Inhibition of Inflammation by Popillia flavosellata Ethanol Extract in LPS- induced RAW264.7 Macrophages
Young-Il Yoon1, Jae-Sam Hwang1, Mi-Ae Kim1, Mi Young Ahn1, Young-Bo Lee1, Myung Sae Han2, Tae-Won Goo3* and Eun-Young Yun1*
1Department of Agricultural Biology, National Academy of Agricultural Science, RDA, Wanju-gun 55365, Korea
2Department of Bio-fibers and materials Science, Kyungpook National University, Daegu 41566, Korea
3Department of Biochemistry, School of Medicine, Dongguk University, Gyeongju 38066, Korea Received July 29, 2015 /Revised August 11, 2015 /Accepted August 11, 2015
The beetle Popillia flavosellata has been no reported its functional effects. In this study, we investigated the anti-inflammatory effect of P. flavosellata ethanol extract (PFE) on RAW 264.7 mouse macrophage cells treated with lipopolysaccharide (LPS) for the induction of inflammation. First, we examined the cytotoxicity of PFE in the RAW 264.7 cells at a concentration of 2,000 μg/ml or less. To evaluate the anti-inflammatory effects of PFE, we investigated the expression levels of proinflammatory cytokines, such as tumor necrosis factor (TNF)-α and interleukin (IL)-6, and proinflammatory enzymes, such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) in LPS-induced RAW 264.7 cells.
In addition, we examined whether PFE inhibited the translocation of nuclear factor kappa B (NF-κB) p65 into the nucleus in the LPS-induced RAW 264.7 cells. We found that the protein levels of TNF-α and IL-6 were decreased in the LPS-induced RAW 264.7 cells after the treatment with PFE in a dose-dependent manner. In addition, we confirmed that PFE inhibited the translocation of NF-κB p65 into the nucleus, as well as the protein expression levels of iNOS and COX-2. Accordingly, we pro- pose that PFE exerts an anti-inflammatory effect through the down-regulation of NF-κB p65, TNF-α, IL-6, iNOS, and COX-2 via the toll like receptor (TLR)-4 inflammatory signaling pathway.
Key words : Anti-inflammation, lipopolysaccharide, Popillia flavosellata, pro-inflammatory cytokine, RAW 264.7 cells
*Corresponding authors
*Tel : +82-63-238-2976, Fax : +82-63-238-3833
*E-mail : [email protected] (Eun-Young Yun)
*Tel : +82-54-703-7801, Fax : +82-54-770-2477
*E-mail : [email protected] (Tae-Won Goo)
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2015 Vol. 25. No. 9. 993~999 DOI : http://dx.doi.org/10.5352/JLS.2015.25.9.993
서 론
현대인은 식생활의 서구화로 인한 비만, 환경오염으로 인한 각종 오염물질 및 스트레스 등으로 인해 염증관련 질환이 증 가하고 있다[32]. 본래 염증반응은 외부 이물질(병원성 미생 물, 중금속 등)의 침입이나 상처에 대해 생체를 방어하기 위해 가장 먼저 일어나는 선천성 면역(innate immunity) 반응으로 [24], 정상적인 염증 반응은 외부 이물질 및 상처를 인식하여 대식세포(macrophage)내에서 염증매개물질인 염증성사이토 카인(pro-inflammatory cytokine), chemokine, nitric oxide (NO) 및 prostaglandin E2 (PGE2) 등에 의해 면역계가 활성화 되어 염증의 원인이 제거된 후 IL-10이나 TGF (transforming growth factor)-β와 같은 항염증 사이토카인에 의해 염증성사
이토카인이 억제되고 조직이 재생되어 종료된다[12, 24]. 염증 반응은 생체의 항상성을 유지하기 위한 매우 필수적이며 유익 한 반응이지만 염증반응 조절에 실패하여 과분비된 염증매개 물질은 세포나 조직의 기능장애 및 괴사를 일으키고 활성화된 면역세포의 생존을 계속 유지시켜 만성 염증반응의 원인이 되어 제2형 당뇨나 염증성장질환과 같은 자가면역질환 및 암 으로 발전될 수 있으므로 염증반응은 매우 적절히 조절되어야 한다[7, 8, 12, 22].
대식세포는 모든 조직과 장기에 존재하는 면역세포로서 외 부 이물질이나 상처가 생겼을 경우 이를 가장 먼저 인식하고 이동하여 급성 염증반응을 개시하는 역할을 한다[12, 23]. 마우 스 대식세포주의 일종인 RAW 264.7에 그람 음성 세균의 세포 벽 성분인 lipopolysaccharide (LPS)를 처리하면 toll like re- ceptor (TLR)-4 신호전달 과정이 활성화되어 세포질에 있던 nuclear factor kappaB (NF-κB)가 핵으로 이동 후 염증매개물 질이 생성되는데 이 생성을 억제하는 물질은 항염증 치료제 후보로 기대되기 때문에 in vitro를 이용한 항염증 물질 선별검 사에서 주로 이용되고 있다[13]. 최근 화학적으로 합성된 항염 증 제제의 장기간 복용 시 위, 신장, 심장 및 혈관에 대한 기능 저해와 같은 심각한 부작용들이 밝혀지면서 상대적으로 부작 용이 적은 천연자원으로 부터 항염증 제제를 개발하려는 노력
이 계속되고 있다[5, 10, 11]. 이러한 연구는 아직까지는 주로 식물 유래의 천연자원으로 연구되고 있지만, 최근 벼메뚜기와 같은 곤충을 이용한 항염증제에 대한 연구가 점차 증가되고 있다[21, 35].
참콩풍뎅이(Popillia flavosellata)는 딱정벌레목, 풍뎅이과의 완전변태 곤충으로 몸길이는 약 10~13 mm이고 4~11월에 출 현하며, 한국, 중국, 일본에서 서식하는 농작물에 해를 끼치는 해충이다[20]. 현재까지 참콩풍뎅이에 대한 연구는 해충 방제 를 위한 연구 외에는 기능성에 대한 보고가 전무한 실정이다 [14]. 최근 FAO보고서에 따르면, 딱정벌레목은 전 세계에서 식용 및 약용으로 가장 많이 이용되고 있는 매우 중요한 곤충 자원이다[30]. 이에 본 연구는 곤충을 이용하여 새로운 항염증 제로 개발하기 위해 LPS로 염증이 유도된 RAW 264.7 세포에 참콩풍뎅이 에탄올 추출물을 처리하여 항염증 효능을 확인하 였다.
재료 및 방법
참콩풍뎅이 및 강황 추출물 제조
추출물 제조를 위해 사용된 참콩풍뎅이(P. flavosellata) 성충 은 경북 안동시 와룡면 가야리에서 채집된 것을 사용하였고 양성대조군으로 사용한 강황은 시중에서 판매되고 있는 국내 산 건조 강황(Curcuma longa)을 사용하였다. 참콩풍뎅이 및 강 황은 흐르는 물에 2회 세척하고, 동결 건조기(Eyela, Japan)로 건조 후 다기능 분쇄기(Korea Medi, Korea)로 분쇄하여 분말 을 제조하였다. 추출물 제조를 위해 분말을 70% 에탄올에 용 해시킨 후 초음파 파쇄기(LabaX, MA, USA)로 230 jule, 20초 간 파쇄하고 4,500 rpm에서 10분 동안 원심분리한 후 회수한 상층액을 0.25 μm syringe filter (Whatman, ND, USA)로 여과 하고 건조하여 참콩풍뎅이 에탄올 추출물(P. flavosellata etha- nol extract; PFE)과 강황 에탄올 추출물(C. longa ethanol ex- tract; CLE)로 사용하였다.
세포 생존률 분석
대식세포주 RAW 264.7은 penicillin-streptomycin 100 unit/ml과 10% fetal bovine serum (Gibco, MD, USA)이 함유 된 Dublecco's Modified Eagle Medium (Gibco, MD, USA) 배지를 사용하여 37℃, 5% CO2 incubator (Thermo Scientific, IL, USA)에서 배양하였으며, 2일 간격으로 계대 배양을 실시 하였다. 세포 생존율 측정을 위해 RAW 264.7 세포를 2×104 cells/ml로 96 well plate에 분주하고 PFE를 농도별(500, 1,000, 2,000 μg/ml)로 처리 후 37℃, 5% CO2 incubator에서 24시간 배양하였다. CellTiter 96® aqueous non-radioactive cell pro- liferation assay reagent (Promega, WI, USA)를 첨가하고 3 7℃, 5% CO2 incubator에서 4시간 반응시킨 후 microplate reader (Beckman Coulter, CA, USA)를 이용하여 450 nm에서
흡광도를 측정하여 세포 생존율을 측정하였다.
Enzyme-linked immunosorbent assay (ELISA) TNF-α와 IL-6와 같은 염증성사이토카인 생성에 미치는 PFE의 효과를 검증하기 위해서 LPS (100 ng/ml)로 RAW 264.7 세포를 자극하고 동시에 PFE를 농도별(500, 1,000, 2,000 μg/ml)로 처리한 후 37℃, 5% CO2 incubator에서 24시간 배양 한 후 배양액을 회수하여 TNF-α와 IL-6 ELISA kit (Thermo Scientific, IL, USA)을 이용하여 TNF-α와 IL-6의 사이토카인 생성량을 microplate reader (Beckman Coulter, CA, USA)를 이용하여 450 nm의 파장에서 흡광도를 측정하고 농도를 계산 하였다.
Western blot 분석
NF-κB p65의 핵으로 이동을 차단 및 염증성효소인 iNOS와 COX-2 생성을 억제하는 PFE의 효과를 검증하기 위해 LPS (100 ng/ml)로 염증 유도된 RAW 264.7 세포에 PFE를 농도별 (500, 1,000, 2,000 μg/ml)로 처리하고 37℃, 5% CO2 incubator 에서 각각 12시간과 24시간 배양 후 단백질을 분리하여 Western blot 분석을 수행하였다. 세포를 PBS로 2회 세척 후 cell scraper (SPL Life Science, Korea)로 모은 다음 원심분리하 여 세포를 채취하였고 NF-κB 분석을 위한 단백질 sample은 NE-PERTM Nuclear & Cytoplasmic Extraction Kit (Thermo Scientific, IL, USA)를 이용하였고, 염증성효소의 분석을 위한 단백질 sample은 CytobusterTM Protein Extraction Reagent (Novagen, ND, USA)를 이용하여 세포를 분쇄한 후 12,000 rpm에서 15분간 원심 분리하여 상층액에 있는 단백질을 회수 하였다.
회수한 단백질은 Bio Rad Protein Assay (Bio Rad, CA, USA)로 농도를 측정하고 10 μg/ml의 농도로 맞춘 후 10%
SDS-PAGE를 수행하였다. PAGE 후 polyvinylidene di- fluoride (PVDF) membrane (GE Healthcare, NJ, USA)에 transfer하고, blotting grade blocker (Bio Rad, CA, USA)를 TBST에 녹여 5%의 농도로 만든 다음 이를 이용하여 PVDF membrane을 1시간 동안 상온에서 blocking시켰다. 1차 항체 NF-κB p65와 COX-2 (Santa Cruz, CA, USA) 및 iNOS (Abcam, MA, USA)는 각각 1:1,000의 비율로 house keeping protein인 β-tubulin (Sigma, MO, USA)은 1:10,000의 비율로 희석하여 4℃에서 밤새 반응시킨 후 다음날 tris buffered sal- ine with tween-20 (TBST; Bio Rad, CA, USA)로 15분씩 4번 세척하였다. 2차 항체 HRP-conjugated secondary antibody (Promega, WI, USA)는 1:3,000의 비율로 40분 동안 상온에서 반응시킨 후 다시 TBST로 15분씩 4번 세척하였다. Western Lightning® Plus ECL (Perkin Elmer, MA, USA)을 PVDF membrane에 처리 후 암실에서 X-ray 필름(Fujifilm, Japan)으 로 감광시켜 단백질 발현량을 확인하였다.
Fig. 1. Viability of RAW 264.7 cells after treatment with PFE (Popillia flavosellata ethanol extract) or CLE (Curcuma longa ethanol extract) for 24 hr. Cell viability was de- terminded by (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy methoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) tet- razolium inner salt (MTS) colormetric assay. PFE has no cell toxicity at 2,000 μg/ml concentration. The values are expressed as means ± SD (n=3). Statistical significance was evaluated by Duncan post-hoc test in ANOVA.
통계 분석
모든 실험은 3회 이상 수행하였고, means ± SEM으로 나타 내었으며, 통계분석법은 R statistical software (version 3.03, Lucent Technologies Inc., NJ, USA)을 이용하여 one-way ANOVA로 일차분석 후 Duncan' post-hoc test를 실시하여 재검증한 다음 p값이 0.05 미만일 때 통계적으로 유의한 것으 로 판단하였다.
결과 및 고찰
PFE와 CLE의 세포 독성 확인
PFE를 RAW 264.7 세포에 농도별(500, 1,000, 2,000 μg/ml) 로 처리한 후 MTS assay로 세포독성 및 생장율을 확인하였다.
그 결과, PFE는 최고처리농도인 2,000 μg/ml까지 통계적인 유의성이 있는 독성이 관찰되지 않았다. 본 실험에서는 항염 증 효능이 보고된 식물 천연물인 강황을 CLE로 제조 하여 양 성대조군으로 사용하였으며[2], PFE와 동일한 방법으로 RAW 264.7 세포에 처리한 결과 2,000 μg/ml의 농도까지 통계적인 유의가 있는 세포독성이 나타나지 않음을 확인하였다(Fig. 1).
따라서 본 연구에서 사용한 PFE 및 CLE의 최고 처리농도는 2,000 μg/ml로 결정하여 사용하였다.
PFE의 NF-κB p65 활성화 억제
LPS, TNF-α 및 상처에 의해 활성화되는 TLR-4 신호전달과 정에서 염증매개물질을 조절하는 염증관련 전사인자 NF-κB p65의 핵으로 이동을 차단하여 활성화 억제 여부를 확인하기 위해 LPS (100 ng/ml)로 염증 유도와 동시에 RAW 264.7 세포
에 PFE를 농도별(500, 1,000, 2,000 μg/ml)로 처리 후 12시간 경과 시 핵분획을 분리하여 NF-κB p65 항체로 Western blot을 통해 단백질 발현량을 확인하였다. 그 결과 PFE의 처리 농도 에 따라 핵분핵으로 NF-κB p65의 단백질 이동이 억제되었음 을 확인할 수 있었다(Fig. 2). NF-κB p65는 IκBα와 결합된 형태 로 세포질에 존재하며 LPS, 중금속, UV, TNF-α에 의한 신호전 달과정을 통해 IκBα가 분해된 후, 분리된 NF-κB p65가 핵으로 이동하여 염증매개물질 유전자의 전사를 활성화시킨다[3, 9, 18]. 따라서 NF-κB p65의 핵으로 이동을 억제하는 물질은 과 도한 염증으로 인한 질병에 대한 치료제로 이용되고 있다[27].
이상의 결과를 통해 PFE는 NF-κB p65의 핵으로 이동을 억제 하므로 이 전사인자 하류에 존재하는 다양한 염증매개물의 유전자 전사 또한 억제할 것이라 생각된다.
PFE의 염증성사이토카인 발현 억제
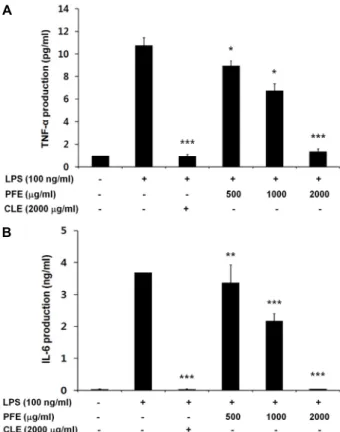
PFE의 염증성사이토카인 억제를 통한 항염증 효능을 분석 하기 위해 우선 RAW 264.7 세포에 LPS (100 ng/ml)로 염증 유도와 동시에 PFE를 농도별(500, 1,000, 2,000 μg/ml)로 처리 후 24시간 경과 시 배양액에서 염증반응의 주요 마커로 알려 진 염증성사이토카인의 단백질 발현 억제여부를 ELISA를 통 해 확인하였다(Fig. 3). 그 결과, TNF-α와 IL-6 모두 2,000 μg/
ml의 PFE 처리 시 LPS 단독처리군에 비해 TNF-α는 약 12.6 배, IL-6는 약 108.7배 감소됨을 확인할 수 있었으며, 통계적인 유의성(p<0.001)을 확인할 수 있었다.
TNF-α는 병원성 세균, 중금속 및 상처 등에 의해 TLR-4 신호전달과정에 의해 NF-κB가 활성화된 대식세포에서 주로 생성되며 염증반응에서 autocrine이나 paracrine을 통해 면역 세포의 TNF 수용체(receptor)에 결합 후 NF-κB를 활성화시켜 염증매개물질의 발현을 증가시키는 염증반응의 상류에 속하 는 염증매개물질이다[4, 28]. IL-6는 JAK-STAT 신호전달 과정 을 통해 염증매개물질의 발현 및 T세포의 활성화를 통해 후천 성면역을 개시하는 역할을 한다. 따라서 이 두 가지 염증성사 이토카인을 억제하는 물질은 염증질환에 대한 치료제로 연구 및 사용되고 있다[4, 8, 28, 33].
본 연구에서 사용한 PFE는 염증매개물 중 가장 중요한 지표 인 염증성사이토카인인 TNF-α와 IL-6를 강하게 억제하므로 항염증 효능을 갖고 있을뿐만 아니라 기존에 항염증 효능이 보고된 강황과 TNF-α와 IL-6 분비량을 비슷한 수준으로 억제 시킴을 확인할 수 있었으므로 새로운 천연 항염증 물질로 사 용될 수 있을 것이라 기대된다.
PFE의 iNOS 및 COX-2 단백질 발현 억제
LPS 처리 시 생성되는 염증성효소인 iNOS와 COX-2의 억 제를 통한 항염증 효능을 확인하기 위해 LPS로 염증 유도와 동시에 RAW 264.7 세포에 PFE를 500, 1,000, 2,000 μg/ml의 농도로 처리한 후 iNOS와 COX-2의 단백질 발현량을 Western
Fig. 2. Effect of PFE (Popillia flavosellata ethanol extract) on NF-κB p65 translocation into the nucleus in LPS-induced RAW 264.7 cells. The cell were co-treated with PFE (500, 1,000, 2,000 μg/ml) and with LPS (100 ng/ml) for 12 hr. The NF-κB p65 nucleus protein was obtained by nuclear extraction kit. Western blot analysis was show that the 2,000 μg/ml concentration of PFE blocked NF-κB p65 translocation into the nucleus.
A
B
Fig. 3. Effect of PFE (Popillia flavosellata ethanol extract) on TNF- α (A) and IL-6 (B) protein expression on in LPS-induced RAW 264.7 cells. The cells were co-treated with PFE (500, 1,000, 2,000 μg/ml) and with LPS (100 ng/ml) for 24 hr. TNF-α and IL-6 protein expression were de- creased in LPS-induced RAW 264.7 cells after treatment with PFE in a dose-dependent manner. The values are expressed as means ± SD (n=3). Statistical significance was evaluated by Duncan post-hoc test in ANOVA.
***p<0.001, **p<0.01 and *p<0.05 compared to the LPS treatment group.
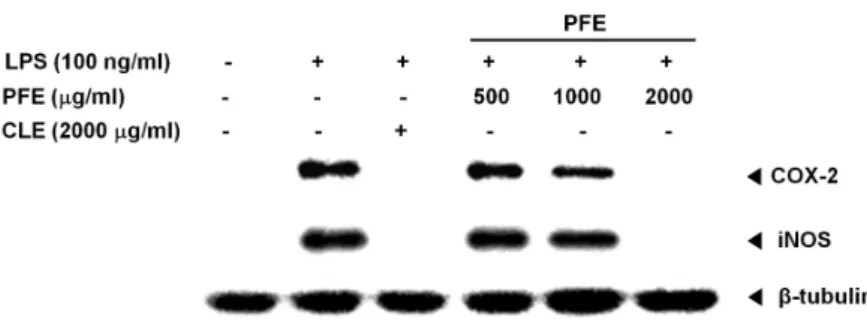
blot 분석을 통해 확인한 결과 두 단백질 모두 2,000 μg/ml의 PFE 처리 시 발현이 현저히 감소된 것을 확인하였다(Fig. 4).
NO는 일반적으로 혈압조절, 세포 신호전달, 항균 작용, 면 역세포의 활성화 등의 다양한 기능을 수행하지만[1, 22, 23, 25], 염증반응에서 과분비된 NO는 DNA의 손상, 염증 증폭,
패혈성 쇼크, 세포의 괴사, 기능장애 및 혈관세포의 투과성을 증가시켜 부종을 일으키는 등 염증반응을 촉진시키는 것으로 알려져 있다[16, 23, 25]. 염증반응에서 생성된 NO의 생성량은 iNOS의 효소의 생성량과 비례하기 때문에[26] 본 연구에서는 RAW 264.7 세포에 염증 유도와 동시에 PFE 처리 후 iNOS Western blot 분석을 통해 NO의 단백질 발현 억제를 확인하 였다. 그 결과 2,000 μg/ml PFE 처리 시 iNOS의 단백질이 거의 발현되지 않음을 확인할 수 있었으며, iNOS의 발현은 NF-κB와 TNF-α에 의해 유도되는데 PFE 2,000 μg/ml 처리 시 NF-κB p65의 활성이 완전히 차단되므로(Fig. 2) TNF-α가 생성되지 않아 iNOS의 발현 감소가 현저하게 이루어진 것으 로 추정된다(Fig. 3, Fig. 4).
LPS, TNF-α, NO, 중금속, UV, 상처 등에 의해 아라키돈산 신호전달과정에서 인지질이 분해되어 생성된 COX-2는 PGE2 를 생성시키는 효소이고, PGE2는 혈관확장, 통증 및 발열작용, 면역세포 활성화 등을 수행한다[1, 5, 19, 23]. RAW 264.7 세포 에 염증유도와 동시에 2,000 μg/ml의 PFE 처리시 COX-2가 감소된 이유는 NF-kB p65의 활성화 억제를 통해 COX-2 생성 과 관련된 TNF-α와 NO 등과 같은 염증매개물질 발현이 감소 되었기 때문으로 추정된다(Fig. 3, Fig. 4). 비스테로이드성 진 통제(non-steroidal anti-inflammatory drug; NSAID)가 COX-2 억제를 통해 진통효과가 발견된 이후[29] 다양한 NSAID가 개발되어 항염증 치료제로 널리 이용되어 왔으나 [17, 31] NSAID를 장기간 투여한 환자에 대해 COX-2가 항상 발현되는 위장, 신장에 대한 기능저해와 심혈관계 질환이 있 는 환자의 혈관확장이 저해되어 사망하는 등 심각한 부작용이 보고되고 있다[11, 15]. 따라서 최근에는 천연물 유래의 COX-2 억제제가 동물이나 임상실험을 통해 부작용이 NSAID보다 안 전하기 때문에 우유엉겅퀴 유래의 silymarin, 강황 유래의 curcumin, 천연허브 6종을 혼합하여 만든 SHINBARO 등과 같은 천연물을 이용한 항염증 치료제를 개발하려는 연구가 진행되고 있다[1, 2, 15]. 이러한 천연물 유래 항염증 치료제 연구는 현재까지는 식물을 이용해서 주로 진행되고 있지만, 최근 벼메뚜기와 같은 곤충을 이용한 항염증 연구는 점차 확 대되고 있다[21, 35].
또한 갈색거저리와 흰점박이꽃무지와 같은 딱정벌레목 곤
Fig. 4. Effect of PFE (Popillia flavosellata ethanol ex- tract) on COX-2 and iNOS protein ex- pression in LPS-induced RAW 264.7 cells.
The cell were co-treated with PFE (500, 1,000, 2,000 μg/ml) and with or without LPS (100 ng/ml) for 24 hr. The 2,000 μg/
ml concentration of PFE decreased COX-2 and iNOS protein expression.
충은 식물에 비해 불포화지방, 단백질, 무기질 등의 함량이 높아 영양적 가치가 매우 높음이 보고되었다[30]. 참콩풍뎅이 의 영양성분을 분석한 연구는 없지만 흰점박이꽃무지와 갈색 거저리와 같은 딱정벌레목 곤충이 단백질 함량이 각각 50.3%
와 57.9%, 총 지질 함량이 각각 33.7%와 16.6%를 함유하는 것이 확인되었으므로[6, 34] 참콩풍뎅이 역시 영양적 가치가 매우 우수할 것으로 예상되며 과도한 염증으로 인한 조직이나 세포의 기능 회복과 치료에 효과가 있을 것으로 생각된다.
현대인은 다양한 원인으로 인해 염증질환이 증가하고 있으 며 과도하게 분비된 염증매개물질은 비정상적인 면역반응과 세포와 조직에 괴사나 기능저해를 일으키며, 면역세포의 조직 내 침투를 증가시켜 만성염증반응의 원인이 되어 제 2형 당뇨, 염증성장질환, 류마치스 등과 같은 심각한 염증성질환을 유발 할 수 있으므로 과도하게 생성된 염증매개물질의 생성을 조절 할 수 있는 물질은 염증질환에 대한 치료제로 연구되고 있다 [7, 8, 12, 22]. 본 연구는 곤충에 대한 다양한 기능성 연구를 확대시키기 위해 참콩풍뎅이의 항염증 효능을 검증하였으며, 그 결과 TLR-4 신호전달과정의 주요 전사인자인 NF-κB p65의 활성화를 억제하고, 염증 반응을 조절하는 염증성사이토카인 TNF-α와 IL-6의 발현억제와 NF-κB p65와 염증성사이토카인 에 의해 발현량이 증가되는 염증성효소인 iNOS와 COX-2의 발현을 억제하는 항염증 효능을 갖고 있음이 확인되었으므로 곤충유래의 천연 항염증 치료제로 개발될 수 있을 것으로 기 대된다.
감사의 글
본 연구는 농촌진흥청에서 지원하는 어젠다프로그램 (PJ01099302)의 연구수행으로 인한 결과물임을 밝힙니다.
References
1. Ahmadi-Ashtiani, H. R., Rastegar, H. and Arkam, M. 2013.
The inhibitory effect of silymarin on cell viability and cel- lular COX-2 and iNOS level in HepG2 cell line. Health Med.
7, 2008-2014.
2. Arora, R. B., Kapoor, V., Basu, N. and Jain, A. P. 1971.
Anti-inflammatory studies on Curcuma longa (turmeric).
Indian J. Med. Res. 59, 1289-1295.
3. Baldwin, A. S. Jr. 1996. The NF-kappa B and I kappa B pro- teins: new discoveries and insights. Annu. Rev. Immunol. 14, 649-683.
4. Brennan, F. M., Chantry, D., Jackson, A., Maini, R. and Feldmann, M. 1989. Inhibitory effect of TNF alpha anti- bodies on synovial cell interleukin-1 production in rheuma- toid arthritis. Lancet 2, 244-247.
5. Cerella, C., Sobolewski, C., Dicato, M. and Diederich, M.
2010. Targeting COX-2 expression by natural compounds:
a promising alternative strategy to synthetic COX-2 in- hibitors for cancer chemoprevention and therapy. Biochem.
Pharmachol. 15, 1801-1815.
6. Chung, M. Y., Hwang, J. S., Goo, T. W. and Yun, E. Y. 2013.
Analysis of general composition and harmful material of Protaetia brevitarsis. J. Life Sci. 23, 664-668.
7. Ferrero-Miliani, L., Nielsen, O. H., Andersen, P. S. and Girardin, S. E. 2007. Chronic inflammation: Importance of NOD2 and NALP3 in interleukin-1beta generation. Clin.
Exp. Immunol. 147, 227-235.
8. Gabay, C. 2006. Interleukin-6 and chronic inflammation.
Arthritis Res. Ther. 8, S3.
9. Giuliani, C., Napolitano, G., Bucci, I., Montani, V. and Monaco, F. 2001. NF-κB transcription factor: role in the pathogenesis of inflammatory, autoimmune, and neoplastic diseases and therapy implications. Clin. Ter. 152, 249-253.
10. Hink, U. and Münzel, T. 2006. COX-2, another important player in the nitric oxide-endothelin cross-talk: good news for COX-2 inhibitors? Circ. Res. 98, 1344-1346.
11. Howes, L. G. 2007. Selective COX-2 inhibitors, NSAIDs and cardiovascular events - is celecoxib the safest choice? Ther.
Clin. Risk Manag. 3, 831-845.
12. Iwalewa, E. O., McGaw, L. J., Naidoo, V. and Eloff, J. N.
2007. Inflammation: the foundation of diseases and disorders. A review of phytomedicines of south african ori- gin used to treat pain and inflammatory conditions. Afr. J.
Biotechnol. 6, 2868-2885.
13. Kang, C. H., Choi, Y. H., Choi, I. W., Lee, J. D. and Kim, G. Y. 2011. Inhibition of lipopolysaccharide-induced iNOS, COX-2, and TNF-α expression by aqueous extract of Orixa Japonica in RAW 264.7 cells via suppression of NF-κB activity. Trop. J. Pharm. Res. 10, 161-168.
14. Kim, J. J., Jang, B. K., Lee, S. M., Choo, H. Y., Zhu, M. and Lee, D. W. 2011. Investigation of host plants and seasonal occurrence of Popillia flavosellata (Coleoptera: Rutelidae) in golf courses. Asian J. Turfgrass Sci. 25, 22-29.
15. Lee, S. Y., Kwon, H. K. and Lee, S. M. 2011. SHINBARO,
a new herbal medicine with multifunctional mechanism for joint disease: first therapeutic application for the treatment of osteoarthritis. Arch. Pharm. Res. 34, 1773-1777.
16. Miyataka, M., Rich, K. A., Ingram, M., Yamamoto, T. and Bing, R. J. 2002. Nitric oxide, anti-inflammatory drugs on renal prostaglandins and cyclooxygenase-2. Hypertension 39, 785-789.
17. Mitchell, J. A., Akarasereenont, P., Thiemermann, C., Flower, R. J. and Vane, J. R. 1993. Selectivity of nonsteroid anti-inflammatory drugs as inhibitors of constitutive and in- ducible cyclo-oxygenase. Proc. Nat. Acad. Sci. USA 90, 11693- 11697.
18. Monaco, C., Andreakos, E., Kiriakidis, S., Mauri, C., Bicknell, C., Foxwell, B., Cheshire, N., Paleolog, E. and Feldmann, M. 2004. Canonical pathway of nuclear factor kappa B acti- vation selectively regulates proinflammatory and pro- thrombotic responses in human atherosclerosis. Proc. Nat.
Acad. Sci. USA 101, 5634-5639.
19. Nakanishi, M. and Rosenberg, D. W. 2013. Multifaceted roles of PGE2 in inflammation and cancer. Semin. Immunopa- thol. 35, 123-137.
20. Paek, M. K., Hwang, J. M., Jung, K. S., Kim, T. W., Kim, M. C., Lee, Y. J., Cho, Y. B., Park, S. W., Lee, H. S., Ku, D. S., Jeong, J. C., Kim, K. G., Choi, D. S., Shin, E. H., Jwang, J. H., Lee, J. S., Kim, S. S. and Bea, Y. S. 2010. Checklist of Korean insects. Nature & Ecology, pp. 113.
21. Park, J. Y., Heo, J. C., Woo, S. U., Yun, C. Y., Kang S. W., Hwang, J. S. and Lee, S. H. 2006. Anti-inflammatory and cellular protective effects on hydrogen peroxide-induced cy- totoxicity of grasshopper extracts. Kor. J. Food Preserv. 13, 796-802.
22. Rajendram, R., Saraswathy, S. and Rao, N. A. 2007. Photor- eceptor mitochondrial oxidative stress in early experimental autoimmune uveoretinitis. Br. J. Ophthalmol. 91, 531-537.
23. Rao, C. V., Indranie, C., Simi, B., Manning, P. T., Connor, J. R. and Reddy, B. S. 2002. Chemopreventive properties of a selective inducible nitric oxide synthase inhibitor in colon carcinogenesis, administered alone or in combination with celecoxib, a selective cyclooxygenase-2 inhibitor. Cancer Res.
62, 165-170.
24. Medzhitov, R. 2008. Origin and physiological roles of inflammation. Nature 454, 428-435.
25. Salvemini, D., Milko, T. P., Masferrer, J. L., Seibert, K., Currie, M. G. and Needleman, P. 1993. Nitric oxide activates cyclooxygenase enzymes. Proc. Nat. Acad. Sci. USA 90, 7240-7244.
26. Shafran, Y., Zurgil, N., Afrimzon, E., Tauber, Y., Sobolev, M., Shainberg, A. and Deutsch, M. 2012. Correlative analy- ses of nitric oxide generation rates and nitric oxide synthase levels in individual cells using a modular cell-retaining device. Anal. Chem. 84, 7315-7322.
27. Tak, P. P. and Firestein, G. S. 2001. NF-kappaB: a key role in inflammatory diseases. J. Clin. Invest. 107, 7-11.
28. Tracey, D., Klareskog, L., Sasso, E. H., Salfeld, J. G. and Tak, P. P. 2008. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol. Ther. 117, 244- 279.
29. Swierkosz, T. A., Mitchell, J. A., Warner, T. D., Botting, R.
M. and Vane, J. R. 1995. Co-induction of nitric oxide syn- thase and cyclo-oxygenase: interactions between nitric oxide and prostanoids. Br. J. Pharmacol. 114, 1335-1342.
30. Van Huis, A., Van ltterbeeck, J., Klunder, H., Mertens, E., Halloran. A., Muir, G. and Vantomme, P. 2013. Edible in- sects: future prospects for food and feed security. Food and agriculture Organization of the United Nations, Rome, Italy.
31. Vane, J. R. 1971. Inhibition of prostaglandin synthesis as a mechanism of action for the aspirin-like drugs. Nature 231, 232-235.
32. Vlassara, H. 2005. Advanced glycation in health and disease role of the modern environment. Ann. N. Y. Acad. Sci. 1043, 452-460.
33. Wang, X., Luo, Y., Liao, W. B., Zhang, J. and Chen, T. M.
2013. Effect of osteoprotegerin in combination with inter- leukin-6 on inhibition of osteoclast differentiation. Chin. J.
Traumatol. 16, 277-280.
34. Yoo, J. M., Hwang, J. S., Goo, T. W. and Yun, E. Y. 2013.
Comparative analysis of nutritional and harmful compo- nents in Korean and Chinese mealworms (Tenebrio molitor).
J. Kor. Soc. Food Sci. Nutr. 42, 249-254.
35. Yoon, Y. I., Chung, M. Y., Hwang, J. S., Goo, T. W., Ahn, M. Y., Lee, Y. B., Han, M. S. and Yun, E. Y. 2014. Anti-in- flammatory effect of Oxya chinensis sinuosa ethanol extract in LPS-induced RAW 264.7 cells. J. Life Sci. 24, 370-376.
초록:LPS로 염증 유도된 RAW 264.7세포에 대한 참콩풍뎅이(Popillia flavosellata) 에탄올 추출 물의 항염증 효과
윤영일1․황재삼1․김미애1․안미영1․이영보1․한명세2․구태원3*․윤은영1*
(1농촌진흥청 국립농업과학원 농업생물부, 2경북대학교 농업생명과학대학 바이오섬유소재학과, 3동국대학교 의과
학대학 생화학교실
본 연구에서는 참콩풍뎅이(Popillia flavosellata) 에탄올 추출물(PFE)의 항염증 효능을 분석하기 위해 PFE를 농도 별(500, 1,000, 2,000 μg/ml)로 대식세포인 RAW 264.7에 처리 시 최고 처리농도인 2,000 μg/ml까지 통계적인 유의 성 있는 독성이 없음을 확인하였다. LPS (100 ng/ml)로 염증 유도된 RAW 264.7 세포에 PFE를 농도별(500, 1,000, 2,000 μg/ml)로 동시 처리 시 농도 의존적으로 염증성사이토카인인 TNF-α와 IL-6의 단백질 생성을 통계적인 유의 성(p<0.001)있게 억제함을 확인하였다. 또한 염증 유도된 RAW 264.7 세포에 PFE 동시 처리 시 NF-κB p65의 핵으 로 이동이 차단됨과 iNOS와 COX-2의 단백질 발현을 감소시키는 것을 확인하였다. 이상의 연구결과를 통해 참콩 풍뎅이는 염증에 의해 활성화된 TLR-4 신호전달과정을 조절하는 NF-κB p65의 활성과 염증성사이토카인 TNF-α 와 IL-6의 생성 및 염증성효소 iNOS와 COX-2의 생성을 억제하는 항염증 효능이 있음을 확인하였다.