili- coflagellate assemblages from the sediments, it is considered that the study area is assigned to neritic and low sal- ine environment influenced by warm water mass, and variable tidal conditions related with submarine topography are formed in the area. In addition, above environmental conditions might mainly control the aberrant forms in- cluding irregular basal rings of silicoflagellates in the study area.
Key words: dark, aberrant, neritic, low saline, warm water mass
참고문헌고영구, 정철환, 오강호. 2011. 서해 고군산군도 동부 연안 해역의 표층퇴적물에서 산출되는 규편모류, 내골격 와편모조류 및 해면골침. 고생물학회지 27:51-67.
김정무, 박병권, 김대철, 김길영. 1999. 유공충과 안정동위원소에 의한 지난 24,000년 동안의 동해 울릉분지 고해양 변화.
고생물학회지 15:1-20.
김창수, 이상호, 손영태, 권효근, 이광희, 김영배, 정우진. 2006. 새만금 4호 방조제 완성 전 ․ 후 HF 레이다로 관측된 표층 M
2조류의 변화. 한국해양학회지 “바다” 11:37-48.
박영숙. 2012. 서해 금강 연안 홀로세 퇴적물에서 규조류 군집과 해수준 변화. 대한지질학회지 48:113-120.
박영숙, 이성주, 천종화, 이종덕. 2010. 울릉분지의 제4기 퇴적물에서 규조를 이용한 고기후 연구. 고생물학회지 26:183-192.
이유대, 유환수, 김석윤. 1996. 동해 울릉분지의 생물층서와 퇴적상. 대한지질학회지 32:73-90.
이희준, 김민지, 김태경. 2008. 고군산군도 내측해역의 현생퇴적환경. Ocean and Polar Research 30:519-536.
임동일, 강미란, 장풍국, 김소영, 정회수, 강양순, 강영실. 2008. 한국 서해 중부 연안역의 수질환경 특성. Ocean and Polar Research 30:379-399.
정혜경, 백광호, 박병권. 1992, 한국 서남해에서 산출된 현생저서 유공충의 동물군 분석 및 해양환경 연구. 한국해양학회지 27:123-136.
최정민, 강소라, 박일흠, 이연규. 2010. 천수만의 표층퇴적물 및 저서 유공충의 분포특성과 퇴적환경. 고생물학회지 26:107-128.
해양수산부. 2008. 새만금 해양환경보전대책을 위한 조사연구(2단계 2차년도): 방조제 외해역 해저환경 이용. 한국해양 연구원 연구보고서, 425 p.
Barron, J. A., Bukry D. and Bischoff, J. L. 2004, High resolution paleoceanography of the Guayamas Basin, Gulf of California, during the past 15000 years. Marine Micropaleontology 50:185-207.
Bukry, D. 1982. Cenozoic silicoflagellates from offshore Guatemala, Deep Sea Drilling Project Site 495. Initial Reports of Deep Sea Drilling Project 67:425-445, U. S. Government Printing Office, Washington.
Bukry, D. and Foster, J. H. 1973. Silicoflagellates and diatom stratigraphy, Leg 16. Initial Reports of Deep Sea Drilling Project 16:815-871, U. S. Government Printing Office, Washington.
Ciesielski, P. F. and Weaver, F. M. 1974. Early Pliocene temperature changes in the Atlantic seas. Geology
41:159-172.
Desikachary, T. V. and Prema, P. 1996. Silicoflagellates (Dictyochophyceae). Bibliotheca Phycologica 100, Gebrüder Borntraeger, 298 p.
Frenguelli, J. 1935. Variaciones de Dictyocha fibula en el Golfo de San Matías (Patagonia septentrional). Anales del Museo Argentino de Ciencias Naturales "Bernadino Rivadavia" 38, Protistologia 4:265-281.
Gemeinhardt, K. 1934. Die Silicoflagellaten des südatlantischen Ozeans. Wissenschaftliche Ergebnisse der deutschen atlantischen Expedition auf dem Forschungs- und Vermessungsschiff "Meteor" 1925-1927, Band 12:274-312.
Henriksen, P., Knipschildt, F., Moestrup, O. and Thompson, H. A. 1993. Autoecology, life history and toxicology of the silicoflagellate Dictyocha speculum (Silicoflagellata, Dictyochophyceae). Phycologia 32:29-39.
Lee, H. J., Kim, M. J. and Kim, T. K., 2008. Modern Sedimentary Environments within the Gogunsan Archipelago.
Ocean and Polar Research 30(4):519-536 (in Korean with English abstract).
Ling, H. Y. 1973. Silicoflagellates and ebridians from Leg 19. Initial Reports of Deep Sea Drilling Project 19:751-775, U. S. Government Printing Office, Washington.
Ling, H. Y. and Kim, B. K. 1983. Miocene archeomonads from Pohang Area, Korea. Geological Society of Korea 19:247-251.
Lipps, J. H. 1970. Ecology and evolution of silicoflagellates. First North American Paleontology Convention, Proceedings G:965-993.
Loeblich III, A. R., Loeblich, R. A., Tappan, H. and Loeblich, A. R. 1968. Annotated index of fossil and recent silicoflagellates and ebridians with descriptions and illustrations of validly proposed taxa. The Geological Society of America, Inc., 319 p.
Mandra, Y, T. 1968. Silicoflagellates from the Cretaceous, Eocene and Miocene of California, U. S. A. Proceedings California Academy of Science 36:231-277.
Mandra, Y. T. 1969. Silicoflagellates: a new tool for the study of Antarctic Tertiary climates. Antarctic Journal of U. S. 4:172-174.
Marshall, S. M. 1934. The silicoflagellates and Tintinnoinea. British Museum, Great Barrier Reef Expedition 1928-29. Sciences Reports 4:623-662.
Onodera, J. and Takahashi, K. 2005. Silicoflagellate fluxes in the western subarctic Pacific, 1997-2000. Deep Sea Research I 52:371-388.
Osawa, M., Takahashi, K. and Hay, B. J. 2005. Shell-bearing plankton fluxes in the central Black Sea, 1989-1991.
Deep-Sea Research I 52:1677-1698.
Parkinson, P. 2002. Ontogeny v. Phylogeny: the Strange Case of the Silicoflagellates. Constancea 83, 2002:
http://ucjeps. berkeley.edu/constancea/83/parkinson/Dictyocha. html.
Poelchau, H. S. 1976. Distribution of Holocene silicoflagellates in north Pacific sediments. Micropaleontology 22:164-193.
Schrader, H. J. 1974. Proposal for a standardized method of cleaning diatom-bearing deep-sea and land-ex- posed marine sediments. Nova Hedwigia, Beihefte 45:403-409.
Shitanaka, M. 1983. Silicoflagellate remains in the sediments of Lake Hiruga, Fukui, Japan. Mizunami Fossil Museum Research Report 10:171-180.
Shitanaka, M., Ogawa, F. and Ichikawa, W. 1970. Silicoflagellate remains in the deep-sea sediments from the Sea of Japan. Nihonkai (Sea of Japan) 4:1-14.
Takahashi, K. 1991. Silicoflagellates and Actiniscus: vertical fluxes at Pacific and Atlantic sediment trap stations; p.
35 in Honjo, S. (ed.), Ocean Biocoenosis Series No. 2. Woods Hole Oceanographic Institute.
Takahashi, K., Onodera, J. and Katsuki, K. 2009. Significant populations of seven-sided Distephanus(Silicoflagellata) in the sea-ice covered environment of the central Arctic Ocean, summer 2004. Micropaleontology 55(2-3): 313-325.
Tsumura, K. 1963. A systematic study of silicoflagellateae. Journal of the Yokohama Municipal University, Series C-45, 146:1-84.
van Valkenburg, S. D., Norris, E. 1970. The growth and morphology of the silicoflagellates Dictyocha fibula Ehrenberg
in culture. Journal of Phycology 6:48-54.
Plate 1
Explanation of Plate 1