저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학 석사학위논문
진균 증식에서 5 nm 은나노입자의
억제 효과와 기전 규명
아 주 대 학 교 대 학 원
의생명과학과/분자의학전공
이 보 경
진균 증식에서 5 nm 은나노입자의
억제 효과와 기전 규명
지도교수 박 선
이 논문을 이학 석사학위 논문으로 제출함.
2018년 2월
아 주 대 학 교 대 학 원
의생명과학과/분자의학전공
이 보 경
이보경의 이학 석사학위논문을 인준함.
심 사 위 원 장 박 선 인
심 사 위 원 신 호 준 인
심 사 위 원 김 경 민 인
아 주 대 학 교 대 학 원
2018년 1월 2일
- i -
-
국문요약-
진균 증식에서 5 nm 은나노입자의 억제 효과와 기전 규명
은나노입자의 증식 억제 효과는 다양한 진균에서 입증되었으나 그 기전은 명확하게 알려지지 않았다. 본 연구에서는 polyvinylpyrrolidone으로 코팅된 5 nm 은나노입자의 항진균효과를 Candida albicans와 Saccharomyces cerevisiae에서 확 인하고 그 기전을 밝히고자 하였다. 실험결과, 5 nm 은나노입자는 두 가지 진균 에서 세포주기를 G1기에 지연시켜 증식억제 효과를 나타냈다. 또한 C. albicans 에서 은나노입자에 의해 세포사멸이 일어났으며, 포도당 소비량이 증가하였다. 활성산소종 제거제인 NAC을 은나노입자와 함께 처리하였을 때 C. albicans에서 G1기의 세포 수가 감소하였다. S. cerevisiae의 유전자 결손균주를 이용한 실험에 서 은나노입자에 의한 세포 증식 억제 효과는 야생형과 유전자 결손균주 간에 차이가 없었다. 본 연구에서는 5 nm 은나노입자의 증식 억제 효과를 두 가지 진균에서 입증하였다. 또한 5 nm 은나노입자의 항진균효과는 C. albicans에서는 활성산소종 생성과 관련이 있으며, S. cerevisiae에서는 활성산소종이나 세포벽 스 트레스 반응과 관련이 없을 가능성을 제시한다.
핵심어: 은나노입자, 증식 억제 효과, Candida albicans, Saccharomyces cerevisiae, 항진균 효과 기전.
- ii -
차 례
국문요약 ... i 차례 ... ⅱ 그림 차례 ... ⅳ I. 서론 ... 1 II. 재료 및 방법 ... 5 1) 은나노입자 및 화학약품 ... 5 2) 진균의 배양 ... 5 3) 진균 배양의 은나노입자 처리 ... 5 4) 진균 증식 분석 ... 6 5) 진균의 세포사멸 분석 ... 6 6) 진균의 세포주기 분석 ... 7 7) 진균의 포도당 소비 분석 ... 7 III. 결과 ... 8 1) C. albicans에서 5 nm 은나노입자의 항진균효과 분석 ... 8 2) S. cerevisiae에서 5 nm 은나노입자의 항진균효과 분석 ... 10 3) 은나노입자가 진균의 세포사멸에 미치는 영향 ... 12 4) 은나노입자가 진균의 세포주기에 미치는 영향 ... 14- iii - 5) 은나노입자가 항산화반응 또는 세포벽 손상반응과 관련한 유전자 결핍 S. cerevisiae의 증식에 미치는 영향 ... 16 6) 은나노입자가 항산화반응 또는 세포벽 손상반응과 관련한 유전자 결핍 S. cerevisiae의 사멸에 미치는 영향 ... 21 7) 은나노입자가 진균의 포도당 소비에 미치는 영향 ... 23 8) 활성산소종 억제제가 5 nm 은나노입자에 의해 유도된 G1기 지연에 미치는 영향 ... 25 IV. 고찰 ... 27 V. 결론 ... 31 참고문헌 ... 32 ABSTRACT ... 37
- iv -
그 림 차 례
Fig. 1. The growth of C. albicans was inhibited by treatment with 5 nm AgNPs ... 9 Fig. 2. The growth of S. cerevisiae was inhibited by treatment with 5 nm AgNPs ... 11
Fig. 3. Treatment with 5 nm AgNPs increased cell death in C. albicans but not in S. cerevisiae .. 13
Fig. 4. Treatment with 5 nm AgNPs induced G1 phase delay in C. albicans and S. cerevisiae ... 15
Fig. 5. The effect of 5 nm AgNPs on the growth of S. cerevisiae were similar in ∆SKN7, ∆YAP1, ∆HOG1 and ∆SLT2 mutants compared to wild type ... 17
Fig. 6. 5 nm AgNPs-induced cell death of S. cerevisiae were similar in ∆SKN7, ∆YAP1, ∆HOG1 and ∆SLT2 mutants compared to wild type ... 22
Fig. 7. Teatment of 5 nm AgNPs differentially affects glucose consumption in C. albicans and S.
cerevisiae ... 24
Fig. 8. ROS scavenger treatment restored cell cycle in C. albicans treated with 5 nm AgNPs but not in S. cerevisiae. ... 26
- 1 -
I.
서 론
백선 및 칸디다증과 같은 진균에 의한 감염성 질병의 종류와 발병률은 높 아지고 있지만 그 치료법은 다양하지 않다. 항생제를 이용한 기존의 치료법은 병증에 따라 효과를 보지 못하는 경우가 있으며, 진균이 항진균제(Anti fungal agent)에 내성을 나타내기도 한다. 또한 플루코나졸(Fluconazole)과 같은 항진균 제는 감염된 진균의 종류에 따라 효과가 다르다(Zeina A. Kanafani, 등 2008). 이 러한 이유로 항진균효과를 갖는 새로운 물질에 대한 연구가 활발하게 이루어지 고 있는데, 그 중 하나가 은나노입자이다.은나노입자(silver nanoparticle, AgNP)는 1 nm에서 100 nm 사이의 은입자이다. 은나노입자는 표면이 시트르산삼나트륨(trisodium citrate)이나 polyvinylpyrrolidone (PVP) 등으로 코팅되어 있으며 구형, 다이아몬드형, 팔각형, 판형 등이 있다. 항 균효과가 있다고 알려진 80 nm 미만의 은나노입자는 크기와 모양이 다양하다 (Christina 등, 2003; Morano 등, 2012).
은나노입자는 세균과 진균에서 독성이 있다고 보고되었다. 세균에서는 아 시네토박터균(Acinetobacter baumannii)(Franci 등, 2015), 살모넬라균(Salmonella
typhi)과 콜레라균(Vibrio cholerae)(Morones 등, 2005), 황색포도상구균 (Staphylococcus aureus)(Biel 등, 2011)등 인간에 감염될 수 있는 여러 종의 세균 에서 은나노입자가 살균효과를 보인다고 보고되었다. 진균에서는 Saccharomyces
cerevisiae와 같은 효모와 칸디다증(Candidasis)의 원인이 되는 Candida 종 등에서
은나노입자의 항진균효과가 입증되었다(Paredes 등, 2014). S. cerevisiae 효모에서 는 85 nm의 은나노입자를 48 시간 처리하였을 때 생장 억제 효과가 관찰되었고, 병원성 진균인 Candida albicans는 15.5 nm의 은나노입자를 5 ug/ml로 10 시간 처 리하였을 때 생장 억제 효과를 보였다. 또한 C. albicans와 Aspergillus flavus에서 은나노입자를 단독으로 처리하였을 때 보다 thiazolidine, hydrazide와 같은 화합
- 2 -
물과 함께 처리하였을 때 항진균효과가 더 증가되었다(Jain 등, 2011). 각막염 (keratitis) 환자에서 분리한 Fusarium, Alternaria등 216개 진균에 은나노입자를처 리하였을 때 netamycin 항생제보다 살균효과가 큰 것이 입증되었다(Xu 등, 2013).
은나노입자의 효과에 영향을 미치는 요인으로는 입자의 크기, 모양, 세포의 농도 등이 있다. 황색포도상구균(Staphylococcus aureus)과 폐렴간균(Klebsiella
pneumoniae)을 이용한 연구에서 80 nm 이상의 은나노입자 보다 30 nm 이하의
작은 은나노입자의 살균효과가 더 크다고 밝혔다(Collins 등, 2010). 은나노입자 는 세균의 표면에 접착하여 축적되면 세포막 구조를 변형시키고 투과성을 높여, 세포를 사멸시킨다(Dutta 등, 2011). 이를 바탕으로 대장균(Escherichia coli)을 이 용한 연구에서 세포막에 더 많이 축적될 수 있는 구형의 은나노입자가 세포에 더 큰 손상을 줄 가능성을 제시하였다(Ivask 등, 2014). 황색포도상구균의 일종인
methicillin-resistant Staphylococcus aureus(MRSA)을 이용한 연구에서는 0.5, 1, 2
ug/ml 농도의 은나노입자를 OD 0.1의 생장기 세포와 0.6의 생장기 세포에 처리 하였을 때 초기 세포 농도가 낮았던 실험군에서 생장 억제 효과가 더 높게 관 찰되었다고 보고하였다(Kalhapure 등, 2015).
은나노입자의 세포 독성 기전으로 생각되는 것은 크게 두 가지가 있다. 첫 번째는 은나노입자가 세포 내에서 활성산소종(reactive oxidative species)의 생산을 유도하여 세포 독성을 일으킨다는 것이다. 2014년에 발표된 논문에 따르면 S.
cerevisiae에 은나노입자를 처리하였을 때 세포의 활성산소종 생산량이 증가되었
다고 한다(Gomez 등, 2014). 두 번째는 은나노입자가 세포벽에 스트레스를 유도 하여 세포에 독성을 일으킨다는 것이다. 2015년 보고에서는 투과전자현미경 (transmission electron microscope, TEM)을 이용하여 S. cerevisiae에 시트르산으로 코팅된 은나노입자를 처리한 경우 세포벽이 손상됨을 관찰하였다(Loredana등, 2015). 그러나 다른 연구에서 S. cerevisiae 산화스트레스 반응에 관여하는 SOD1 과 2, YAP1 유전자와 세포벽의 integrity에 관여하는 mnn10, knr4, kre6와 내포작 용에 관여하는 end3등 9개 유전자의 결손 균주에 은나노입자를 처리한 후 활성
- 3 - 산소종 생성량, 세포 생존율, 독성효과 등을 비교한 결과 유일하게 end3가 야생 형과 차이를 보였다(Käosaar 등, 2016). 이것은 선행 연구와는 달리 은나노입자 가 세포의 활성산소종 생산이나 세포 벽 스트레스 반응과 연관이 적을 가능성 을 시사하는 한편 은나노입자의 작용 기작이 내포작용과 연관되어 있을 가능성 을 보여주었다. 또한 최근에는 은나노입자가 인간의 조골세포(osteoblast)에서 세 포 주기의 지연을 유발하여 세포자멸사(apoptosis)가 일어났다는 결과(Rosário 등, 2016)를 통해 은나노입자가 세포 내 다양한 대사 과정과 관련이 있을 가능성을 제시하였다. 이처럼 은나노입자의 세포 독성 기전은 아직 명확하게 밝혀지지 않았다. 그동안 은나노입자에 대한 연구는 진균보다는 세균에서 더 활발하게 이루 어졌다. 2014년 이전에 발표된 논문의 수를 살펴보면 세균에서는 590여 편이 발 표된 반면에, 진균에서는 70여 편으로 매우 적다(Franci 등, 2015). 은나노입자를 진균에 의한 감염성 질병의 치료제로 개발하기 위해서는 진균에서 은나노입자 의 작용 기전 연구가 더 필요하다고 생각된다. S. cerevisiae는 각종 분자생물학적 연구에 널리 이용된다. 이는 S. cerevisiae
의 효모게놈프로젝트(yeast genome project)를 통해 16개 염색체, 6275개의 유전자 가 밝혀졌고(Botstein 등, 1997), 유전자 대부분의 결손 균주가 이미 제작되어 있 기 때문으로 생각된다. 또한 S. cerevisiae는 비병원성 진균으로 실험이 용이하다 는 장점이 있기 때문에 S. cerevisiae의 유전자 결손균주는 은나노입자의 항진균 효과 기전에 대한 연구에 이용하기 적합할 것이다. 활성산소종과 세포벽 스트 레스 반응에 관여하는 유전자의 결손 균주를 이용한 보고는 한편 발표되어 있 다. 스트레스 반응에 관여하는 유전자 YAP1의 변이체를 이용한 연구의 경우 YAP1의 결손을 보상할 수 있는 SKN7의 동시 결손 변이체를 이용한 실험이 수 행되지 않았다. 이에 본 연구에서는 기존 보고와 차별화된 유전자의 결손 균주 를 이용하여 실험을 수행하였다. 항산화반응 유전자의 전사인자인 YAP1은 세 포질에 존재하고 있다가 활성산소종이 생산되면 핵으로 이동하여 타겟 유전자
- 4 -
인 TRX2(thioredoxin encoding gene) 발현을 유도함으로써 항산화 반응을 촉진시 킨다. TRX2 유전자는 또 다른 전사인자인 SKN7에 의해서도 조절되는데 SKN7 과 YAP1은 각각 독립적으로, 또는 상호 보완적으로 항산화반응에 관련된 유전 자의 전사를 조절하기도 한다(Mulfold 등, 2011). 본 연구에서는 이 두 유전자 각각의 결손 진균과 두 유전자 모두의 결손 균주를 이용하여 은나노 입자의 항 진균효과를 평가하고자 하였다. 또한 세포벽 스트레스 반응 관련 유전자인 HOG1과 SLT2는 mitogen activated protein kinase(MAPK) 신호전달을 매개하여 고 삼투성 환경과, 세포벽의 스트레스에 대응 반응을 유도하고 이 중 HOG1은 고 삼투성 환경 뿐 만 아니라 항산화반응에도 관여한다고 알려져 있다(Zhao 등, 2007). 이 두 가지 유전자가 결손된 진균에 대한 은나노입자의 증식 억제 효과 는 아직 보고되지 않았다. 본 연구의 목적은 병원성 및 비병원성 진균에서 5 nm 은나노입자의 항진균 효과를 확인하고 그 기전을 밝히는 것이다. 이를 위해 C. albicans와 S. cerevisiae 에 polyvinylpyrrolidone로 코팅된 5 nm 은나노입자를 처리하여, 세포의 생장과 사멸을 분석하여 은나노입자의 항진균효과를 관찰하고 S. cerevisiae의 야생형 (BY4741)과 SKN7, YAP1, HOG1, SLT2 유전자의 단독 혹은 복합 결손 균주를 이 용하여 진균에서 은나노입자의 항진균효과 기전을 탐색하고자 하였다.
- 5 -
II.
재료 및 방법
1) 은나노입자 및 화학약품
본 연구에서는 polyvinylpyrrolidone으로 코팅된 5 및 100 nm 크기의 은나 노입자(I&C Technology, Seoul, 한국)를 이용하였다. Difco Sabouraud Dextrose(SD) Broth와 yeast extract, peptone, dextrose(YPD)를 BD사(Miami, 미국)에서 구입하였다. Propidium iodide는 Millipore사(Billerica, MA, 미국)에서 구매하였고, RNAse A는 Thermofisher사(Waltham, MA, 미국)에서 구매하였다. N-Acetyl-L-cystein은 SIGMA 사(St. Louis, MO, 미국)에서 구매하였다.
2) 진균의 배양
C. albicans는 SD 한천배지에 선조접종(streaking)하여 30℃에서 1 일 배
양한 후 단일집락을 취해 SD 액체배지에 접종하여 30℃ 140 rpm으로 배양하였 다. S. cerevisiae의 야생형(BY4741)과 ∆SKN7, ∆YAP1, ∆SKN7&YAP1, ∆HOG1, ∆ SLT2, ∆HOG1&SLT2의 7개 균주를 YPD 한천배지에 선조접종하여 30℃에서 2 일 배양하였다. 이후 단일집락을 취해 YPD 액체 배지에 접종하여 30℃에서 140 rpm으로 배양하면서 흡광도를 측정하여(BECKMAN) OD 600 nm 0.6 ~1.0의 배 양액으로 15% glycerol stock 1 ml을 만들어 –70℃에 보관하였다.
3) 진균 배양의 은나노입자 처리
냉동 보관한 C. albicans 1 ml을 SD 액체배지 30 ml에 녹여 37℃에서 140 rpm으로 OD 0.8 ~1.0의 생장기가 되도록 배양하였다. 이를 OD 600 nm 0.1 이 되- 6 -
도록 SD 액체배지로 희석한 다음 24 well plate에 넣고 100 및 5 nm의 은나노입 자 각각을 2, 10, 20, 50 ug/ml 농도로 처리한 후 37℃에서 24 시간 배양하였다.
냉동 보관한 S. cerevisiae 7개 균주 1 ml을 YPD 액체배지 30 ml에 녹여 OD 0.3 ~0.5 의 생장기가 되도록 배양하였다. 이를 OD 600 nm 0.1 이 되도록 YPD 액체배지로 희석한 다음 24 well plate에 넣고 100 및 5 nm의 은나노입자 각각을 2, 10, 20, 50 ug/ml 농도로 처리한 후 30℃에서 28 시간 배양하였다.
4) 진균의 증식 분석
3항의 기술과 같이 배양하면서 C. albicans의 경우 2 시간 간격으로 S. cerevisiae의 경우 4 시간 간격으로 600 nm 파장에서 OD값을 측정하여 진균의 증식을 측정하였다. 또한 C. albicans의 경우 배양 24 시간에 10 배씩 계단 희석 한 배양액 100 ul를 SD 한천배지에 도말한 후 37℃에서 1일 배양하고 생성된 단일집락 수를 측정하여 증식을 분석하였다. S. cerevisiae의 경우 spot 검사로 증 식을 분석하였는데 간략히 기술하면, 배양 28 시간에 10 배씩 계단 희석한 배 양액 5 ul를 YPD 한천배지에 떨어뜨린 후 3일 배양하여 증식을 분석하였다.5) 진균의 세포사멸 분석
600 nm OD 0.1의 C. albicans와 S. cerevisiae에 100 및 5 nm 은나노입자 각 각을 2, 20, 50 ug/ml로 처리하고 C. albicans는 37℃에서, S. cerevisiae는 30℃에서 배양하였다. 6 시간과 24 시간 배양한 다음 세포수 약 10⁶ /ml (OD 600 nm 0.1)이 되도록 액체배지로 희석하고 상온에서 3000 xg, 5 분 원심 분리하여 상청액을 제거하였다. 인산완충식염수(PBS) 1 ml로 세포를 2 회 세척한 후에 상청액을 모 두 제거하고 200 ul PBS로 세포를 재부유 한다음 Propidium iodide(Millipore사)을 250 ng/ml이 되도록 첨가하였다. 상온에서 30 분간 정치한 후 유세포 분석기- 7 -
(flow cytometry)(BD facs Canto™Ⅱ)로 분석하였다.
6) 진균의 세포주기 분석
600 nm OD 0.1의 C. albicans와 S. cerevisiae의 야생형 균주 BY4741 배양액 4 ml을 round bottom tube에 넣고 100 및 5 nm의 은나노입자 각각을 2, 20, 50 ug/ml 농도로 단독 처리하거나 5 mM의 NAC을 함께 처리하였다. C. albicans는 37℃에 서 S. cerevisiae는 30℃에서 140 rpm으로 배양하면서 1 시간, 1 시간 30 분, 2 시 간, 4 시간에 1 ml의 배양액을 수거하여 다음과 같이 세포주기를 분석하였다. 배 양액을 상온에서 3000 xg, 5 분간 원심 분리하여 상청액을 제거하고 PBS를 넣어 침전된 세포를 두 번 세척하였다. 세포를 PBS 200 ul로 재부유하고 sonication 10 초 및 pipetting를 40회하여 단일 세포로 분리하였다. RNAse A를 100 ug/ml이 되 도록 첨가하여 37℃에서 3 시간 정치한 후 PBS로 두 번 세척하여 PBS 200 ul으 로 세포를 재부유하였다. 여기에 propidium iodide를 10 ug/ml이 되도록 첨가하고 상온에서 4 시간 정치한 후 유세포 분석기로 분석하였다.
7) 진균의 포도당 소비 분석
600 nm OD 0.1의 C. albicans와 S. cerevisiae의 야생형에 6항에 기술한 것 과 같이 은나노입자를 처리한 후 C. albicans는 37℃에서 S. cerevisiae는 30℃에 서 140 rpm으로 배양하였다. 배양 시작 후 1 시간, 2 시간, 4 시간에 배양액 1 ml 을 수거하여 4℃에서 13000 rpm으로 5 분간 원심 분리 하였다. 상청액 900 ul를 수집하여 YSI (7100MBS) 기기로 포도당 소비량을 측정하였다.- 8 -
III. 결 과
1) C. albicans에서 5 nm 은나노입자의 항진균효과 분석
본 연구에서는 먼저 병원성 진균에서 5 nm 은나노입자의 항진균효과를분석 하고자 하였다. 생장기의 C. albicans에 2, 10, 20, 50 ug/ml의 5 nm 은나노입자와 비교군으로 100 nm 은나노입자를 처리한 후 24 시간 배양하였다. 은나노입자를 처리하지 않은 대조군의 C. albicans 배양의 경우 2 시간마다 측정한 흡광도가 14 시간까지 증가하다가 이후 증가하지 않았다. 각 농도의 100 nm 은나노입자 를 처리한 경우와 2 ug/ml 농도의 5 nm 은나노입자를 처리한 경우 대조군과 비 슷한 흡광도 변화를 보였다. 그러나 10 ug/ml 또는 20 ug/ml의 5 nm 은나노입자 를 처리한 경우 흡광도가 12 시간까지 증가 후 변하지 않았으며, 대조군의 최 고 흡광도의 약 50%를 넘지 못하였다. 또한 50 ug/ml의 5 nm 은나노입자를 처 리한 경우 흡광도의 증가를 관찰하지 못하였다(Fig. 1A). 그러므로 5 nm 은나노 입자는 10 ug/ml 이상의 농도에서 증식억제효과를 보였다. 흡광도 측정에 의한 균 성장속도 결과를 확인코자 각 조건에서 24 시간 배양한 진균을 계단 희석하 여 SD 한천 배지에 도말한 후 CFU를 측정하였다. 대조군과 100 nm 은나노입자 를 처리한 군의 CFU는 10⁹ CFU/ml로 비슷하였으며 2 ug/ml의 5 nm 은나노입자 를 처리한 경우 비슷한 CFU를 보였다. 10 또는 20 ug/ml의 5 nm 은나노입자를 처리한 경우에서 대조군에 비해 각각 약 10 배, 100 배 감소한 CFU/ml이 측정 되었으며 이는 통계적으로 유의한 차이를 보였다. 또한 50 ug/ml의 농도에서 진 균의 증식이 관찰되지 않았다(Fig 1B, C). 그러므로 흡광도 측정 결과와 유사하 게 10 ug/ml 이상의 5 nm 은나노입자의 C. albicans 증식억제효과를 관찰하였다.- 9 -
(A)
(B) (C)
Fig. 1. The growth of Candida albicans was inhibited by treatment with 5 nm AgNPs. C. albicans were treated with the indicated concentrations of 5 or 100 nm AgNPs for 24 hour. Cultures were incubated at 37℃ with shaking. (A) Cell growth was measured using spectrophotometer at every 4 hour. (B),(C) Cultures of 24 hour were incubated on SD agar plate to measure CFU.(*** p<0.0001)
- 10 -
2) S. cerevisiae에서 5 nm 은나노입자의 항진균효과 분석
다음으로 비병원 진균 S. cerevisiae에서 5 nm 은나노입자의 증식 억제 효과 를 분석하였다. 은나노입자를 처리하지 않은 대조군의 흡광도는 20 시간까지 증가하다가 이후 증가하지 않았다. 각 농도의 100 nm 은나노입자를 처리한 군 의 흡광도는 대조군과 비슷한 변화를 보였다. 반면에 2 ug/ml의 5 nm 은나노입 자를 처리한 경우 20 시간 이후의 흡광도가 대조군에 비해 낮게 측정되었다. 또한 10, 20, 50 ug/ml의 5 nm 은나노입자를 처리한 경우에는 12 시간까지 흡광 도가 증가하다가 이후 증가하지 않았으며 대조군의 최고 흡광도의 약 50%를 넘지 못하였다(Fig. 2A). 흡광도 측정에 의한 진균 증식 결과를 확인코자 각 조 건에서 28 시간 배양한 진균을 계단 희석하여 YPD 한천 배지에 spot test를 수 행하였다. spot test 결과에서도 동일하게 대조군에 비해 5 nm 은나노입자는 S.- 11 -
(A) Wild type (B)
Fig. 2. The growth of Saccharomyces cerevisiae was inhibited by treatment with 5 nm AgNPs. S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs for 28 hour. Cultures were incubated at 30℃ with shaking. (A) Cell growth was measured using spectrophotometer at every 4 hour. (B) Cultures(5 ul) of 28 hour were spotted on SD agar plate and then incubated for 3 days at 30℃.
- 12 -
3) 은나노입자가 진균의 세포사멸에 미치는 영향
5 nm 은나노입자의 증식억제효과가 세포사멸에 의한 것인지 알아보고자 다 음의 실험을 수행하였다. C. albicans와 S. cerevisiae에, 은나노입자를 처리한 후 6 시간과 24 시간에 세포사멸을 유세포 분석기로 분석하였다. 아무것도 처리하지 않은 C. albicans와 100 nm 은나노입자를 처리한 경우 세포 사멸 빈도가 1% 미 만이었다. 5 nm의 은나노입자를 처리한 6 시간의 세포사멸 빈도가 10, 20, 50 ug/ml의 농도에서 각각 약 30%, 40%, 70% 로 관찰되었고, 24시간에 약 50%, 80%, 90%로 관찰되었다 (Fig. 3A). 한편, S. cerevisiae는 100 nm 은나노입자를 처리한 경우 6 시간과 24시간에 1% 미만의 세포 사멸이 관찰되었고, 5 nm 은나노입자 를 처리하였을 때 2~4 %의 세포 사멸이 관찰되었다 (Fig. 3B). 이 결과는 5 nm 은나노입자가 C. albicans의 세포사멸을 유도하지만 S. cerevisiae에서는 세포사멸 을 유도하지 않음을 보이며 이는 5 nm 은나노입자의 증식 억제 효과가 S.- 13 -
(A
)
Histogram at 6 hr (B) C. albicans(C) (D)
S. cerevisiae
Fig. 3. Treatment with 5 nm AgNPs increased cell death in C. albicans but not in S. cerevisiae. C. albicans and S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs for 6 and 24 hour. Cell death was analyzed by labeling with propidium iodide (250 ng/ml) and flow cytometry. (A),(C) Representative histograms (B),(D) Data from two independent experiment with triplicate.
- 14 -
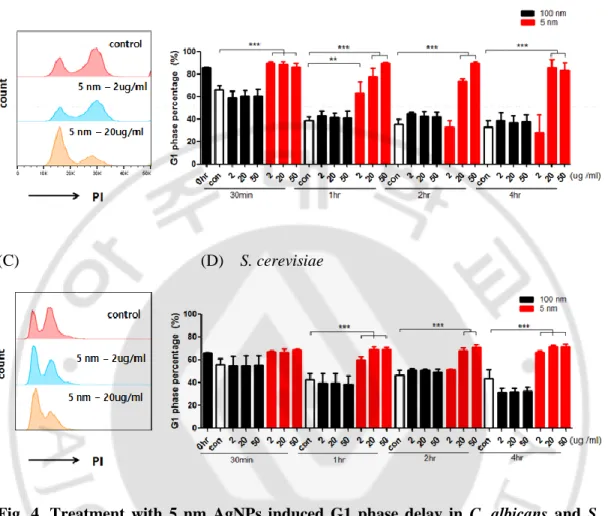
4) 은나노입자가 진균의 세포주기에 미치는 영향
다음으로는 은나노입자가 세포 주기에 영향을 미쳐 세포의 증식을 억제하 는지 알아보고자 하였다. C. albicans와 S. cerevisiae에 은나노입자를 처리한 후 30 분, 1, 2, 4 시간에 G1기 세포 비율을 유세포 분석기로 분석하였다. C. albicans 의 경우 은나노입자를 처리하지 않은 대조군과 100 nm 은나노입자를 처리한 경 우 G1기 세포 비율이 배양 시작 시점 약 90% 내외에 비해 배양 30 분에 약 60%, 배양 1 시간에 약 40%로 감소하였으며 4 시간까지 약 40%가 유지되었다. 그러나 5 nm 은나노입자를 처리한 경우 배양 30 분에 G1기 세포 비율이 약 90 %로 대조군에 비해 높은 수준이며, 2 ug/ml의 농도에서는 배양 2 시간에 40%로 감소하여 대조군과 차이를 보이지 않았으나 20 ug/ml과 50 ug/ml에서는 분석한 모든 시점에서 G1기 세포 비율이 대조군에 비해 높은 수준을 유지하였다(Fig. 4A, B). 이러한 결과는 5 nm 은나노입자가 세포주기의 정상적인 진행을 방해함 으로써 증식 억제 효과를 나타낼 것을 보여준다. 그러므로 5 nm 은나노입자는 농도에 의존적으로 C. albicans의 세포주기에 영향을 미쳐 G1기의 지연을 일으 키는 것을 알 수 있었다. 유사하게 S. cerevisiae에서 2 ug/ml의 5 nm 은나노입자 의 처리는 일시적으로 G1기 지연을 유도하고 20 ug/ml과 50 ug/ml의 5 nm 은나 노입자의 처리는 4 시간동안 지속되는 G1기 지연을 유도함을 관찰하였다(Fig. 4C, D).- 15 -
(A) Histogram at 2 hr (B) C. albicans
(C) (D) S. cerevisiae
Fig. 4. Treatment with 5 nm AgNPs induced G1 phase delay in C. albicans and S. cerevisiae. C. albicans and S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs at the indicated time point. The cells were labeled with propidium iodide and the cell cycle was analyzed using flow cytometry. (A),(C) Representative histograms (B),(D) Data from two independent experiment with triplicate.(** p<0.005, *** p<0.0001)
- 16 -
5) 은나노입자가 항산화반응 또는 세포벽 손상반응과 관련한 유전자
결핍 S. cerevisiae의 증식에 미치는 영향
은나노입자의 진균 증식 억제 기전이 활성산소종 생산 또는 세포벽 손상과 관련이 있는지 알아보고자 S. cerevisiae의 4 가지 유전자 결핍균주(SKN7, YAP1, HOG1, SLT2)를 이용하여 실험하였다. SKN7과 YAP1는 전사인자로 세포에 활성 산소종이 생산되었을 때 항산화 반응을 촉진하는 유전자인 TRX2의 발현을 촉 진한다. HOG1과 SLT2는 MAPK 신호 전달을 매개하여 고삼투성, 세포벽 스트레 스에 대한 보호 반응을 촉진하는 유전자이다. S. cerevisiae ∆SKN7, ∆YAP1, ∆ SKN7과 ∆YAP1, ∆HOG1, ∆SLT2, ∆HOG1과 ∆SLT2 균주에 은나노입자를 처리 한 후 28 시간까지 배양하면서 흡광도를 측정하였다. 야생형과 마찬가지로 각 농도의 100 nm 은나노입자를 처리한 경우 S. cerevisiae 유전자 결손 균주의 증식 을 억제시키지 못하였다. 5 nm 은나노입자는 2 ug/ml부터 농도에 비례하여 증식 을 억제 하였으나, 그 효과는 야생형과 비교하여 유전자 결손 균주 사이에 통 계적으로 유의적인 차이를 보이지 않았다(Fig 5A~F). S. cerevisiae 야생형을 포함 한 7개 균주의 증식 그래프를 이용하여 생장 억제 비율을 계산하였다. 배양 8 시간에 5 nm 은나노입자의 50% 증식억제농도(Inhibitory concentration, IC50)는 야 생형과 유전자 결손 균주에서 약 20 ug/ml로 차이가 없었으며, 24 시간에도 그 농도가 약 10 ug/ml 이하로 차이가 없었다(Fig. 5G). 다음으로 S. cerevisiae ∆ SKN7, ∆YAP1, ∆SKN7과 ∆YAP1, ∆HOG1, ∆SLT2, ∆HOG1과 ∆SLT2 균주에 은 나노입자를 처리하여 28 시간 배양한 후 spot test를 통해 선행 실험 결과를 한 번 더 확인하였다. spot test 결과 모든 유전자 결손 균주에서 5 nm 은나노입자에 의한 증식 억제효과가 야생형에서의 효과와 차이가 없었다(Fig. 5H~M).- 17 -
(A) (B)
(C) (D)
- 18 -
(G)
- 19 -
(J) ∆SKN7&YAP1 (K) ∆HOG1
- 20 -
Fig. 5. The effect of 5 nm AgNPs on the growth of S. cerevisiae were similar in ∆ SKN7, ∆YAP1, ∆HOG1 and ∆SLT2 mutants compared to wild type. Deletional mutans of S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs for 28 hour. (A~F) Cell growth was measured using spectrophotometer at every 4 hour. (G) Growth inhibition at 8 and 20 hour was calculated by {1-[(O.D of each strain treated with AgNPs at the indicated time point)-(O.D of each strain at 0 time point)]/[(O.D of untreated strain at the indicated time point)-(O.D of untreated strain at 0 time point)]}*100 (H)~(M) At 28 hour, serial dilution of each culture was spotted on agar plate and then cultured for 3 days.
- 21 -
6) 은나노입자가 항산화반응 또는 세포벽 손상반응과 관련한 유전자
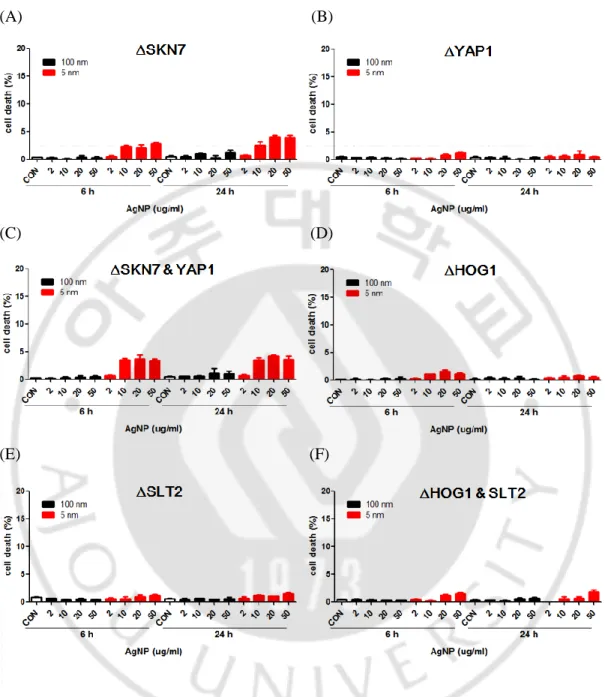
결핍 S. cerevisiae의 사멸에 미치는 영향
다음으로 S. cerevisiae ∆SKN7, ∆YAP1, ∆SKN7과 ∆YAP1, ∆HOG1, ∆SLT2, ∆ HOG1과 ∆SLT2 균주에 은나노입자를 처리한 후 6 시간과 24 시간에 세포사멸 을 유세포 분석기로 분석하였다. 야생형과 비교하여 S. cerevisiae ∆SKN7, ∆ SKN7과 ∆YAP1 균주에서 세포사멸 비율이 약간 높았으나 모든 경우의 세포 사멸 비율이 4 % 이하로 낮게 관찰되었다(Fig. 6A~F). 이 결과는 S. cerevisiae에 서 은나노입자의 증식 억제 기전이 활성산소종 생산에 대한 방어 기작 또는 세 포벽 스트레스 반응과 관련이 적음을 시사하였다.
- 22 -
(A) (B)
(C) (D)
(E) (F)
Fig. 6. 5 nm AgNPs-induced cell death of S. cerevisiae were similar in ∆SKN7, ∆ YAP1, ∆HOG1 and ∆SLT2 mutants compared to wild type. Deletional mutants of S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs for 6 and 24 hour. Cell death was analyzed by labeling with propidium iodide(250 ng/ml) and flow cytometry
- 23 -
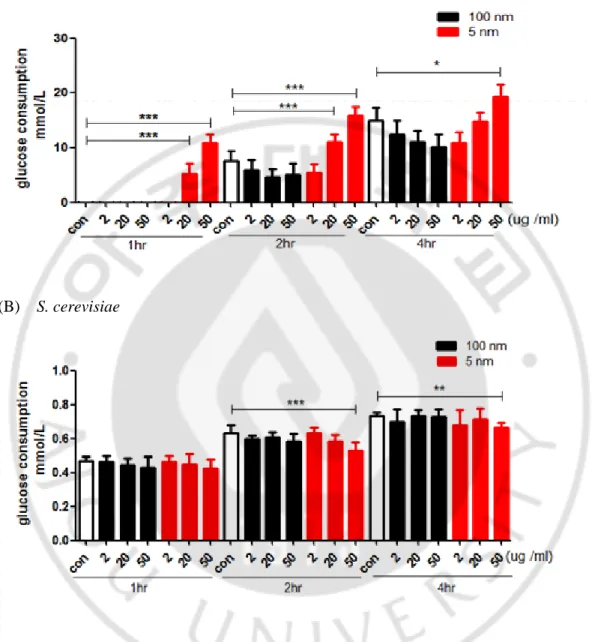
7) 은나노입자가 진균의 포도당 소비에 미치는 영향
은나노입자가 사람의 간암 세포주인 HepG2와 Huh7에서 포도당 대사에 변 화를 일으켰다는 보고(Mi Jin 등, 2015)를 바탕으로 진균에서 은나노입자가 포도 당 소비를 변화시켜 세포주기에 영향을 미치는지 알아보고자 하였다. C. albicans 와 S. cerevisiae에 은나노입자를 처리한 후 1, 2, 4 시간에 세포의 배양액에서 포 도당 소비량을 측정하였다. C. albicans에서는 1 시간에 대조군과 각 농도의 100 nm 은나노입자와 2 ug/ml의 5 nm 은나노입자를 처리한 경우에 포도당이 거의 소비되지 않았다. 그러나 대조군에 비해 포도당 소비의 증가는 20 ug/ml의 5 nm 은나노입자를 처리한 경우 2 시간까지 지속되었으며 50 ug/ml의 5 nm 은나노입 자를 처리한 경우 4 시간까지 지속되었다(Fig. 7A). 한편 S. cerevisiae에서는 배양 1 시간에 모든 군에서 포도당 소비량이 비슷하게 나타났고, 2 시간과 4 시간에 는 대조군과 각 농도의 100 nm, 2, 20 ug/ml의 5 nm 은나노입자를 처리한 경우 포도당 소비량은 비슷했지만 50 ug/ml의 5 nm 은나노입자를 처리한 군에서는 통계적으로 의미있게 포도당 소비량이 감소하였다(Fig. 7B). 이 실험결과 5 nm 은나노입자는 C. albicans와 S. cereivisiae에서 포도당의 소비에 영향을 미칠 수 있으나 두 가지 진균에서 서로 다르게 영향을 미치며, 따라서 5 nm 은나노입자 에 의한 포도당 대사 변화가 세포주기 조절 기전으로 작용할 가능성이 낮음을 시사하였다.- 24 -
(A) C. albicans
(B) S. cerevisiae
Fig. 7. Teatment of 5 nm AgNPs differentially affects glucose consumption in
C. albicans and S. cerevisiae. C. albicans and S. cerevisiae were treated with the indicated concentrations of 5 or 100 nm AgNPs. Glucose consumption was measured using a YSI biochemical analyzer at the indicated time points.(* p<0.05, ** p<0.005, *** p<0.0001)
- 25 -
8) 활성산소종 억제제가 5 nm 은나노입자에 의해 유도된 G1기 지
연에 미치는 영향
다음으로 진균에서 5 nm 은나노입자에 의해 유도된 G1기 지연 현상에 활 성산소종 제거제인 N-acetyl-L-cysteine(NAC)이 어떠한 영향을 미치는지 알아보 고자 실험을 수행하였다. C. albicans와 S. cerevisiae에 은나노입자와 함께 5mM의 NAC을 처리한 후 1 시간, 1 시간 30 분, 2 시간, 4 시간에 G1기 세포의 비율을 유세포 분석기로 분석하였다. 실험결과 C. albicans에서는 아무 것도 처리하지 않은 대조군과 NAC을 단독으로 처리한 경우 모든 분석시점에 G1기 세포의 비 율은 약 30~40%로 비슷하게 관찰되었으므로 NAC 단독 처리는 C. albicans의 세 포주기에 영향을 미치지 않음을 보였다. 그러나 1시간에 2 ug/ml의 5 nm 은나노 입자를 단독으로 처리한 군의 G1기 세포 비율이 70%였으나 NAC을 함께 처리 한 경우 40%로 감소하였으며, 4 시간까지 이 비율이 유지되었다. 또한 5 nm 은 나노입자를 20 ug/ml 또는 50 ug/ml로 단독 처리한 경우 G1기 세포의 비율이 80 %로 관찰되었는데 NAC을 함께 처리하면 그 비율이 시간이 지날수록 감소하여 G1기 지연현상이 사라지는 것을 관찰하였다(Fig. 8A). 이는 은나노입자가 C. albicans의 세포주기에 미치는 영향이 NAC에 의해 감소함을 보여주며 이는 C. albicans에서 은나노입자에 의한 G1 지연 현상에 활성산소종 생성이 관련되어 있을 가능성을 제시한다. 반면 S. cerevisiae에서는 5 nm 은나노입자를 단독으로 처리한 경우에 비해 5 nm 은나노입자와 NAC을 함께 처리한 경우에도 G1기 세 포 비율에 변화가 관찰되지 않았다(Fig. 8B). 이러한 결과는 5 nm 은나노 입자에 의한 S. cerevisiae 세포주기 지연에 활성산소종이 관여할 가능성이 낮음을 시사 한다.- 26 -
(A) C. albicans
(B) S. cerevisiae
Fig. 8. ROS scavenger treatment restored cell cycle in C. albicans treated with 5 nm AgNPs but not in S. cerevisiae. C. albicans(A) and S. cerevisiae(B) were treated with the indicated concentrations of 5 or 100 nm AgNPs with or without NAC(5mM) for 1, 2, 4 hour. The cells were labeled with propidium iodide and the cell cycle was analyzed using flow cytometry.(* p<0.05, ** p<0.005)
- 27 -
IV.
고 찰
본 연구에서는 polyvinylpyrrolidone(PVP)으로 코팅된 5 nm 은나노입자의 항 진균효과를 두 가지 진균 세포에서 확인하였다. 병원성 진균인 Candida albicans 와 Saccharomyces cerevisiae에서 5 nm 은나노입자를 10 ug/ml이상으로 처리한 경 우 증식이 억제되었다. 또한 두 진균에서 공통적으로 은나노입자에 의한 G1 지 연 현상을 보였으나, C. albicans에서만 활성산소종 제거제인 NAC이 은나노입자 에 의해 유도된 G1 지연 현상을 감소시켰다. 5 nm 은나노입자는 C. albicans에서 세포사멸을 유도하였지만 S. cerevisiae에서는 세포사멸을 유도하지 않았다. 또한 5 nm 은나노입자는 두 진균에서 포도당 소비를 서로 다르게 변화시켰다. S. cerevisiae의 세포 스트레스 반응과 연관된 6개의 유전자 결손 균주에서 은나노 입자에 의한 증식 억제와 세포사멸은 야생형과 차이가 없었다. C. albicans에서 다양한 종류의 은나노입자의 항진균효과가 이미 보고 되어 있다. 15.5 nm의 구형 은나노입자는 5 ug/ml로 10 시간 처리하였을 때 생장 억제 효과를 보였으며, 느타리버섯의 한 종류인 sajor-caju에서 합성된 11.85 nm의 은 나노입자를 48 시간 처리하였을 때 최소 저지 농도(minimum inhibitory concentration, MIC)는 250 ug/ml, 최소 살균 농도(minimum fungicidal concentration, MFC)는 500 ug/ml로 매우 높은 농도에서 효과를 보였다(Musa 등, 2017). 그러나 92.3 nm의 구형 은나노입자는 항진균효과를 보이지 않았다(Xue 등, 2016). 본 연 구에서도 100 nm 은나노입자는 C. albicans에서 항진균효과를 보이지 않았다. 이 는 80 nm 이상의 은나노입자는 항진균효과를 보이지 않는다는 기존의 보고와 동일한 결과이다. 본 연구에서 5 nm 은나노입자는 2 ug/ml의 낮은 농도에서는 효과가 없었으나, 10 ug/ml 이상의 농도에서 2 시간째부터 증식 억제 효과가 나 타났다. 또한 5 nm 은나노입자는 10 ug/ml 이상의 농도에서 6 시간째부터 세포 사멸을 일으켰다. 이 결과는 기존에 C. albicans에서 보고된 바 없는 5 nm 은나
- 28 -
노입자의 항진균효과에 대한 자료를 제시하였다.
S. cerevisiae의 야생형(BY4741)에 은나노입자를 처리하였을 때, Candida albicans에서와 마찬가지로 10 ug/ml 이상의 농도에서 생장 억제가 관찰되었다. 그러나 S. cerevisiae에서는 5 nm 은나노 입자를 처리한 후 24 시간까지 세포사멸 은 4% 이하로 매우 낮아 C. albicans와 차이를 보였다. 하지만 두 가지 진균에 서 공통적으로 은나노입자를 처리한 후 초기 시간에 세포주기가 G1에 지연되 는 것이 관찰되었다. 이 결과를 통해 은나노입자는 진균에서 세포주기를 G1에 지연시킴으로서 증식 억제 효과를 유발하며, 은나노입자에 대한 민감성은 C. albicans가 S. cerevisia에 비해 높다는 것을 확인하였다. 본 연구에서는 병원성, 비병원성 진균에서 5 nm 은나노입자의 효과를 세포사멸, 세포주기 분석 등 다 양한 방법으로 측정하였으며, 아직 보고된 바 없는 은나노입자의 G1기 지연 효 과를 보고하였다. 은나노입자의 항진균효과 기전에 대해서는 다양한 이견이 있는데, 그 중 한 가지는 활성산소종 생성에 관한 것이다. 기존 보고는 C. albicans에서 수산기 (hydroxy radical) 측정을 통해 은나노입자를 처리한 경우 활성산소종이 생성되어 진균에 손상을 가져온다고 보고하였다(Insok 등, 2012). 그러나 S. cerevisiae의 항 산화반응에 연관된 유전자인 YAP1 또는 SOD1, SOD2가 결손된 균주를 이용한 연구에서 은나노입자에 의한 이들의 증식 억제는 야생형과 차이가 없었다 (Käosaar 등. 2016). 본 연구에서는 이전에 연구된 바 없는 S. cerevisiae의 SKN7 단독 또는 YAP1과의 복합 유전자 결손 균주를 이용하여 S. cerevisiae에서 은나 노입자가 활성산소종 생성과 연관되어 있을 가능성이 적음을 한 번 더 입증하 였다. 또한 활성산소종 제거제인 NAC을 이용한 실험결과를 통해 C. albicans에 서 은나노입자의 항진균효과가 활성산소종 생성과 관련되어 있음을 증명하였다. 이것은 은나노입자가 세포 내 다양한 대사 경로에 영향을 미치며 그 기전은 진 균의 종에 따라 다를 수 있음을 시사한다. 그동안 S. cerevisiae에서 은나노입자의 항진균효과가 세포벽 스트레스와 연
- 29 - 관성은 명확하게 밝혀지지 않았다. 기존 연구에서 S. cerevisiae에 시트르산으로 코팅된 은나노입자를 처리하였을 때 세포벽의 손상이 유발되는 것을 투과전자 현미경을 이용하여 관찰하였다(Loredana 등 2014). 그러나 S. cerevisiae의 세포벽 스트레스에 대한 대응 반응을 유도하는 Kre6와 knr4 유전자의 결손 균주에서 은나노입자에 의한 증식 억제 효과는 야생형과 차이가 없음을 보여주었다 (Käosaar 등. 2016). 본 연구에서는 세포벽 스트레스와 은나노입자의 항진균효과 의 연관성을 알아보고자 지금까지 연구된 바 없는 HOG1과 SLT2의 단독 또는 복합 유전자 결손 균주를 이용하여 S. cerevisiae에서 은나노입자가 세포벽 스트 레스 반응과 연관이 적을 가능성을 보여주었다. 본 연구에서는 5 nm 은나노입자가 포도당 소비에 영향을 미치는지 알아보 았다. 그 결과 5 nm 은나노입자에 의해 C. albicans의 포도당 소비량이 증가한 반면 S. cerevisiae에서는 고농도의 5 nm 은나노입자에 의해 포도당 소비량이 감 소하였다. 포도당 소비에서도 은나노입자는 두 가지 균주에서 서로 다른 효과 를 나타냈다. 이 결과만으로 기전을 설명하기는 어려우나 은나노입자의 항진균 효과가 포도당 이용과 연관되어 있을 가능성을 배제할 수 없다. 또한 C. albicans와 S. cerevisiae는 진균에 속하지만 유전적인 상동성이 20% 미만으로 낮
기(Jonathan 등, 2014) 때문에 C. albicans와 S. cerevisiae에서 은나노입자의 항진균 효과는 서로 다른 세포 내 대사 경로와 연관되어 있을 것이라고 생각할 수 있 다. 진균의 포도당 이용에 은나노입자가 어떻게 영향을 미치는지 자세히 알아 보기 위해서는 더 많은 연구가 필요하다. 본 연구에서는 그동안 상반된 의견이 존재했던 은나노입자의 항진균효과 기전을 밝히고자 두 가지 진균에서 다양한 실험을 수행하였다. 그 결과 은나노 입자의 항진균효과는 C. albicans에서 활성산소종 생성과 연관이 있음을 확인하 였다. 또한 S. cerevisiae에서 은나노입자의 항진균효과는 활성산소종 및 세포벽 스트레와 관련이 없음을 입증하였다. 또한 본 연구에서는 기존 연구에서 사용 하지 않았던 NAC과 S. cerevisiae 유전자 결손 균주를 이용해 이를 증명하고자
- 30 - 하였다는 점에서 차별성이 있다. 또한 이전 논문에서 수행하지 않았던 세포주 기 분석을 통해 은나노입자가 다양한 세포 대사 경로를 통해 G1 지연 효과를 가져오고 그로 인해 항진균효과가 나타남을 설명하였다. 서로 다른 진균에서 포도당 소비량을 측정함으로서 은나노입자가 진균의 포도당 이용 과정에 관여 할 가능성을 제시하였다. 은나노입자는 항진균제에 내성을 갖는 병원성 진균의 치료제로 대체될 가 능성을 가지고 있다. 은나노입자가 새로운 항진균제로서 효과를 높이기 위해서 는 아직 명확하지 않은 은나노입자의 항진균효과의 기전을 밝히기 위해 더 많 은 연구가 필요하다. 본 연구는 그동안 여러 갈래로 나뉘었던 은나노입자의 항 진균효과 기전 연구의 방향을 제시하였으며, 이 결과는 후에 포유류 동물 세포 에서 은나노입자의 독성 기전을 연구하는데 기초자료가 될 수 있다. 또한 항진 균제로서 은나노입자의 가치를 확인하는 연구가 될 것이다.
- 31 -
V.
결 론
본 연구에서는 PVP로 코팅된 100, 5 nm 은나노입자의 효과를 C. albicans와 S. cerevisiae에서 확인하고, S. cerevisiae의 항산화반응 및 세포벽 스트레스 반응 과 관련한 6가지 유전자의 결핍 모델과 NAC을 이용하여 은나노입자의 항진균 효과 기전을 밝히고자 하였다. 세포 증식 및 세포 주기 분석 결과 C. albicans와 S. cerevisiae 모두 공통적으로 5 nm 은나노입자에 의해 세포 증식이 억제되었으 며, 세포 주기가 G1기에서 지연됨을 관찰하였다. 그러나 두 가지 균주에서 5 nm 은나노입자에 의한 세포 사멸은 C. albicans에서만 유도되는 차이를 보였다. 포도당 소비량을 측정한 결과 5 nm 은나노입자를 처리한 경우 C .albicans에서는 포도당 소비량은 증가하였으나 S. cerevisiae에서는 감소하였다. NAC을 은나노입 자와 함께 처리한 후 세포주기를 분석한 결과 S. cerevisiae에는 NAC이 효과를 나타내지 못하였으나 C. albicans에서는 NAC이 은나노입자에 의한 G1 지연 효 과를 크게 감소시켰다. 따라서 5 nm 은나노입자는 두 가지 진균에서 서로 다른 기전에 의해 항진균효과를 나타냄을 알 수 있었다. 결과적으로 은나노입자의 항진균효과는 C.albicans에서는 활성산소종의 생성과 연관되어 있으며 S. cerevisiae에서는 활성산소종 생성 및 세포벽 스트레스와 관련이 없음을 확인하 였다.- 32 -
참 고 문 헌
1. Kanafani ZA, Perfect JR. Perfect Resistance to Antifungal Agents: Mechanisms and Clinical Impact. Clinical Infectious Diseases. 2008;46(1):120-128.
2. Christina Graf, Dirk L. J. Vossen, Arnout Imhof, andAlfons van Blaaderen. A General Method To Coat Colloidal Particles with Silica. Langmuir. 2003;19 (17), pp 6693–6700.
3. Morano KA, Grant CM, Moye-Rowley WS. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics. 2012;190(4):1157-1195.
4. Franci G, Falanga A, Galdiero S, Palomba L, Rai M, Morelli G, Galdiero M. Silver nanoparticles as potential antibacterial agents. Molecules. 2015;20(5):8856-8874.
5. Morones JR, Elechiguerra JL, Camacho A, Holt K, Kouri JB, Ramírez JT, Yacaman MJ. The bactericidal effect of silver nanoparticles. Nanotechnology. 2005;(10):2346-2253.
6. Biel MA, Sievert C, Usacheva M, Teichert M, Wedell E, Loebel N, Rose A, Zimmermann R. Reduction of Endotracheal Tube Biofilms Using Antimicrobial Photodynamic Therapy. Lasers Surg Med. 2011;43(7):586-590.
7. Paredes D, Ortiz C, Torres R. Synthesis, characterization, and evaluation of antibacterial effect of Ag nanoparticles against Escherichia coli O157:H7 and methicillin-resistant
Staphylococcus aureus (MRSA). Int J Nanomedicine. 2014;9:1717-1729.
8 Jain N, Bhargava A, Majumdar S, Tarafdar JC, Panwar J. Extracellular biosynthesis and characterization of silver nanoparticles using Aspergillus flavus NJP08: a mechanism
- 33 -
perspective. Nanoscale. 2011;3(2):635-641.
9. Xu Y, Gao C, Li X, He Y, Zhou L, Pang G, Sun S. In vitro antifungal activity of silver nanoparticles against ocular pathogenic filamentous fungi. J Ocul Pharmacol Ther. 2013;29(2):270-274.
10. Collins TL, Markus EA, Hassett DJ, Robinson JB. The effect of a cationic porphyrin on Pseudomonas aeruginosa biofilms. Curr Microbiol. 2010;61(5):411-416.
11. Dutta S, Shome A, Kar T, Das PK. Counterion-induced modulation in the antimicrobial activity and biocompatibility of amphiphilic hydrogelators: influence of in-situ-synthesized Ag-nanoparticle on the bactericidal property. Langmuir. 2011 Apr 19;27(8):5000-5008.
12. Ivask A, Elbadawy A, Kaweeteerawat C, Boren D, Fischer H, Ji Z, Chang CH, Liu R, Tolaymat T, Telesca D, Zink JI, Cohen Y, Holden PA, Godwin HA. Toxicity mechanisms in Escherichia coli vary for silver nanoparticles and differ from ionic silver. ACS Nano. 2014;8(1):374-386.
13. Kalhapure RS, Sonawane SJ, Sikwal DR, Jadhav M, Rambharose S, Mocktar C, Govender T. Solid lipid nanoparticles of clotrimazole silver complex: An efficient nano antibacterial against Staphylococcus aureus and MRSA. Colloids Surf B Biointerfaces. 2015;136:651-658.
14. Gomez M, Pérez-Gallardo RV, Sánchez LA, Díaz-Pérez AL, Cortés-Rojo C, Meza Carmen V, Saavedra-Molina A, Lara-Romero J, Jiménez-Sandoval S, Rodríguez F, Rodríguez-Zavala JS, Campos-García J. Malfunctioning of the iron-sulfur cluster assembly
- 34 -
machinery in Saccharomyces cerevisiae produces oxidative stress via an iron-dependent mechanism, causing dysfunction in respiratory complexes. PLoS One. 2014;9(10):e111585.
15. Loredana S. Dorobantu, Clara Fallone, Adam J. Noble, Jonathan Veinot, Guibin Ma, Greg G. Goss, Robert E. BurrellToxicity of silver nanoparticles against bacteria, yeast, and algae J Nanopart Res. 2015;17:172
16. Käosaar S, Kahru A, Mantecca P, Kasemets K Profiling of the toxicity mechanisms of coated and uncoated silver nanoparticles to yeast Saccharomyces cerevisiae BY4741 using a set of its 9 single-gene deletion mutants defective in oxidative stress response, cell wall or membrane integrity and endocytosis. Toxicol In Vitro. 2016;35:149-162.
17. Rosário F, Hoet P, Santos C, Oliveira H. Death and cell cycle progression are differently conditioned by the AgNP size in osteoblast-like cells. Toxicology. 2016;368-369:103-115.
18. Franci G, Falanga A, Galdiero S, Palomba L, Rai M, Morelli G, Galdiero M. Silver nanoparticles as potential antibacterial agents. Molecules. 2015;20(5):8856-74.
19. Botstein D, Chervitz SA, Cherry JM. Yeast as a model organism. Science. 1997;277(5330):1259-1260.
20. Mulford KE, Fassler JS. Association of the Skn7 and Yap1 transcription factors in the
Saccharomyces cerevisiae oxidative stress response. Eukaryot Cell. 2011;10(6):761-769.
21. Zhao X, Mehrabi R, Xu JR. Mitogen-activated protein kinase pathways and fungal pathogenesis. Eukaryot Cell. 2007;6(10):1701-1714.
- 35 -
22. Lee MJ, Lee SJ, Yun SJ, Jang JY, Kang H, Kim K, Choi IH, Park S. Silver nanoparticles affect glucose metabolism in hepatoma cells through production of reactive oxygen species. Int J Nanomedicine. 2015;11:55-68.
23. Musa SF, Yeat TS, Kamal LZM, Tabana YM, Ahmed MA, El Ouweini A, Lim V, Keong LC, Sandai D. Pleurotus sajor-caju can be used to synthesize silver nanoparticles with antifungal activity against Candida albicans. J Sci Food Agric. 2017.
24. Xue B, He D, Gao S, Wang D, Yokoyama K, Wang L. Biosynthesis of silver nanoparticles by the fungus Arthroderma fulvum and its antifungal activity against genera of Candida, Aspergillus and Fusarium. Int J Nanomedicine. 2016;11:1899-1906.
25. Hwang IS, Lee J, Hwang JH, Kim KJ, Lee DG. Silver nanoparticles induce apoptotic cell death in Candida albicans through the increase of hydroxyl radicals. FEBS J. 2012;279(7):1327-1338.
26. Binkley J, Arnaud MB, Inglis DO, Skrzypek MS, Shah P, Wymore F, Binkley G, Miyasato SR, Simison M, Sherlock G. The Candida Genome Database: the new homology information page highlights protein similarity and phylogeny. Nucleic Acids Res. 2014;(42):D711-716.
27. Lee J, Romeo A, Kosman DJ. Transcriptional remodeling and G1 arrest in dioxygen stress in Saccharomyces cerevisiae. J Biol Chem. 1996;271(40):24885-24893.
28. Zinzalla V, Graziola M, Mastriani A, Vanoni M, Alberghina L. Rapamycin-mediated G1 arrest involves regulation of the Cdk inhibitor Sic1 in Saccharomyces cerevisiae. Mol
- 36 -
Microbiol. 2007;63(5):1482-1494.
29. de Bruin RA, Kalashnikova TI, Chahwan C, McDonald WH, Wohlschlegel J, Yates J 3rd, Russell P, Wittenberg C. Constraining G1-specific transcription to late G1 phase: the MBF-associated corepressor Nrm1 acts via negative feedback. Mol Cell. 2006;23(4):483-496.
30. Newcomb LL, Diderich JA, Slattery MG, Heideman W. Glucose regulation of
Saccharomyces cerevisiae cell cycle genes. Eukaryot Cell. 2003 Feb;2(1):143-149.
31. Abdul Razack S, Duraiarasan S, Mani V. Biosynthesis of silver nanoparticle and its application in cell wall disruption to release carbohydrate and lipid from C. vulgaris for biofuel production. Biotechnol Rep 2016;11:70-76
32. Panácek A, Kolár M, Vecerová R, Prucek R, Soukupová J, Krystof V, Hamal P, Zboril R, Kvítek L. Antifungal activity of silver nanoparticles against Candida spp. Biomaterials. 2009;30(31):6333-6340.
- 37 -
- ABSTRACT -
The inhibitory effect of 5 nm silver nanoparticles on the growth of
fungi and its underlying mechanisms
Bokyoung Lee
Department of Microbiology, Ajou University
(Supervised by Professor Sun Park)
Silver nanoparticles(AgNPs) have shown the inhibitory effect on the growth of various fungi, however, the underlying mechanisms of AgNPs remain poorly understood. In this study, the antifungal effect of PVP-coated 5 nm AgNPs on Candida albicans and Saccharomyces cerevisiae and its underlying mechanisms was investigated. The inhibitory effect of AgNPs on the proliferation and cell cycle progression was observed in both fungi. However, increase in cell death and glucose consumption was observed only in C. albicasns. Further, analysis of proliferations of S. cerevisiae mutants(∆SKN7, ∆YAP1, ∆HOG1 or ∆SLT2) and analysis of cell cycle in the presence of ROS scavenger showed the relevance of reactive oxygen species with the effect of AgNPs in C. albicans but not in S. cerevisiae. My results suggest the differential action mechanisms of underlying the antifungal effect of AgNPs.
Keyword: Silver nanoparticles, Growth inhibitory effect, Candida albicans, Saccharomyces