Bacillus velezensis L2를 이용하여 제조한 멸치메주의 품질 특성
박지선1․이호우2․서예슬1․양은주1
1(재)전남생물산업진흥원 식품산업연구센터
2청우F&B
Quality Characteristics of Anchovy-Meju Fermented with Bacillus velezensis L2
Ji Sun Park1, Ho Woo Lee2, Ye-Seul Seo1, and Eun Ju Yang1
1Food Research Center, Jeonnam Bioindustry Foundation
2Chungwoo F&B Co.
ABSTRACT To develop a starter of anchovy-meju, soybeans with anchovy were fermented using four strains isolated from traditional fermented food. Among them, a strain, L2, which showed high protease activity and cell numbers in the fermented sample, was selected as a starter. The selected strain L2 was identified as Bacillus velezensis based on 16S rRNA sequencing. Anchovy-meju was prepared using B. velezensis L2, and its quality properties were compared with those of the control (without a starter) during fermentation. Microbial analysis of the starter anchovy-meju revealed B. velezensis L2 to be the dominant organism after 30 days of fermentation. After 30 days of fermentation, the protease activities of control and starter anchovy-meju were 4,880 unit/g and 6,565 unit/g, respectively. The content of amino-type nitrogen was higher in the starter anchovy-meju (953.2 mg%) compared to that in the control anchovy-meju (711.7 mg%) after 30 days of fermentation. The pH increased, and the acidity decreased gradually during fermentation. In the case of color, the L and b values increased gradually in the starter anchovy-meju, while it decreased gradually in the control anchovy-meju during fermentation. The concentration of glutamic acid, which imparts a savory flavor to the meju, was 224.02 mg% in the starter anchovy-meju, which is more than 99.94 mg% of the control.
Key words: meju, anchovy, Bacillus velezensis, fermentation, quality characteristics
Received 16 June 2020; Accepted 3 August 2020
Corresponding author: Eun Ju Yang, Food Research Center, Jeon- nam Bioindustry Foundation, Naju, Jeonnam 58275, Korea E-mail: [email protected], Phone: +82-61-339-1251
서 론
전통된장은 한국의 대표적인 대두 발효식품으로서 필수 아미노산, 지방산, 유기산, 미네랄, 비타민 등을 보충해 줄 수 있어 우수한 영양원이 될 뿐만 아니라(Kim 등, 2011) 항산화 활성(Oh 등, 2014), 혈전 용해(Hyun 등, 2005), 항 암 효과(Kwon과 Shon, 2004), 항돌연변이 활성(Park 등, 1996) 및 항고혈압 활성(Yu 등, 1996) 등 다양한 생리활성 이 보고되어 있다. 또한 발효 및 숙성과정에서 생성되는 아 미노산, 펩타이드, 유리당 등에 의해 구수한 맛과 독특한 향 미를 형성하므로 한식에서 중요한 조미식품으로 식생활에 이용되고 있다(Lee 등, 2012). 최근에는 장류 시장이 정체되 면서 전통 발효식품인 된장의 소비 촉진 및 현대인의 기호에 맞는 새로운 제품을 개발하기 위해 녹차(Park 등, 2005), 다시마(Cui 등, 2002), 고구마(Bae 등, 2012), 마늘(Kang 등, 2014), 여주(Hwang 등, 2017) 등 다양한 소재를 첨가하
여 맛과 기능성이 증진된 고품질 된장을 개발하기 위한 노력 이 진행되고 있다. 또한 전통된장은 제조장소 및 시기에 따 라 품질이 균일하지 못하고 발효 중 혼입되는 잡균으로 인해 위생성과 안전성의 문제가 제기되면서 개량식 된장과는 차 별화하여 전통된장에 적합한 종균 개발 및 적용 연구도 진행 되고 있다(Cho 등, 2016; Rhee 등, 2008). 그러나 메주를 포함한 전통된장의 연구에서 대두 외에 새로운 소재를 첨가 하면서 종균 발효를 시도한 연구는 극히 소수에 불과하다 (Lee 등, 2012).
멸치는 우리나라 연안에서 연간 평균 18,000톤가량 어획 되는 어종으로(KOSTAT, 2018) 칼슘이 풍부하고 양질의 아미노산과 각종 무기질 및 정미성분 등을 함유하고 있어 가공식품의 원료로 잠재적 가치가 높은 수산자원이다(Kang 등, 2016). 그러나 우리나라에서 멸치의 이용은 조리 음식의 육수를 내는 데 사용하거나 멸치 액젓을 제조하여 김치의 부재료 및 조미료로 활용하는 형태로 사용이 매우 제한적이 고 소비가 정체되어 있다(Kwak 등, 2010). 멸치의 이용도를 높이기 위해 멸치의 식품 소재화 필요성이 증가하면서 멸치 를 발효한 분말조미료 소재(Jo 등, 1999), 멸치를 함유한 고칼슘 어묵(Bae 등, 2007), 멸치를 이용한 스낵제품 개발
(Kang 등, 2016) 등이 진행되어 왔다. 또한 영양 및 기능성 이 향상된 된장의 개발을 위해 생된장에 멸치분말을 첨가하 여 숙성한 결과, 품질 지표인 아미노산성 질소와 관능 기호 도가 우수하였다는 연구 결과도 보고되어(Kim 등, 2004) 멸치가 장류 소재로 활용될 수 있음을 확인하였다. 그러나 전통된장의 제조 방법에 따라 메주 제조 단계에서 멸치를 첨가하여 품질 특성을 분석한 연구는 전무하다.
메주는 전통된장의 주원료로서 대두를 수침, 자숙 및 성 형한 후 1~2개월 발효 시켜 만든 것을 말한다. 한식메주의 경우 볏짚 등을 이용하여 자연 발효하는 과정 중 Bacillus subtilis나 Aspergillus oryzae 등의 미생물이 증식하여 아 미노산, 당, 유기산 등 풍미 성분을 생성하고, 이렇게 제조된 메주에 식염 등을 섞어 발효 및 숙성을 통해 전통된장 등의 발효식품을 생산한다(Cho 등, 2016; MFDS, 2020).
본 연구에서는 전통된장의 원료로서 정미 특성과 품질이 우수한 메주의 개발을 위해 멸치분말을 첨가한 메주를 제조 하였으며, 대두 및 멸치 혼합 원료에 대한 발효능이 우수한 Bacillus velezensis L2를 멸치메주의 종균으로 활용하여 발효에 따른 종균의 점유율, protease 활성, 아미노태 질소 함량, pH, 산도, 색도의 변화를 조사하고, 유리아미노산을 분석하여 종균을 접종하지 않은 멸치메주와 비교하였다.
재료 및 방법
균주의 분리 및 protease 활성 측정
Protease 활성이 우수한 균주를 분리하기 위해 가정에서 재래식으로 제조한 된장 3종과 멸치액젓(Chungwoo F&B, Yeonggwang, Korea) 시료 1 g을 각각 멸균수 10 mL에 현탁한 희석액을 준비한 후 2% skim milk를 첨가한 Tryp- tic Soy Agar(TSA, Difco Laboratories, Detroit, MI, USA) 배지에 도말하였다. 도말한 배지는 37°C에서 24시간 배양 한 후 시료당 skim milk 분해환을 가장 크게 형성하는 균주 를 선별하였다.
선별된 균의 protease 활성은 azocasein법을 일부 변형 하여 측정하였다(Kim 등, 1998). 분리균주를 Tryptic Soy Broth(TSB, Difco Laboratories) 액체배지에 접종하여 37°C에서 24시간 동안 배양한 후 배양액을 원심분리(8,000 rpm, 10 min, Micro 17TR, Hanil Science Inc., Gimpo, Korea)하여 얻은 상등액을 조효소액으로 사용하였다. 기질 용액은 100 mM sodium phosphate buffer(pH 7.0)에 azo- casein(Sigma-Aldrich, St. Louis, MO, USA)의 농도가 0.5%(w/v)가 되도록 용해하여 사용하였다. 기질용액 150 μL에 조효소액 50 μL를 혼합하여 30°C에서 1시간 동안 반 응시킨 후 10% trichloroacetic acid 400 μL를 첨가하여 반응을 종료하였다. 반응액을 원심분리(10,000 rpm, 10 min)한 후 상등액 500 μL에 525 mM NaOH 700 μL를 가하 여 혼합한 다음 432 nm에서 흡광도(PowerWave XS, Bio- Tek Instruments Inc., Winooski, VT, USA)를 측정하였
다. 효소활성은 30°C에서 1시간 동안 효소반응으로 유리되 는 azo기에 의해 432 nm에서 흡광도를 0.01 증가시키는 양 을 1 unit/mL로 정의하였다.
발효 종균의 선정 및 동정
멸치분말을 첨가한 메주의 발효 종균을 선정하기 위해 분 리균 4종을 이용하여 멸치분말이 첨가된 대두를 각각 발효 시킨 후 발효 시료의 protease 활성과 발효균 수를 측정하 였다. 함평 농협에서 구입한 대두를 실온에서 24시간 수침 후 100°C에서 1시간 증자하고 마쇄하였다. 증자 대두 90 g에 멸치분말(Chamhanfood, Yeonchen, Korea) 10 g을 혼합한 후 미리 배양한 분리균 4종의 배양액을 1%(v/w) 접 종하여 30°C에서 3일간 발효하였다. 발효 시료의 protease 활성은 각 시료 1 g을 증류수 4 mL로 현탁한 후 원심분리 (8,000 rpm, 10 min)하여 얻은 상등액을 조효소액으로 사 용하여 상기와 동일한 방법으로 측정하였다. 발효 시료의 발효균 수를 측정하기 위해 각 시료 1 g을 멸균 생리식염수 를 이용하여 10배씩 단계적으로 희석하였다. 희석액을 TSA 배지에 도말하여 30°C에서 24시간 배양한 후 균주의 집락 을 계수하여 발효균 수를 측정하였다.
발효 종균으로 선정된 L2 균주의 동정을 위해 universal primer인 785F(5′-GGATTAGATACCCTGGTA-3′)와 907R(5′-CCGTCAATTCMTTTRAGTTT-3′)을 이용하여 16S rRNA 염기서열을 결정한 후 NCBI(National Center for Biotechnology Information)의 Blastn 프로그램을 이 용하여 GenBank에 등록된 염기서열들과 비교했으며, 표준 균주(type strain)와의 상동성은 Mega version 6.06 프로 그램(http://www.megasoftware.net/)의 ClustalW를 이용 하여 분석하였다(Saitou와 Nei, 1987).
종균을 이용한 멸치메주 제조
대두를 실온에서 24시간 수침하고 100°C에서 1시간 증 자한 후 증자 대두 53.4 kg과 멸치분말 6 kg을 혼합하였다.
멸치분말이 첨가된 대두에 미리 배양한 L2 종균을 1%(v/w) 접종한 후 메주 하나당 3 kg 중량(9.5×13.5×20.5 cm)으로 사각 멸치메주를 성형하였으며, 대조군 멸치메주는 L2 종균 을 접종하지 않고 동일한 규격으로 성형하였다. 성형한 멸치 메주는 40°C에서 24시간 건조한 후, 30°C에서 30일 동안 발효시키면서 10일 간격으로 시료를 채취하여 분석시료로 사용하였다. 멸치메주의 protease 활성과 발효균 수 분석은 발효 종균 선정을 위한 발효 시료 분석과 동일한 방법으로 측정하였다.
멸치메주의 pH 및 산도 측정
멸치메주 시료 1 g에 증류수 100 mL를 가하여 균질화한 후 3,000 rpm에서 20분 동안 원심분리한 상등액을 시료액 으로 사용하였다. pH는 시료액을 pH meter(Mettler Tole- do GmbH, Greifensee, Switzerland)를 사용하여 측정하였
0 500 1000 1500 2000 2500
TSD YSD GG2 L2
Isolated strains
Protease activity (unit/mL) .
b
b
a
a
Fig. 1. Protease activity in the culture supernatant of isolated strains. Means with different letters (a,b) above the bars are significantly different (P<0.05).
0 2000 4000 6000 8000
Isolated strains
Protease activity (unit/g) .
0 5 10 15 Protease activity
Viable cell number
b b c c d b
a a
TSD YSD GG2 L2
Viable cell number (Log CFU/g) .
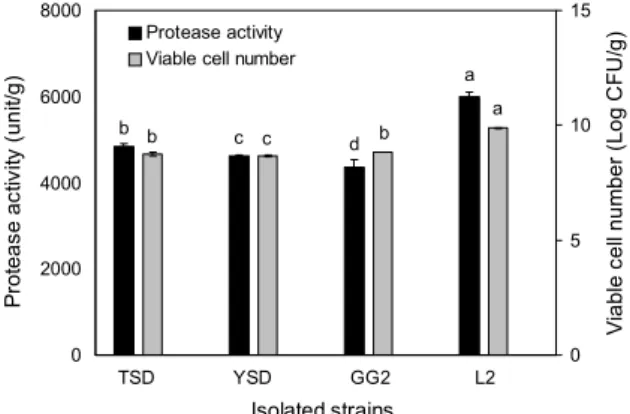
Fig. 2. Protease activity and viable cell number of soybeans with anchovy fermented using isolated strains. Means with different letters (a-d) above the bars are significantly different (P<0.05).
다. 산도는 식품공전(MFDS, 2020) 상의 시험법에 따라 측 정했으며, 시료액 10 mL를 취하여 1% 페놀프탈레인 지시 약을 약 2~3방울 가한 후 0.1 NaOH로 pH 8.3(미홍색)이 될 때까지 적정한 다음 소비된 NaOH의 부피를 젖산량으로 환산하여 계산하였다.
멸치메주의 아미노태 질소 측정
아미노태 질소 함량은 식품공전(MFDS, 2020)의 formol 적정법으로 측정하였다. 산도 분석을 위해 준비한 시료액 5 mL, 중성 포르말린 용액 10 mL, 증류수 10 mL를 넣은 플라스크에 1% 페놀프탈레인 용액을 2~3방울 가한 후 0.1 N NaOH로 pH 8.4(미홍색)가 될 때까지 적정하였다. 이때 소모된 0.1 N NaOH의 mL 수를 아미노태 질소 함량으로 결정하였다. 대조구는 중성 포르말린 용액 대신 증류수를 사 용하여 측정하였다.
멸치메주의 색도
색도는 색차계(CM-3500d, Konica Minolta, Tokyo, Ja- pan)를 이용하여 측정했으며, 분쇄한 멸치메주 시료 10 g을 petri dish에 담아 L(명도, lightness), a(적색도, redness), b(황색도, yellowness)값을 측정하여 나타내었다. 이때 사 용한 표준 백색판(Standard plate) 값은 각각 L=99.03, a=
-0.19, b=-0.29였다.
멸치메주의 유리아미노산 분석
유리아미노산은 식품공전(MFDS, 2020)의 유리아미노 산 시험법에 준하여 측정하였다. 멸치메주 시료 1 g에 70%
에탄올 15 mL를 가하여 30분간 진탕한 후 원심분리(8,000 rpm, 15 min)하였다. 남은 잔사를 다시 2회 반복 추출하고 원심분리한 상등액을 모두 합하여 감압 농축하였다. 농축액 을 0.02 N HCl 20 mL에 용해하여 0.45 μm syringe filter (Advantec Toyo Kaisha Ltd., Tokyo, Japan)로 여과한 후 분석 시료로 사용하였다. 분석은 injection volume 20 μL, flow rate 0.3 mL/min으로 하여 아미노산 자동분석기(L- 8900, Hitachi Ltd., Tokyo, Japan)로 분석하였다.
통계분석
모든 실험은 3회 반복하여 수행되었으며 실험 결과는 평 균값±표준편차(mean±SD)로 나타내었다. 처리군과 대조군 간의 차이는 SPSS program(Statistical Package for the Social Sciences, version 17.0, SPSS Inc., Chicago, IL, USA)을 이용하여 Student t-test 분석을 통해 분석하였고, 3군 이상에서의 유의적 차이 비교 및 발효일에 따른 차이는 일원배치 분산분석(one-way ANOVA)을 실시한 후 Dun- can’s multiple range test로 사후 검증하였다(P<0.05).
결과 및 고찰
Protease 활성이 우수한 균주의 선별 및 동정
Skim milk 2%가 포함된 TSA 배지에 전통된장 3종 및 멸치액젓 희석액을 각각 도말하여 평판 배양한 후 skim milk 분해환을 가장 크게 형성하는 집락을 시료별로 1종씩 선정 하였다. 된장 3종에서 분리한 protease 활성 균주는 각각 TSD, YSD, GG2로, 멸치액젓에서 분리한 protease 활성 균 주는 L2로 명명한 후 TSB 배지에 배양하여 배양 상등액의 protease 활성을 비교한 결과 GG2가 1,855 unit/mL, L2가 1,655 unit/mL로 TSD(1,046 unit/mL)와 YSD(1,176 unit/
mL)에 비해 더 우수한 활성을 보였다(Fig. 1). 4종의 분리균 주를 대상으로 멸치분말 첨가 메주(멸치메주)의 발효에 적 합한 균주를 최종 선정하기 위해 멸치분말 10%를 첨가한 증자 대두에 각각의 분리균주를 1% 접종하여 3일 동안 발효 시킨 후 발효 시료의 protease 활성과 발효균 수를 측정하 였다. Fig. 2와 같이 멸치분말이 첨가된 대두 발효액에서 접종 균주에 따른 protease 활성은 L2가 5,990 unit/g으로 가장 높았으며 발효균 수에서도 L2가 9.9 Log CFU/g으로 다른 3종의 균주에 비해 우수한 생육도를 나타내었기 때문
Table 1. Changes in the viable cell count of anchovy-meju during fermentation for 30 days Viable cell count (Log CFU/g) Fermentation time (days)
0 10 20 30
Control Total cell count 8.34±0.06Ca1) 10.41±0.02Aa 8.79±0.06Bb 7.11±0.07Db Starter
Total cell count B. velezensis L2
Dominance of starter (%)
7.03±0.02Db 6.25±0.02B
17%
9.15±0.03Ab 8.95±0.03A
64%
8.97±0.03Ba 8.84±0.02A
75%
8.89±0.01Ca 8.70±0.58A
71%
Values are expressed as the mean±SD (n=3).
1)Means with different letters within the same row (A-D) and column (a,b) are significantly different (P<0.05).
Fig. 3. Phylogenetic tree based on 16S rRNA sequences of isolated strain L2 and related species.
에 멸치메주의 발효균으로 L2를 최종 선정하였다. L2 균주 의 동정을 위해 16S rRNA 염기서열을 분석하여 GenBank 에 등록된 표준 균주들과 염기서열을 비교한 결과, L2 균주 는 Bacillus velezensis NRRL B-41580과 99%의 상동성 을 나타내었다(Fig. 3). B. velezensis는 전통간장 및 멍게 젓갈 등 발효식품에서 분리된 바 있으며(Chang 등, 2012;
Moon 등, 2018; Yao 등, 2019) 내염성, 항균 활성 및 우수 한 단백분해 활성을 나타내는 연구 결과들이 보고되었다 (Chang 등, 2012; Meena 등, 2018; Ye 등, 2018). 본 연구 에서 분리된 B. velezensis L2 균주는 멸치와 대두의 혼합 원료에서 우수한 생육도와 protease 활성을 나타내므로 아 미노산 생성능이 우수한 발효 종균으로 활용 가능성이 높을 것으로 생각된다.
멸치메주 발효에 따른 미생물 수 변화
Protease 활성이 우수하며 발효 종균으로 선정된 B. ve- lezensis L2를 이용하여 멸치분말이 첨가된 메주를 제조하 고, 제조한 멸치메주를 30일간 발효시키면서 10일 간격으 로 미생물 수의 변화를 측정하여 종균을 첨가하지 않은 대조 구 멸치메주와 비교하였다(Table 1). 대조구 멸치메주에서 총 균수는 발효 초기에 8.34 Log CFU/g에서 발효 10일 후에 10.41 Log CFU/g으로 증가했으며, 이후 점진적으로 감소하여 발효 30일에는 7.11 Log CFU/g을 나타내었다.
L2 균주로 발효한 종균 멸치메주의 총 균수는 발효 초기에
7.03 Log CFU/g에서 발효 10일 후에 9.15 Log CFU/g으로 증가했으며, 이후 조금씩 감소하여 발효 30일에는 8.89 Log CFU/g을 나타내었다. Yoo와 Kim(1998)은 순창지역의 메 주 발효 중 호기성 세균수는 발효 초기 1.29×107 CFU/g에 서 발효 20일에는 2.4×108 CFU/g으로 증가하다가 서서히 감소하여 발효 60일에는 4.07×106 CFU/g을 나타낸 결과 를 보고하였다. 반면 Yoo 등(1998)이 강원도 재래식 메주의 발효 중 품질 변화를 연구한 다른 보고에서는 메주의 호기성 세균수가 발효 초기부터 발효 70일까지 108 CFU/g 정도의 수준을 유지하는 결과를 보여 메주 발효에서 미생물의 변화 는 발효 환경 및 조건에 따라 큰 차이를 나타내는 것으로 생각된다. 본 연구에서 종균 멸치메주의 발효 기간에 B. ve- lezensis L2의 점유율을 분석한 결과 발효 초기에는 6.25 Log CFU/g으로 17%의 점유율을 보였으나, 이후 종균 수와 점유율 모두 증가하는 경향을 나타내며 발효 20일에 75%로 가장 높은 점유율을 나타내었다(Table 1). Cho 등(2016)은 B. amyloliquefaciens를 포함한 4종의 혼합 종균이 첨가된 메주의 미생물 분석 결과 bacilli 균수는 초기 3×106 CFU/g 에서 7일 후 8×108 CFU/g으로 증가한 결과를 보고하였으 나 종균 점유율은 확인되지 않았다. 멸치메주의 발효에서 B. velezensis L2의 우점은 발효 품질을 저하시킬 수 있는 잡 균의 오염 및 증식을 억제하여 멸치메주의 품질을 안정적으 로 유지할 수 있는 우수한 특성으로 생각된다.
0 2000 4000 6000 8000
0 10 20 30
Fermentation time (days) Protease activity (unit/g) . Control
Starter
Fig. 4. Changes in the protease activity of anchovy-meju during fermentation for 30 days.
0 200 400 600 800 1000
0 10 20 30
Fermentation time (days) Amino type nitrogen (mg%) . Control
Starter
Fig. 5. Changes in amino-type nitrogen content of anchovy-meju during fermentation for 30 days.
멸치메주 발효에 따른 protease 활성 변화
장류의 발효에서 protease는 발효에 관여하는 미생물을 생산하며 단백질의 소화성과 영양성 개선에 큰 역할을 하고, 아미노태 질소 및 유리아미노산 함량에 영향을 주어 특유의 맛을 내는 데 중요한 역할을 하는 것으로 알려져 있다(Rhee 등, 2008; Jung 등, 2009). 대조구 멸치메주와 종균 멸치메 주의 발효에 따른 protease 활성 변화를 측정한 결과는 Fig.
4와 같다. 대조구 멸치메주는 발효 초기 266 unit/g에서 발 효가 진행될수록 크게 증가하여 발효 20일에 5,260 unit/g 으로 가장 높은 활성을 보인 후 발효 30일에 4,880 unit/g으 로 감소하였다. 반면 종균 멸치메주는 발효 초기부터 1,346 unit/g으로 대조구에 비해 높은 활성을 나타내었으며, 발효 가 진행될수록 증가하여 발효 20일과 30일에 각각 6,510 unit/g과 6,565 unit/g으로 높은 protease 활성을 유지하였 다. Rhee 등(2008)은 B. subtilis를 종균으로 첨가한 메주로 제조한 된장의 발효에 따른 protease 활성 변화를 측정한 결과 전체 발효 기간에 대조구보다 종균된장의 효소 활성이 높은 결과를 보였다. Jung 등(2009)의 연구에서도 된장 발 효균의 protease 활성이 높을수록 발효 된장의 protease 활성도 높아지는 결과를 보이므로 protease 활성이 높은 균주로 된장을 제조할 경우 재래식 된장 고유의 풍미를 유지 하면서 속성 발효가 가능할 것으로 보고하였다.
멸치메주 발효에 따른 아미노태 질소 함량 변화
아미노태 질소는 장류 발효식품의 숙성도와 품질 수준에 대한 중요한 지표로서 발효숙성 중 미생물이 생산하는 pro- tease 작용으로 인해 단백질이 가수분해되면서 증가하여 발 효식품의 구수한 맛의 척도가 되고 있다(Lee 등, 2014; Lee 등, 2016). B. velezensis L2를 이용한 종균 멸치메주의 발 효에 따른 아미노태 질소 함량을 대조구 멸치메주와 비교한 결과는 Fig. 5와 같다. 대조구와 종균 멸치메주 모두 발효에 따라 아미노태 질소가 크게 증가하였으나 발효 10일부터 종균 멸치메주의 아미노태 질소 함량이 더 높게 측정되었다.
발효 30일에 대조구와 종균 멸치메주의 아미노태 질소 함량 은 각각 711.7 mg%와 953.2 mg%로 큰 차이를 나타내었다.
이러한 결과는 종균 멸치메주에서 발효가 진행될수록 B. velezensis L2가 생산하는 protease의 활성으로 멸치와 대 두 단백질의 가수분해가 촉진되었기 때문으로 생각된다.
Shin 등(2018)의 연구에서 시판 전통메주들의 아미노태 질 소 함량을 비교한 결과 시료에 따라 257.29~839.58 mg%
로 유의적으로 큰 차이가 있었으며, 이러한 결과는 메주의 원료인 콩의 품종이나 메주 발효 시 숙성 정도의 차이뿐만 아니라 메주의 발효 과정 중 미생물의 종류나 미생물이 생산 하는 효소작용 조건이 메주마다 다르기 때문이라고 보고하 였다. Cho 등(2016)은 복합종균을 접종하여 발효한 메주의 아미노태 질소 함량을 분석한 결과, 발효 21일까지는 접종 한 종균의 단백질 분해능에 의해 메주의 아미노태 질소 함량 이 큰 차이를 나타내었으나, 발효 28일 이후에는 유의적인 차이가 나타나지 않았다. 본 연구에서 B. velezensis L2를 이용하여 제조한 멸치메주의 아미노태 질소 함량은 발효 30 일에 953.2 mg%의 높은 함량을 나타내며 대조구뿐만 아니 라 전통식품 표준규격(MAFRA, 2018)에 제시된 메주의 아 미노태 질소 함량(110.0 mg% 이상)보다 월등하게 높은 결 과를 보였다. 아미노태 질소는 발효식품의 풍미와 감칠맛에 영향을 주며 그 함량이 높은 장류가 성분 측면에서 좋은 제 품으로 평가되므로(Hwang, 2010; Rho 등, 2008), B. ve- lezensis L2를 이용하여 발효한 멸치메주는 품질 특성이 우 수하면서 관능적으로 기호도가 더 높을 것으로 기대된다.
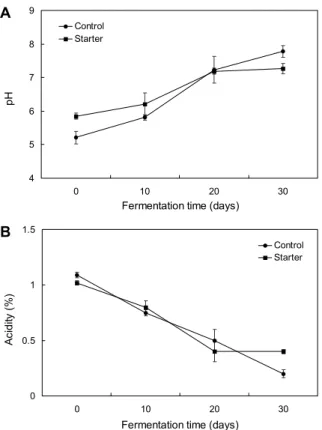
멸치메주 발효에 따른 pH 및 산도 변화
대조구와 종균 멸치메주의 발효에 따른 pH와 산도 변화 는 Fig. 6과 같다. 대조구 멸치메주의 pH는 발효 초기 5.21 에서 발효에 따라 지속적으로 증가하여 발효 30일에 7.78로 나타났다. 종균 멸치메주의 pH는 발효 초기에 5.85로 대조 구에 비해 유의적으로 높았으며, 발효에 따라 증가하여 발효 20일에 7.19를 나타내었으나 발효 30일에는 7.27로 유사한 수준을 보였다. 산도는 pH와 반대의 경향으로 발효에 따라 감소하였으며, 발효 초기에는 대조구 멸치메주의 산도가 1.1%로 종균 멸치메주에 비해 좀 더 높았으나 발효 30일에 는 대조구 멸치메주의 산도가 0.2%로 종균 멸치메주(0.4%)
Table 2. Changes in color of anchovy-meju during fermentation for 30 days
Color value Anchovy-meju Fermentation time (days)
0 10 20 30
L Control
Starter
52.82±0.34A1) 53.55±0.49B
51.99±0.70AB 53.14±0.21B
50.98±0.25Bb 53.18±0.18Ba
49.72±0.84Cb 55.07±0.60Aa
a Control
Starter
6.92±0.12Ca 6.61±0.04Cb
7.48±0.06B 7.33±0.16B
7.57±0.01B 7.46±0.08B
7.93±0.20A 7.71±0.06A
b Control
Starter
17.85±0.45A 17.93±0.27B
17.51±0.04Ab 18.06±0.18Ba
16.67±0.17Bb 18.21±0.15Ba
16.64±0.44Bb 20.07±0.08Aa Values are expressed as the mean±SD (n=3).
1)Means with different letters within the same row (A-D) and column (a,b) are significantly different (P<0.05).
4 5 6 7 8 9
0 10 20 30
Fermentation time (days)
pH .
Control Starter
A
0 0.5 1 1.5
0 10 20 30
Fermentation time (days)
Acidity (%) .
Control Starter
B
Fig. 6. Changes in pH (A) and acidity (B) of anchovy-meju during fermentation for 30 days.
Fig. 7. Appearance and cross section of anchovy-meju fermented without (A and B) and with (C and D) starter.
보다 낮은 결과를 보였다. Yoo 등(1998)은 메주의 pH가 발 효 초기의 5.5에서 발효가 진행될수록 급격히 증가하여 발 효 26일에 8.5까지 증가하였는데, 이는 메주에 미생물이 번 식하면서 대두 단백질의 분해물 및 암모니아성 질소화합물 이 다량 생성되었기 때문으로 보고하였다. 그러나 Shin 등 (2018)은 지역별 시판 전통메주 18종의 pH를 측정한 결과 5.31~8.21로 시료 간에 큰 차이를 보였으며, 이는 지역 및 산업체별 메주의 발효조건 및 환경이 다르기 때문으로 추정 하였다.
멸치메주 발효에 따른 색도 변화
메주 및 장류의 색은 관능적 기호도 및 숙성과 관련이 있 으며, 숙성 중에 생성되는 아미노산과 유리당에 의해 멜라노
이딘이 생성되어 장류의 색이 점차 진한 갈색으로 변하게 된다(Lee 등, 2002; Chang 등, 2010; Ahn과 Bog, 2007).
발효에 따른 대조구와 종균 멸치메주의 색도 변화를 측정한 결과는 Table 2와 같이 발효 초기에는 대조구와 종균 멸치 메주의 명도(L), 적색도(a), 황색도(b)가 유사한 수준을 나타 내었으나 발효 10일부터 명도와 황색도에서 메주 시료 간에 반대의 경향을 나타내었다. 대조구 멸치메주에서 명도와 황 색도는 발효초기 52.82와 17.85에서 발효 30일 후 49.72와 16.64로 발효에 따라 감소하는 경향을 나타낸 반면, 종균 멸치메주의 명도와 황색도는 발효초기 53.55와 17.93에서 발효 30일 후 55.07과 20.07로 증가하는 결과를 나타내었 다. 적색도는 대조구와 종균 멸치메주 모두 발효에 따라 증 가했으며 전체 발효 기간에 대조구 멸치메주의 적색도가 좀 더 높은 결과를 보였다. Fig. 7과 같이 발효 30일 후 대조구 와 종균 멸치메주의 외관 및 내부 단면을 관찰한 결과, 외관 의 색은 큰 차이가 없는 반면 메주 내부의 색은 대조구 멸치 메주가 종균 멸치메주에 비해 어두운 갈색을 나타내었다.
Kwon 등(2000)은 보리등겨로 제조한 메주의 색도를 분석 한 결과 발효에 따라 명도와 황색도는 지속적으로 감소하였 고, 적색도는 발효 초기 2.4에서 발효 5일에 4.9로 증가한 후 서서히 감소하여 발효 25일 후에 3.3을 나타냈다. Choi 등(2007)은 싹튼 콩으로 제조한 메주를 48시간 동안 발효하 면서 색도를 측정한 결과 명도, 적색도, 황색도가 모두 감소 하는 결과를 보였다. Shin 등(2018)은 전통메주 18종의 색
Table 3. Free amino acid contents of anchovy-meju after fer- mentation
Free amino acid Content (mg%) Control Starter Taurine
Aspartic acid Threonine Serine Glutamic acid Sarcosine
α-Aminoadipic acid Glycine
Alanine Citrulline
α-Aminobutyric acid Valine
Cystine Methionine Cystathionine Isoleucine Leucine Tyrosine Phenylalanine β-Alanine
β-Aminoisobutyric acid γ-Aminobutyric acid Ornithine
Lysine
1-Methylhistidine Histidine 3-Methylhistidine Carnosine Arginine Proline
43.52±0.02 0.67±0.01 0.00±0.00 5.45±0.04 99.94±0.07 1.76±0.02 13.75±0.20 0.00±0.00 25.63±0.01 157.70±0.09 8.72±0.01 38.63±0.11 0.00±0.00 10.24±0.03 3.90±0.03 16.20±0.04 24.86±0.02 7.34±0.02 8.14±0.01 9.85±0.01 9.82±0.05 25.22±0.02 9.17±0.01 14.82±0.02 0.00±0.00 5.57±0.06 0.00±0.00 0.00±0.00 0.00±0.00 0.00±0.00
30.60±0.04 26.79±0.04 8.09±0.06 10.97±0.03 224.02±0.07 26.06±0.04 19.02±0.06 23.89±0.02 0.00±0.00 153.49±0.02 14.82±0.01 17.96±0.04 25.76±0.02 10.61±0.01 3.62±0.02 13.64±0.04 21.23±0.05 21.85±0.09 19.63±0.02 2.92±0.01 0.00±0.00 4.31±0.02 3.07±0.01 27.72±0.03 0.00±0.00 23.98±0.02 0.00±0.00 0.00±0.00 3.01±0.02 278.61±0.03 Total 540.77±0.26 1,015.68±0.37 Values are expressed as the mean±SD (n=3).
도를 비교했을 때 명도(39.07~67.92), 적색도(3.57~8.87), 황색도(7.48~20.67)가 메주 간에 큰 차이를 나타냈고, 이러 한 차이는 메주 제조 시 원부재료의 종류 및 첨가비율, 메주 의 발효 정도에 영향을 받는 것으로 보고하였다. 본 연구에 서 제조한 대조구와 종균 멸치메주는 원료와 발효조건이 동 일함에도 B. velezensis L2의 접종 여부에 따라 색도 변화 에서 다른 경향을 나타내며 종균 멸치메주의 색이 더 밝은 결과를 보였다. Kim 등(2004)과 Cha 등(2017)의 연구에서 다른 부원료가 첨가된 된장을 제조한 후 된장의 명도와 색에 대한 기호도의 연관성을 분석했을 때, 밝은색을 띠는 된장의 기호도가 더 높은 결과를 보였다. 종균 멸치메주를 이용해 된장 등의 장류를 제조할 경우 제품의 품질 및 소비자의 기 호도에 긍정적인 영향을 줄 것으로 판단된다.
멸치메주의 유리아미노산 함량
장류의 발효 과정 중 효소작용으로 인해 단백질이 분해되 어 생성되는 아미노산은 맛을 좌우하는 중요한 성분으로 알 려져 있다. 대조구와 종균 멸치메주를 30일 동안 발효한 후
유리아미노산 함량을 측정하여 비교한 결과 대조구 멸치메 주에서 주요 아미노산은 citrulline, glutamic acid, taurine, valine 등으로 검출되었고, 종균 멸치메주에서 주요 아미노 산은 proline, glutamic acid, citrulline, taurine 등으로 검 출되었다(Table 3). 감칠맛을 나타내는 주요 성분인 gluta- mic acid는 종균 멸치메주에서 224.02 mg%로 측정되어 대조구 멸치메주의 함량인 99.94 mg%에 비해 2.2배 높은 결과를 보였다. 또한 단맛을 나타내는 proline은 종균 멸치 메주에서 278.61 mg%의 높은 함량이 검출된 반면, 대조구 멸치메주에서는 검출되지 않아 B. velezensis L2의 발효에 따른 단백분해효소 활성이 멸치메주의 유리아미노산 함량 에 큰 영향을 준 것으로 판단된다. 상대적으로 적은 함량이 기는 하나 대조구에 비해 종균 멸치메주에서 감칠맛을 나타 내는 aspartic acid와 단맛을 나타내는 glycine이 더 높은 함량을 나타내었다. 총 유리아미노산 함량은 대조구와 종균 멸치메주에서 각각 540.77 mg%와 1,015.68 mg%를 나타 내어 종균 멸치메주가 대조구에 비해 1.9배 높은 결과를 보 였다. Lee 등(2009)은 크기에 따른 메주의 유리아미노산 함량을 분석한 결과 메주의 크기와 상관없이 glutamic acid 가 가장 많은 함유량을 나타내었고, 그 외 arginine, histi- dine, leucine 등이 주요 아미노산으로 검출되었다. Kwon 등(2000)은 보리등겨를 첨가하여 제조한 메주의 유리아미 노산을 분석한 결과, 발효 25일 후에 glutamic acid의 함량 이 가장 많고 다음으로 proline, leucine, alanine 순으로 주요 아미노산이 검출되었다. Kim 등(2002)은 B. subtilis로 30일 동안 발효한 메주의 유리아미노산 분석 결과 glutamic acid의 함량이 가장 많고 다음으로 phenylalanine, alanine 이 높게 측정되었다. 메주의 유리아미노산 함량에서 공통적 으로 높은 함량을 보인 glutamic acid 외에 주요하게 검출되 는 아미노산의 종류는 메주의 원료 및 접종하는 균주에 따라 차이가 큰 것으로 생각된다. 본 연구에서 멸치를 첨가하여 B. velezensis L2로 발효한 종균 멸치메주는 감칠맛 성분인 glutamic acid와 단맛 성분인 proline의 비율이 높으므로 관능적인 품질 특성이 우수할 것으로 판단된다.
요 약
멸치메주의 발효 종균을 선정하기 위해 전통 발효식품으로 부터 분리된 protease 활성이 우수한 4종의 균을 이용하여 멸치분말이 첨가된 대두의 발효를 진행한 결과, 멸치액젓에 서 분리된 L2 균의 생육도 및 protease 활성이 가장 우수하 였다. 발효 종균으로 선정된 L2 균은 B. velezensis로 동정 되었으며, B. velezensis L2를 종균으로 한 멸치메주와 종 균이 접종되지 않은 대조구 멸치메주를 제조하고 30일 동안 발효를 진행하면서 품질 특성의 변화를 조사하였다. 총 균수 는 대조구와 종균 멸치메주 모두 발효 10일에 가장 높았으 며, 발효 30일 후에도 종균 멸치메주에서 B. velezensis L2 는 71%의 높은 점유율을 나타내었다. 멸치메주의 protease
활성에서 대조구는 발효 20일에 5,260 unit/g으로 증가한 후 발효 30일(4,880 unit/g)에 감소했지만 종균 멸치메주는 발효 20일(6,510 unit/g)과 30일(6,565 unit/g)에도 높은 활 성을 유지하였다. 아미노태 질소는 발효에 따라 증가했으며 발효 30일에 종균 멸치메주는 953.2 mg%로 대조구(711.7 mg%)에 비해 높은 함량을 나타냈다. 발효에 따라 pH는 증가 하고 산도는 감소하는 경향을 나타냈으며, 발효 30일에 대 조구에 비해 종균 멸치메주의 pH가 더 낮고 산도는 더 높은 결과를 보였다. 색도에서 적색도는 발효에 따라 증가했으나, 명도와 황색도는 발효에 따라 대조구는 감소했고 종균 멸치 메주는 증가하는 결과를 보였다. 발효 30일 후 유리아미노 산을 분석한 결과 감칠맛을 나타내는 glutamic acid는 대조 구(99.94 mg%)에 비해 종균 멸치메주(224.02 mg%)에서 더 높은 함량을 나타내었다. 본 연구 결과에 따라 B. vele- zensis L2로 발효된 멸치메주는 우수한 품질 특성을 나타내 며 전통된장의 품질향상에 도움이 될 것으로 판단된다.
감사의 글
본 연구는 농림축산식품부의 재원으로 농림식품기술기획평 가원의 미래형혁신식품기술개발사업의 지원을 받아 연구되 었음(119022-2).
REFERENCES
Ahn SC, Bog HJ. Consumption pattern and sensory evaluation of traditional doenjang and commercial doenjang. Korean J Food Cult. 2007. 22:633-644.
Bae JO, Lee KJ, Park JS, Choi DS. Preparation of sweet potato doenjang using colored sweet potato. Korean J Food Nutr.
2012. 25:529-537.
Bae MS, Ha JU, Lee SC. Quality properties of high calcium fish paste containing anchovy. Korean J Food Cook Sci. 2007.
23:561-566.
Cha SJ, Park SR, Kim DH. Quality characteristics of Doenjang prepared with sweet potato. Korean J Food Preserv. 2017.
24:221-229.
Chang M, Kim IC, Chang HC. Effect of solar salt on the quality characteristics of Doenjang. J Korean Soc Food Sci Nutr.
2010. 39:116-124.
Chang M, Moon SH, Chang HC. Isolation of Bacillus velezensis SSH100-10 with antifungal activity from Korean traditional soysauce and characterization of its antifungal compounds.
Korean J Food Preserv. 2012. 19:757-766.
Cho MJ, Shim JM, Lee JY, Lee KW, Yao Z, Liu X, et al.
Properties of meju fermented with multiple starters. Microbiol Biotechnol Lett. 2016. 44:109-116.
Choi UK, Kim MH, Lee NH, Jeong YS, Hwang YH. Changes in quality characteristics of Meju made with germinated soy- bean during fermentation. Korean J Food Sci Technol. 2007.
39:304-308.
Cui CB, Lee EY, Lee DS, Ham SS. Antimutagenic and anti- cancer effects of ethanol extracts from Korean traditional Doenjang added sea tangel. J Korean Soc Food Sci Nutr.
2002. 31:322-328.
Hwang CE, Joo OS, Lee JH, Song YH, Hwang IG, Cho KM.
Changes of physiochemical properties and biological activity during the fermentation of Doenjang with bitter melon (Mo- mordica charantia L.). Korean J Food Preserv. 2017. 24:134- 144.
Hwang JH. The fermentative characteristics of Cheonggukjang prepared by starter culture of Bacillus spp. with fibrinolytic activity. J Korean Soc Food Sci Nutr. 2010. 39:1832-1838.
Hyun KW, Lee JS, Ham JH, Choi SY. Isolation and identi- fication of microorganism with potent fibrinolytic activity from Korean traditional Doenjang. Kor J Microbiol Biotech- nol. 2005. 33:24-28.
Jo JH, Oh SW, Choi JG. Processing of fermented and powdered anchovy seasoning material. J Korean Fish Soc. 1999. 32: 725- 729.
Jung HK, Jeong YS, Youn KS, Kim DI, Hong JH. Quality char- acteristics of soybean paste (Doenjang) prepared with Bacillus subtilis DH3 expressing high protease levels, and deep-sea water. Korean J Food Preserv. 2009. 16:348-354.
Kang JR, Kim GM, Hwang CR, Cho KM, Hwang CE, Kim JH, et al. Changes in quality characteristics of soybean paste Doen- jang with addition of garlic during fermentation. Korean J Food Cook Sci. 2014. 30:435-443.
Kang KH, Park SY, Je HS, Kang YM, Seoung TJ, Yoon MJ, et al. Preparing and maintaining the quality of snacks made from rice Oryza sativa and dried anchovy Engraulis japoni- cus. Korean J Fish Aquat Sci. 2016. 49:758-765.
Kim HJ, Lee JJ, Cheigh MJ, Choi SY. Amylase, protease, perox- idase and ascorbic acid oxidase activity of Kimchi ingredients.
Korean J Food Sci Technol. 1998. 30:1333-1338.
Kim IJ, Lee JO, Park MH, Shon DH, Ha YL, Ryu CH. Prepara- tion method of meju by three step fermentation. Korean J Food Sci Technol. 2002. 34:536-539.
Kim JW, Doo HS, Kwon TH, Kim YS, Shin DH. Quality char- acteristics of doenjang meju fermented with Aspergillus spe- cies and Bacillus subtilis during fermentation. Korea J Food Preserv. 2011. 18:397-406.
Kim SJ, Moon JS, Park JW, Park IB, Kim JM, Rhim JW, et al. Quality of soybean paste (Doenjang) prepared with sweet tangle, sea mustard, and anchovy powder. J Korean Soc Food Sci Nutr. 2004. 33:875-879.
KOSTAT. Fishery production survey. Statistics Korea, Dae- jeon, Korea. 2018. p 109.
Kwak JH, Kim KBWR, Song EJ, Lee CJ, Jung JY, Choi MK, et al. Effect of salt soluble protein extracts from anchovy on quality characteristics of sausage. J Korean Soc Food Sci Nutr.
2010. 39:1839-1845.
Kwon OJ, Choi UK, Lee EJ, Cho YJ, Cha WS, Son DH, et al.
Chemical changes of Meju made with barley bran using fer- mentation. Korean J Food Sci Technol. 2000. 32:1135-1141.
Kwon SH, Shon MY. Antioxidant and anticarcinogenis effects of traditional doenjang during maturation periods. Korean J Food Preserv. 2004. 11:461-467.
Lee JG, Kwon KI, Choung MG, Kwon OJ, Choi JY, Im MH.
Quality analysis on the size and the preparation method of meju for the preparation of Korean traditional soy sauce (kanjang). J Appl Biol Chem. 2009. 52:205-211.
Lee KH, Choi HS, Hwang KA, Song J. Quality changes in doen- jang upon fermentation with two different Bacillus subtilis strains. J East Asian Soc Diet Life. 2016. 26:163-170.
Lee KS, Lee JC, Lee JK, Hwang ES, Lee SS, Oh MJ. Quality of 4-recommended soybean cultivars for meju and doenjang.
Korean J Food Preserv. 2002. 9:205-211.
Lee SY, Eom JS, Choi HS. Quality characteristics of fermented soybean products by Bacillus sp. isolated from traditional soy-
bean paste. J Korean Soc Food Sci Nutr. 2014. 43:756-762.
Lee SY, Park NY, Kim JY, Choi HS. Quality characteristics of rice-doenjang during fermentation by differently shaped meju and adding starter. Korean J Food Nutr. 2012. 25:505-512.
Meena KR, Tandon T, Sharma A, Kanwar SS. Lipopeptide anti- biotic production by Bacillus velezensis KLP2016. J Appl Parm Sci. 2018. 8:91-98.
Ministry of Agriculture, Food and Rural Affairs (MAFRA). Ko- rean Traditional Food Quality Standard. 2020. Gimcheon, Korea. No. T002.
Ministry of Food and Drug Safety (MFDS). Food code. Cheon- gju, Korea. 2020. p 5-11.
Moon S, Park Y, Kim I, Chang H. Isolation and characterization of Bacillus velezensis SS360-1 from seed soy sauce. Korean J Community Living Sci. 2018. 29:49-58.
Oh SI, Sung JM, Lee KJ. Physicochemical characteristics and antioxidative effects of barley soybean paste (Doenjang) con- taining kelp extracts. J Korean Soc Food Sci Nutr. 2014. 43:
1843-1851.
Park JH, Ha AW, Cho JS. Effects of green tea-soybean paste on weights and serum lipid profiles in rats fed high fat diet.
Korean J Food Sci Technol. 2005. 37:806-811.
Park KY, Moon SH, Cheigh HS, Baik HS. Antimutagenic effects of doenjang (Korean soy paste). Prev Nutr Food Sci. 1996.
1:151-158.
Rhee CH, Kim WC, Rhee IK, Park HD. Effects of inoculation of Bacillus subtilis cells on the fermentation of Korean tradi- tional soy paste (Doenjang). Korean J Food Preserv. 2008.
15:598-605.
Rho JD, Choi SY, Lee SJ. Quality characteristics of soybean paste (doenjang) prepared using different types of microor- ganisms and mixing ratios. Korean J Food Cook Sci. 2008.
24:243-250.
Saitou N, Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987.
4:406-425.
Shin DS, Han SI, Choi ID, Lee SK, Park JY, Kim NG, et al.
Physicochemical characteristics and microbiological distribu- tion of Korean traditional Meju of various region. Korean J Food Nutr. 2018. 31:712-719.
Yao Z, Kim JA, Kim JH. Characterization of a fibrinolytic en- zyme secreted by Bacillus velezensis BS2 isolated from sea squirt jeotgal. J Microbiol Biotechnol. 2019. 29:347-356.
Ye M, Tang X, Yang R, Zhang H, Li F, Tao F, et al. Characteris- tic and application of a novel species of Bacillus: Bacillus velezensis. ACS Chem Biol. 2018. 13:500-505.
Yoo JY, Kim HG, Kim WJ. Physico-chemical and microbio- logical changes of traditional Meju during fermentation in Kangweondo area. Korean J Food Sci Technol. 1998. 30:908- 915.
Yoo JY, Kim HG. Changes in microflora and enzyme activities of traditional Meju during fermentation at Sunchang area. J Korean Soc Food Sci Nutr. 1998. 27:448-454.
Yu R, Park SA, Chung DK, Nam HS, Shin ZI. Effect of soybean hydrolysate on hypertension in spontaneously hypertensive rats. J Korean Soc Food Sci Nutr. 1996. 25:1031-1036.