저작자표시-비영리-동일조건변경허락 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. l 이차적 저작물을 작성할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 동일조건변경허락. 귀하가 이 저작물을 개작, 변형 또는 가공했을 경우 에는, 이 저작물과 동일한 이용허락조건하에서만 배포할 수 있습니다.
Gingko biloba extracts protect
auditory hair cells from
cisplatin-induced ototoxicity by inhibiting
perturbation of gap junctional
intercellular communication
by
Seong Jun Choi
Major in Medicine
Department of Medical Sciences
The Graduate School, Ajou
Gingko biloba extracts protect auditory hair cells from
cisplatin-induced ototoxicity by inhibiting perturbation
of gap junctional intercellular communication
by
Seong Jun Choi
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements for
the Degree of Ph.D.
in Medicine
Supervised by
Yun-Hoon Choung, M.D., D.D.S., Ph.D.
Major in Medicine
Department of Medical Sciences
The Graduate School, Ajou University
February, 2014
This certifies that the dissertation
of Seong Jun Choi In is approved.
SUPERVISORY COMMITTEE
Keehyun Park
Yun-Hoon Choung
Chul- Ho Kim
Seon-Yong Jeong
Shi-Nae Park
The Graduate School, Ajou University
November, 4, 2013
i
-ABSTRACT-
Gingko biloba extracts protect auditory hair cells from
cisplatin-induced ototoxicity by inhibiting perturbation of gap
junctional intercellular communication
Gap junctional intercellular communication (GJIC) may play an important role in the hearing process. Cisplatin is an anticancer drug that causes hearing loss and Gingko biloba extracts (EGb 761) have been used as an antioxidant and enhancer for GJIC. The purpose of this study was to examine the efficiency of EGb 761 in protecting against cisplatin-induced apoptosis and disturbance of GJIC. HEI-OC1 auditory cells were cultured and treated with cisplatin (50µM) and EGb (300µg/ml) for 24 h, and then analyzed by immunocytochemistry (Annexin V/ propidium iodide) and Western blots. The function of GJIC was evaluated by scrape-loading dye transfer (SLDT). Basal turn organ of Corti (oC) explants from neonatal (p3) rats were exposed to cisplatin (1–10µM) and EGb (50–400µg/ml). The number of intact hair cells was counted by co-labeling with phalloidin and MyoVIIa. EGb prevented cisplatin-induced apoptosis in immunostaining and decreased caspase 3 and poly-ADP- ribose polymerase (PARP) bands, which were increased in cisplatin-treated cells in Western blots. EGb prevented abnormal intracellular locations of Cx 26, 30, 31, and 43 in cells treated with cisplatin and increased quantities of Cx bands. EGb also prevented cisplatin-induced disturbance of GJIC in SLDT. In oC explants, EGb significantly prevented hair cell damage induced by cisplatin. In animal studies, EGb significantly prevented cisplatin-induced hearing loss across 16 kHz and 32 kHz. These results show that cisplatin induces ototoxicity including hearing loss as well as down-regulation of GJIC and inhibition of Cxs in auditory cells. EGb prevents hearing loss in cisplatin-treated rats by inhibiting down-regulation of Cx expression and GJIC. The disturbance of GJIC or Cx expression may be one of important mechanisms of cisplatin-induced ototoxicity.
Key word : Gap Junction, Connexin, Hearing loss, Apoptosis, Gingko biloba extracts, Cisplatin
ii
TABLE OF CONTENTS
ABSTRACT∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ i TABLE OF CONTENTS ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ ii LIST OF FIGURES∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ iii ABBREVIATION∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ iv Ⅰ. INTRODUCTION∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 1 Ⅱ. MATERIAL AND METHODS∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 7 A. In vitro experiments – apoptosis of auditory cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙7 1. Auditory cell culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 7 2. Cell viability assay ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙7 3. 4'6-Diamidino-2-phenylindole (DAPI) staining ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙8 4. Western blot analysis of caspase-3, poly-ADP-ribose polymerase (PARP), Cx26, and Cx43 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 8 5. Quantitative real time-polymerase chain reaction (qRT-PCR) of Cx26, Cx30,
Cx31, and Cx43 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙9 6. Immunocytochemistry of Cx26, Cx30, Cx31, and Cx43 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙10 7. Scrape-loading dye transfer (SLDT) assay ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙10 B. Ex vivo experiments - in vitro organ of Corti (oC) culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙12 C. In vivo experiments ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙13 1. Auditory Brainstem Response (ABR) Test ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙13 2. Phalloidin staining ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙14 3. Scanning electron microscope (SEM) ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙14 Ⅲ. RESULTS ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙ 16 A. In vitro experiments – apoptosis of auditory cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙16 1. Protective effects of EGB 761 on cisplatin-induced death in HEI-OC1 cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙16 2. Protective effects of EGB 761 on cisplatin-induced apoptosis ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙18 3. Protective effects of EGb 761 on the inhibition of GJIC by cisplatin ∙∙∙∙∙∙∙∙∙20 B. Ex vivo experiments - in vitro oC culture ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙26 C. In vivo experiments ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙28
iii
1. Shift of ABR thresholds ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙28 2. Cochlear morphology - phalloidin staining and SEM ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙30 Ⅳ. DISCUSSION ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙32 Ⅴ. CONCLUSION ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙39 REFERENCES ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙40 국문요약 ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙56
iv
LIST OF FIGURES
Fig. 1. The effect of cisplatin on antioxidant system in cochea ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙2 Fig. 2. Gap junctions in cellular homeostasis ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙4 Fig. 3. Cisplatin-induced cytotoxicity of HEI-OC1 cells in MTT assay ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙17 Fig. 4. EGB 761 reduces apoptosis in cisplatin-treated HEI-OC1 cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙19 Fig. 5. mRNA and protein expression of Cxs in HEI-OC1 cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙21 Fig. 6. Immunocytochemistry of Cxs in HEI-OC1 cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙23 Fig. 7. SLDT experiment to evaluate functional GJIC ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙25 Fig. 8. Immunohistochemistry of in oC explants ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙27 Fig. 9. Shifts of ABR thresholds and morphology of oC explants ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙29 Fig. 10. Phalloidin staining and SEM of cochlear hair cells ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙31
v
ABBREVIATION
ABR, auditory brainstem response; ANOVA, analysis of variance; BSA, bovine serum
albumin; CDDP, cis-diamine-dichloroplatinum II; Cx, connexin; DAPI,
4'6-diamidino-2-phenylindole; DMEM, Dulbecco’s modified Eagle medium; DMSO,
dimethyl sulfoxide; EGb, extract of Gingko biloba; FBS, fetal bovine serum; GJIC,
gap junctional intercellular communication; HEI-OC1, House Ear Institute-Organ of
Corti 1; IHC, inner hair cell; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl
tetrazolium bromide; oC, organ of Corti; OHC, outer hair cell; PARP, poly-ADP- ribose
polymerase; PBS, phosphate buffered saline; ROS, reactive oxygen species; RT-PCR, real time- polymerase chain reaction; SD, standard deviation; SDS-PAG, sodium
dodecyl sulfate-polyacrylamide gel; SEM, scanning electron micrograph; SLDT, Scrape-loading dye transfer; SOD, super oxide dismutase; TBS, Tris buffer solution
- 1 -
I.
INTRODUCTION
Cisplatin (cis-diamminedichloroplatinum; CDDP), a potent inducer of growth arrest and/or apoptosis in most cell types, has been clinically used for decades against a variety of human malignancies, including ovarian, testicular, bladder, head and neck, esophageal, and small-cell lung cancers The major antineoplastic effect of cisplatin is its binding to DNA and the inhibition of cell replication (Siddik, 2003). Cisplatin-induced cell death has been associated with both necrosis and apoptosis. Apoptosis is an active form of cell death characterized by energy dependence, DNA and nuclear condensation and fragmentation, cell shrinkage and formation of apoptotic bodies in presence of an intact cell membrane and expression of several biochemical hallmarks.(Previati et al., 2007)
However, the risk of ototoxic and nephrotoxic side effects commonly hinders the use of higher doses that could maximize its antineoplastic effects (Humes, 1999). Cisplatin-induced ototoxicity is usually manifested by bilateral, progressive, and usually irreversible sensorineural hearing loss. Ototoxicity has a significant negative impact on patient quality of life, especially in pediatric cases, where more than 50% of children develop ototoxicity (Knight et al., 2005). Even minimal and mild hearing loss in high-frequency regions above 2000 Hz considerably increase a child’s risk for academic difficulties, social-emotional problems (Knight et al., 2005), and increased levels of fatigue in the learning environment (Bess et al., 1998).The presence of these side effects has prompted research to achieve a better understanding of the biochemical mechanisms underlying the cytotoxicity.
Cisplatin ototoxicity has been shown to have at least three major tissue targets in the cochlea: organ of Corti, spiral ganglion cells and lateral wall (stria vascularis and spiral ligament) (Rybak LP et al., 2007). Studies in guinea pig reveal that cisplatin affects both the organ of Corti (primarily the outer hair cells) and the spiral ganglion cells (Van
- 2 -
Ruijven et al., 2005). Cisplatin ototoxicity in rats was manifested by deleterious effects on the basal turn stria vascularis, including strial edema, bulging, rupture and compression of the marginal cells and depletion of organelles from the cytoplasm (Meech et al., 1998). The ototoxic effect of cisplatin on lateral wall tissues in the gerbil was further substantiated with in vitro studies of the spiral ligament(Liang et al., 2005). Platinated DNA was immunolocalized to the nuclei of outer hair cells, supporting cells of the organ of Corti, marginal cells of the stria vascularis and the cells in the spiral ligament of the basal turn (Van Ruijven et al., 2005b). This finding confirms that these cells are targets for cisplatin toxicity (Van Ruijven et al., 2005).
Fig. 1. The effect of cisplatin on antioxidant system in cochea. Cisplatin depletes the antioxidant sytem in the cochlea. Cisplatin treatment leads to the depletion of glutathione and the antioxidant enzymes, allowing an increase in lipid peroxidation (Somani et al., 2001).
- 3 -
Reactive oxygen species, such as superoxide anions (Dehne et al., 2001), are among the important factors that lead to cisplatin-induced ototoxicity. Cochlear tissues from animals receiving ototoxic doses of cisplatin were depleted of glutathione and antioxidant enzymes (superoxide dismutase, catalase, and glutathione peroxidase and glutathione reductase) with a reciprocal increase in malondialdehyde levels, an indicator of lipid peroxidation (Rybak et al., 2000). Superoxide leads to the formation of hydrogen peroxide, which when catalyzed by iron forms the highly reactive hydroxyl radical and leads to damaging lipid peroxidation. Peroxynitrite, formed by the reaction of superoxide with nitric oxide, reacts with susceptible proteins to form nitrotyrosine, an indicator of oxidative damage to proteins (Radi et al., 2001). Protein nitration can also be derived by peroxidase-mediated mechanism (van der Vliet et al., 1997). Depletion of cochlear antioxidant enzyme activities may result from: (1) direct binding of cisplatin to essential sulfhydryl groups within the enzymes (Rybak LP et al., 2007).; (2) depletion of copper and selenium, which are essential for superoxide dismutase and glutathione peroxidase activities (DeWoskin and Riviere, 1992); (3) increased ROS and organic peroxides which inactivate antioxidant enzymes (Pigeolet et al., 1990); and (4) depletion of glutathione and the cofactor NADPH, which are essential for the activities of glutathione peroxidase and glutathione reductase activities (Somani et al., 2001). Inhibition of antioxidant enzyme activity by cisplatin can allow ROS, such as superoxide and hydrogen peroxide, and toxic lipid peroxides to increase within the cochlea. This can lead to calcium influx within cochlear cells, leading to apoptosis (Clerici et al., 1995).
Protection from cisplastin-induced ototoxicity has been widely studied, because hearing preservation is crucial for the patient’s quality of life. The cochlea has endogenous mechanisms to deal with oxidative stress caused by agents like cisplatin. Protective molecules include: glutathione and the antioxidant enzymes, heat shock proteins, adenosine A1 receptors, NRF2 and heme-oxygenase-1, and the kidney injury molecule (KIM-1) (Mukherjea et al., 2006). Exogenous antioxidants provide upstream protection of the cochlea to prevent initiation of cell death. Several thiol antioxidants protect against cisplatin ototoxicity. The rationale for the use of these agents is the high
- 4 -
affinity of sulfur for platinum (Rybak and Whitworth, 2005). Such thiols include: sodium thiosulfate (Leitao et al., 2003), D- or L-methionine (Campbell et al., 1996), N-acetylcysteine (Najafi et al., 2009), glutathione (GSH) (Hamers et al., 1993) or glutathione ester (Campbell et al., 2003) and amifostine (Rybak and Whitworth, 2005). Perilymphatic perfusion of sodium thiosulfate in guinea pigs prevents cisplatin ototoxicity (Wang et al., 2003). Other antioxidant agents that are free radical scavengers that protect the cochlea from cisplatin damage and hearing loss in experimental animals include: alpha-tocopherol (alone or in combination with tiopronin), aminoguanidine, sodium salicylate and ebselen (alone or in combination with allopurinol, which is an inhibitor of xanthine oxidase) (Lynch et al., 2005a,b; Rybak and Whitworth, 2005).
Fig. 2. Gap junctions in cellular homeostasis. Endogenous and exogenous extracellular signals which can trigger various intracellular signal transduction mechanisms can either increase or decrease gap junctional intercellular communication between cells in a multicellular organism. Cell proliferation, differentiation, apoptosis, and adaptive responses of differentiated cells can occur as a consequence of the modulation of gap junctional intercellular communication (Trosko et al., 1998).
- 5 -
All tissues or organs require intercellular as well as intracellular communications to differentiate and operate normal function. Gap-junctional intercellular communication has long been hypothesized to play a crucial role in the maintenance of homeostasis, morphogenesis, cell differentiation, and growth control in multi cellular organisms. Gap junctions are channels connecting adjacent cells and allow molecular movements by passive diffusion based on molecular size. The molecules smaller than 1,000 Da such as cAMP and inorganic ions (Na?, K?, Ca2? etc.) can pass through gap junctions, but proteins or nucleic acids cannot. The cells communicate with neighboring cells by transfer of these small molecules. This gap junctional intercellular communication (GJIC) is an important mechanism to maintain tissue homeostasis, normal cell growth, differentiation, and development (Loewenstein, 1979). Gap junctions are made of two hemi-channels, called connexons, and each in turn is composed of six molecules of the membrane-spanning connexin (Cx) protein (Unwin et al., 1980).
Gap junctions directly connect the cytoplasms of neighboring cells, thereby mediating direct intercellular communication of cytoplasmic signaling molecules. Almost every cellular and tissue level process is affected by gap junction, including differentiation, migration, and apoptosis (Willecke et al., 2002; Mesnil et al., 2005). Many studies have shown that gap junctions play important roles in cancer biology and drug resistance (Prochazka et al., 2007; Trosko et al., 2001). It has been reported that cisplatin toxicity is enhanced by the presence of functional gap junctions between the target cells (Jensen et al., 2004). Gap junction expression acted additively with cisplatin in promoting apoptosis, cell cycle arrest, and down-regulation of BCL-2 in bladder cancer cell lines (Tanaka et al., 2001). Also, functional change or mutation of Cx has been linked to diverse functional impairments such as hearing loss or inflammatory skin lesions (Choung et al., 2002; Choung et al., 2006) and cisplatin or hydrophobic platinum complex induces apoptosis or premature senescence with the inhibition of gap-junctional intercellular communication (GJIC) (Procházka et al., 2007; Zhao et al., 2004). Therefore, we considered that the change of GJIC could be an important mechanism of cisplatin-induced ototoxicity.
- 6 -
Ginkgo is a dietary supplement derived from the deciduous tree, Ginkgo biloba, considered to be the world’s oldest living tree species, which contains unique chemical substances unknown in other living things. Extracts and infusions made from Ginkgo leaves have been used in traditional Chinese medicine for thousands of years (Major, 1967). G. biloba extract, EGb 761, is a standardized extract of G. biloba (Drieu, 1988), developed by Dr. Willmar Schwabe Pharmaceuticals in Germany, containing various components of the Ginkgo leaf so as to give maximal medicinal benefits. EGb 761 consists of 24% flavone glycosides (flavonoids), 6% terpene lactones (ginkgolides and bilobalide), 7% proanthocyaniclines and other uncharacterized compounds; one of the proposed mechanisms for its beneficial pharmacologic effect is its potent antioxidant action (Clostre, 1986). The demonstrated effects of EGb include vasomodulation, platelet-activating factor antagonism, antioxidation, antiapoptosis, and neuroprotection (Diamond et al., 2000; Guidetti et al., 2001). Studies have shown the in vitro and in vivo protective effects of EGb 761 against oxidative stress (Oyama et al., 1996; Rong et al., 1996), and these are closely related to its ability to scavenge free radicals, such as superoxide anion, hydroxyl, and peroxyl radicals (Pincemail et al., 1989; Maitral et al., 1995; Yoshikawa et al., 1999). Furthermore, EGb 761 also inhibits lipid peroxidation reactions and enhances antioxidant enzyme activities (Bridi et al., 2001). The free radical scavenging activity of EGb 761 is comparable to that of other known antioxidants such as vitamins A and C, as well as the tri-peptide GSH (Kose et al., 1995). EGb 761 also can scavenge nitric oxide and inhibit its production by nitric oxide synthase (NOS) (Sharma et al., 2000; Wong et al., 2001). In addition, EGb 761 has anti-inflammatory and antiapoptotic properties (Christen et al., 2002). In another study, EGb displayed neuroprotective effects by improving Cx43 expression and GJIC in ischemic or hypoxic injuries in rat brain (Li et al., 2005).
This study investigated the protection effect of EGb 761 on cisplatin-induced ototoxicity and the change of GJIC in auditory cells and in vivo.
- 7 -
II. Materials and methods
A. In vitro experiments – apoptosis of auditory cells
1. Auditory cell culture.
House Ear Institute-Organ of Corti 1 (HEI-OC1) cells are conditionally immortalized mice cochlear cells. They have been previously characterized (Devarajan et al., 2002). HEI-OC1 cells were cultured in high glucose Dulbecco’s modified Eagle medium (DMEM; GIBCO/BRL, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS; GIBCO/BRL) and 50 U/mL interferon-γ (Genzyme, Cambridge, MA, USA) in the absence of antibiotics.
2. Cell viability assay.
Cell viability was measured by an established
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)-based assay (Watson et al., 1998). Cells were seeded in 96 well plates (1 x 104 cells). To examine the effects of cisplatin and EGb 761 on cell viability, cells were exogenously treated with various concentrations of cisplatin (1, 5, 10, 20, 50, 100, and 300 µM) or EGb 761 (10, 25, 50, 100, 200, 300, and 600 µg/ml) for 24 h. In addition, to analyze the protective effects of EGb 761 against cisplatin, cells were pretreated with various concentrations of EGb 761 (5, 25, 50, 100, 300, and 600 µg/ml) for 24 h and then were continuously maintained with 50 µM cisplatin with EGb 761 for 24 h. After 24 h, the culture medium was discarded completely, and 0.5 mg/ml MTT was added at 100 μl/well. After a 4 h incubation at 33°C in the dark in a 5% CO2 atmosphere, MTT was aspirated and 100 µl dimethylsulfoxide (DMSO) was added to each well for 20 min to
- 8 -
dissolve the formazan crystals. The absorbance was measured using a Power Wave X ELISA reader (Bio-Tek, Winooski, VT, USA) at 540 nm. All experiments were repeated three times. The viability of the EGb and/or cisplatin treated cells was expressed as a percentage of population growth plus the standard error of the mean relative to that of control cells.
3. 4'6-Diamidino-2-phenylindole (DAPI) staining.
DAPI staining was performed to evaluate the nuclear morphology. The cells were cultured on four-chamber slides and fixed by incubating with 4% paraformaldehyde (PFA) for 30 min. They were washed with phosphate buffered saline (PBS) and incubated with 1 μg/ml DAPI for 30 min in the dark. Nuclear morphology was analyzed by fluorescence microscopy (Zeiss, Oberköchen, Germany). Apoptotic nuclei were identified by the condensed chromatin gathering at the periphery of the nuclear membrane or a total fragmented morphology of nuclear bodies. More than 150 cells were counted and the percentage of apoptotic nuclei was determined.
4. Western blot analysis of caspase-3, poly-ADP-ribose polymerase (PARP), Cx26, and Cx43.
Western blots were performed targeting Cx26 (antibody dilution of 1:200; Zymed Laboratories, South San Francisco, CA, USA), Cx43, caspase-3, and poly-ADP- ribose polymerase (PARP) (each antibody dilution of 1:1000; Cell Signaling Technology, Danvers, MA, USA) by the standard method. Cell lysates were used in Pro-prep lysis buffer (Intron Biotechnology, Seongnam-Si, Korea). The proteins were immediately heated for 5 min at 100°C and were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Separated proteins were transferred to nitrocellulose membranes, and Western blotting was performed using a gel loading kit and protein transfer system kit (BioRad Laboratories, Hercules, CA,
- 9 -
USA). Membranes were blocked by treatment with 5% skim milk in Tris buffer saline (TBS) and subsequently incubated with the primary polyclonal antibodies (caspase-3 and PARP, 1: 1000; Cx26, 1:200; Cx43, 1:1000). Immunoreactive bands were visualized with horseradish peroxidase-conjugated secondary antibody to caspase-3, PARP, Cx26, or Cx43 using an enhanced chemiluminescence assay (Amersham Pharmacia Biotech, Piscataway, NJ, USA) following the manufacturer’s instructions.
5. Quantitative real time-polymerase chain reaction (qRT-PCR) of Cx26, Cx30, Cx31, and Cx43.
Total RNA was isolated from HEI-OC1 cells according to the manufacturer’s instructions using an Easy-BLUE RNA extraction kit (iNtRON Biotech, Sungnam, Korea). The concentration of total RNA in the final eluants was determined by spectrophotometry. Total RNA (2.5 μg) was heated at 65°C for 10 min and then chilled on ice. Each sample was reverse-transcribed to cDNA for 90 min at 37°C using a cDNA synthesis kit (Amersham Pharmacia Biotech). The samples were analyzed by real-time PCR using the 7500 Fast Real-Time PCR System (Applied Biosystems, Tokyo, Japan) for quantitative evaluation. One microliter of cDNA was amplified with the SYBR® Premix Ex Taq (TaKaRa). PCR was performed with the following primers for mouse (Cx26: 5’ CAT TCC TCC TAG TCC AGA GGT TCA G 3’, 5’ GTG GCA GTT GTC GGC ATA TCC TAT C 3’; Cx30: 5’ TGA CAA CTC CTC CTC TAG GT 3’, 5’ GGT AAG GAA GGC CAG TCA TT 3’; Cx31: 5’ CCT GTC GCT GTC ACC ACA AG 3’, 5’ ACC AGG TAG CCA GCA CAG AG 3’; Cx43: 5’ ACT TCA ATG GCT GCT CCT CAC CAA C 3’, 5’ CTG GAA GGT CGC TGA TCC ACG ATA G 3’; glyceraldehyde 3-phosphate dehydrogenase (GAPDH): 5’ TCA ACA GCA ACT CCC ACT CTT CCA 3’, 5’ ACC CTG TTG CTG TAG CCG TAT TCA 3’). The annealing temperature was 62°C for Cx26, 52°C for Cx30, 57°C for Cx31, and 63°C for Cx43. Amplified fragment sizes for Cx26, Cx30, Cx31, and Cx43 were 294 bp, 353 bp, 252 bp, and 295 bp, respectively. Electrophoresis of the products were done on a 1.5% agarose gel and visualized by staining with ethidium bromide. Quantitative data were
- 10 -
expressed as the mean relative to that of control cells ± standard deviation (SD). The results were evaluated by one-way ANOVA test using SPSS® 12.0 software (SPSS, Chicago, IL, USA). The statistical level of significance was set as P < 0.05.
6. Immunocytochemistry of Cx26, Cx30, Cx31, and Cx43.
Immunocytochemistry was used to verify the expression and subcellular localization of Cx26, Cx30, Cx31, and Cx43. After treatment with chemicals for 24 h, the cells were fixed in 4% paraformaldehyde in PBS for 10 min at room temperature. Samples were washed with PBS twice and permeabilized with 0.15% Triton X-100 in PBS for 10 min and then blocked with 2% bovine serum albumin (BSA) in PBS containing 0.1% Tween 20 for 30 min. For identification of Cx26 and Cx30, cells were co-labeled overnight with polyclonal rabbit anti-Cx26 antibody (Zymed Laboratories) and monoclonal mouse anti-Cx30 antibody (Zymed Laboratories). For the expression of Cx31 and Cx43, cells were incubated overnight with monoclonal mouse anti-Cx31 antibody (Zymed Laboratories) and polyclonal rabbit anti-Cx43 antibody (Cell Signaling Technology). They were then incubated with fluorescein isothiocyanate (FITC)-conjugated AffiniPure goat anti-rabbit IgG (Jackson ImmunoResearch, West Grove, PA, USA) and CyTM3-conjugated AffiniPure rabbit anti-mouse IgG (Jackson ImmunoResearch) for 1 h at room temperature. Cells were visualized using a LSM510 confocal microscope (Carl Zeiss Meditec, Jena, Germany).
7. Scrape-loading dye transfer (SLDT) assay.
The SLDT technique was used for evaluating GJIC of cells (El-Fouly et al., 1987). HEI-OC1 cells were incubated until 80–90% cell confluence in 35 mm-diameter dishes. Cells were treated with 10 µM cisplatin, 300 µg/ml EGb 761, or the combination of both chemicals for 24 h. Cells were washed twice with PBS, flooded with PBS containing 0.5% Lucifer Yellow (Sigma-Aldrich), and then scrape-loaded with a
- 11 -
surgical scalpel. The dye solution was left on the cell monolayer for 10 min, and the cultures were rinsed with PBS. Gap junctional dye transfer was observed using an Axiovert 200 fluorescent microscope (Carl Zeiss Meditec). We converted the photographs taken to gray scale and measured the dye spreading distance from the base line (the scraped line) with Image J® software (NIH, Bethesda, MD). To accurately measure the dye spreading distance or reduce errors, we measured the distance between the base line and the furthest point at 22 points per picture. The experiment was repeated ten times and more than 20 pictures (microscopic visual fields) were included in the statistical analysis. Quantitative data were expressed as mean ± standard deviation (SD). The results were evaluated by one-way ANOVA t test using SPSS® 12.0 software (SPSS, Chicago, IL, USA). The statistical level of significance was set as P < 0.05.
- 12 -
B. Ex vivo experiments - in vitro organ of Corti (oC) culture
Explants consisting of the basal turn cochlear epithelium from Sprague–Dawley rats at postnatal day 3 or 4 (p3 or p4) were isolated as previously described (Van de Water et al., 1974; Sobkowicz et al., 1993). Explants were maintained in DMEM including 10% FBS, 25 mM HEPES, and 30 U/ml penicillin in eight-chamber slide culture plates (Lab-Tek II Chamber Slide; Nalge Nune International, Naperville, IL, USA). They were cultured in an incubator at 37 °C with 5% CO2 and 95% humidity. The cultures were maintained in the aforementioned initial medium for one day. Then, explants were exposed to the same medium containing cisplatin (1, 3, 6, or 12 µM) and EGb (50, 100, 200, or 400 µg/ml) for 2 days. The explants were then fixed with 4% paraformaldehyde for 20 min and washed twice with PBS. The explants were permeabilized with 5% Triton X-100 (Sigma-Aldrich) in PBS with 10% FBS for 20 min. Specimens were labeled with a rabbit polyclonal anti-MyoVIIa primary antibody (1:500; Proteus Science, Chagrin Falls, OH, USA) at room temperature for 2 h. After a PBS wash, specimens were co-labeled with Phalloidin-Texas Red (1:100; Invitrogen, Carlsbad, CA, USA) and with FITC-conjugated, donkey anti-rabbit secondary antibody (1:100; Invitrogen) at room temperature for 1 h. Specimens were then stained with DAPI (1:500) at room temperature for 10 min, washed two times with PBS, and visualized by fluorescence microscopy. Loss of stereocilia on hair cells was recorded if no stereocilia or cuticular plate could be observed in the appropriate location. Quantitative analysis was obtained by evaluating the 30 outer hair cells (OHCs) associated with 10 inner hair cells (IHCs) in a given microscope field and expressed as mean ± standard deviation (SD). All experiments were repeated six times and more than six explants were obtained on each condition. Three-to-four random microscope fields were counted for each explant, and these were averaged to obtain a value for that explants. Statistical analysis was performed with ANOVA test and Bonferroni test as posthoc test, and differences associated with P values < 0.05 were considered to be statistically significant.
- 13 -
C. In vivo experiments
The study was approved by the the Institutional Animal Care of Ajou University School of Medicine (112508-05). Healthy adult, 8-week-old, 200–300 g, male Sprague-Dawley rats (n = 105) were used. Before the procedures, all ears were examined and normal eardrums were confirmed by the surgical microscopic findings. We always used the same anesthetic method: rats were anesthetized with 0.025 ml/g avertin (1.25% tribromoethanol, intraperitoneal, i.p., injection). We divided rats into four groups and they were treated as follows: 1) control group (n = 15); 2) cisplatin group (13 mg/kg, i.p.; n = 34); 3) EGb 761 group (200 mg/kg, i.p.; n = 21); and 4) EGb 761 + cisplatin group (n = 27). Auditory brainstem responses (ABRs) were measured at the time of pre-injection and 72 h post-injection, and hearing threshold shifts were analyzed. After initial ABR testing, rats received a single intraperitoneal injection of either EGb 761 (200 mg/kg) or an equal volume of normal saline. After 1 h, each rat received a single i.p. injection of either cisplatin (13 mg/kg) or an equal volume of normal saline. The rats were housed for 72 h and, after follow-up ABR testing, the cochleae were harvested for cochlear morphology studies such as phalloidin stainning, scanning electron microscopy (SEM), and immunohistochemistry. We used Viberq’s method (Viberq and Canlon, 2004) to identify the locations corresponding to 16 and 32kHz in cochlear turns.
1. Auditory Brainstem Response (ABR) Test.
Rats were anesthetized as described previously. A differential active needle electrode was placed subcutaneously below the tested ear (inverting), a reference electrode at the vertex (noninverting), and a ground electrode below the contralateral ear. Acoustic stimuli consisted of tone bursts (5ms rise/fall time with no plateau, with a cos2 envelope , at 20/s) of different frequencies (16 and 32 kHz) which were digitally generated by SigGenRP software (Tucker-Davis Technologies, Gainesville, FL, USA)
- 14 -
and RX6 Piranha Multifunction Processor hardware (Tucker-Davis Technologies, Gainesville, FL, USA). Stimuli were delivered monaurally into the external auditory meatus of the rats using an EDC1 electrostatic speaker (Tucker-Davis Technologies, Gainesville, FL, USA). The evoked potentials were filtered (0.3-3.0kHz), averaged (500 waveforms)(,) and stored for offline analysis. ABR waveforms were checked in an electrically shielded, sound-treated booth and were recorded in 5 dB sound pressure level intervals declining from the maximum amplitude. The auditory threshold was defined as the stimulus intensity that evoked waveforms with a peak-to-peak voltage greater than 2 standard deviations (SD) of the background activity. (Cediel et al., 2006; Garcia-Pino et al., 2010) Data was obtained from each group. Comparison of mean values of ABR thresholds of each group was performed with ANOVA and Bonferroni test as posthoc test, and significance was defined as a P value < 0.05.
2. Phalloidin staining.
Sprague-Dawley rats as described above were anesthetized as described previously, and their cochleae were isolated, dissected, and perfused through the oval and round windows with 4% paraformaldehyde in 0.1 M PBS. The cochleae were kept in the fixative overnight at 4°C. After fixation, the cochleae were then rinsed with 0.1 M PBS and incubated in 1% osmium tetroxide overnight, followed by immersion in 5% EDTA for 3 days. Under microscopic magnification, the otic capsule was removed, followed by removal of the lateral wall, Reissner's membrane, and tectorial membrane. The remaining oC was stained with rhodamine phalloidin (stock solution of 200 U/ml methanol diluted 1:100 in PBS) for 1 h. The basal and middle portions of oCs were visualized by fluorescence microscope (Zeiss, Oberköchen, Germany).
3. Scanning electron microscope (SEM).
The cochleae were dissected and perfused through the oval and round windows with 2% glutaraldehyde in PBS by placing a Pasteur pipette with a modified butterfly
- 15 -
catheter. Each specimen was then placed in the same glutaraldehyde solution overnight. The specimen was perfused with 1% osmium tetroxide and placed on a tissue rotator for 15 min. Under the dissecting microscope, the bony capsule of the cochlea was carefully removed and the lateral wall was cut away to reveal the oC. The tissue was serially dehydrated in 50%, 70%, 90%, 95%, and 100% acetone. Each specimen was treated with hexamethyldisilazane and then dried and placed on a stub for sputter coating with gold. The basal (32kHz) and middle (16kHz) turn tissues were then viewed and photographs were taken with a JSM-5410 LV scanning electron microscope (JEOL, Tokyo, Japan).
- 16 -
III. RESULTS
A. In vitro experiments – apoptosis of auditory cells
1. Protective effects of EGB 761 on cisplatin-induced death in HEI-OC1 cells. Cisplatin decreased the viability of HEI-OC1 cells in a dose-dependent manner. Cells treated with 1, 5, 10, 20, 50, 100, and 300 μM cisplatin for 24 h showed viabilities from about 100% to 30.6%. EGb 761 at 200 μg/ml slightly promoted HEI-OC1 cell proliferation showing maximal 125.2% viability of cells compared to control cells. To evaluate the protective effect of EGb 761 against cisplatin, HEI-OC1 cells were simultaneously treated with 50 μM cisplatin and 5, 25, 50, 100, 300, or 600 μg/ml of EGb 761 for 24 h. They were pretreated with same concentration of EGb 761 for 24 h before adding cisplatin. EGb 761 significantly protected from cisplatin-induced cytotoxicity, as shown in Fig. 1. The protection was dose-dependent and the maximal protective effect was observed at about 300 μg/ml of EGb 761. Therefore, 300 μg/ml concentration of EGb 761 was used for this study as an optimal concentration.
- 17 -
Fig. 3. Cisplatin-induced cytotoxicity of HEI-OC1 cells in MTT assay.
Cisplatin decreased the viability of cells in a dose-dependent manner during 24h exposure (A). EGb 761 increased cell proliferation compared to the control cells (B) and also significantly prevented cell death in HEI-OC1 cells treated with cisplatin 50 μM (C). ANOVA test was used for statistical analysis. The percentage of Y axis means the ratio of cell numbers in each group compared with the control group. MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide, CDDP: cis-diamine- dichloroplatinum II
- 18 -
2. Protective effects of EGB 761 on cisplatin-induced apoptosis.
In DAPI staining, the nuclei of the normal control cells appeared round with homogenous intensity (Fig. 2A). However, 50 μM cisplatin alone induced characteristic features of apoptotic nuclei such as small sized, angulated, fragmented shape (Fig. 2B). EGb 761 successfully prevented cisplatin-induced apoptotic change in some cells, although this effect was not seen for all cells (Fig. 2C). This finding was also consistent with the results of the MTT assay. The data supported the view that EGb 761 has an anti-apoptotic effect against cisplatin-induced cell death in HEI-OC1cell line (Fig. 2D).
Caspase-3 and PARP activity could be involved in cisplatin-induced toxicity and related to apoptotic changes in cisplatin ototoxicity. As shown in Fig. 2C, 10 and 50 μM cisplatin increased the activity of caspase-3 and PARP. Pretreatment of HEI-OC1 cells with EGb 761, however, significantly decreased the activity of caspase-3 and PARP, compared with cells treated with cisplatin alone. Inhibitory effects of EGb 761 for caspase-3 and PARP were evident upon Western blotting (Fig. 2E).
- 19 -
Fig. 4. EGB 761 reduces apoptosis in cisplatin-treated HEI-OC1 cells. (A-C) DAPI staining showed characteristic apoptotic nuclei with small sized, angulated, fragmented shape in CDDP-treated cells (B). Cells (C) treated with 300 μg/ml EGB 761 with 50 μM cisplatin for 24 h showed well preserved normal nuclei (arrows) compared to CDDP. There was significant difference of the proportions of apoptotic cells among the groups (D). In Western Blot (E), activity of caspase 3 and PARP was increased in the CDDP group, but inhibited by EGb.
- 20 -
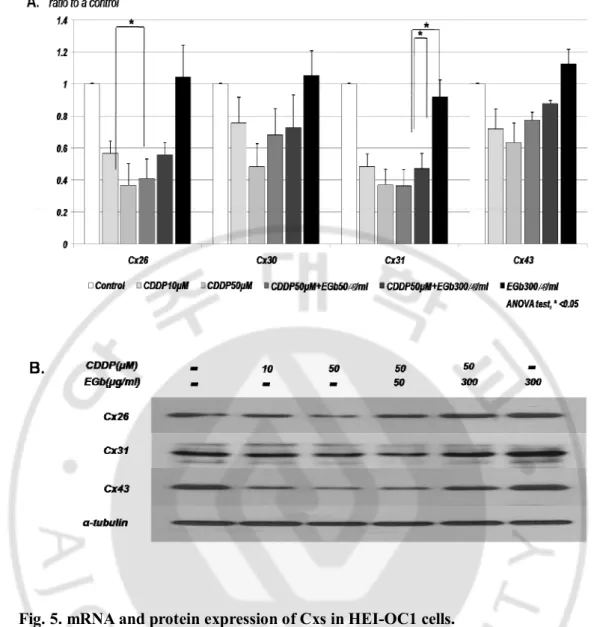
3. Protective effects of EGb 761 on the inhibition of GJIC by cisplatin.
As shown in Fig. 3A, the mRNA expression of Cx26, Cx30, Cx31, and Cx43 in HEI-OC1 cells treated with 10 and 50 μM cisplatin was significantly decreased when compared with the control group. However, pretreatment of HEI-OC1 cells with EGb 761 significantly exerted a protective effect on the mRNA level of Cx43 (both EGb 761 50 and 300 μg/ml) and 26 (only EGb 761 300 μg/ml) and there were also statistically significant differences (P < 0.05). In the Western blots (Fig. 3B), the intensity of Cx26, and 43 proteins was decreased in the presence of 50 μM cisplatin, but this down-regulation was inhibited by 50 and 300 μg/ml EGb 761. This suggests that EGb 761 has the preventive effect against down-regulation of Cxs induced by cisplatin in HEI-OC1cells.
- 21 -
Fig. 5. mRNA and protein expression of Cxs in HEI-OC1 cells.
(A) In quantitative PCR, mRNA expression of Cx26, Cx30, Cx31, and Cx43 in the cells treated with 10 and 50 μM cisplatin was significantly decreased in the dose-dependent manner, however, EGb 761 significantly increased the mRNA expression of Cx26and 43. (B) In the Western blot, the intensity of Cx26 and 43 were decreased under 50 μM cisplatin, but this was inhibited by 50 and 300 μg/ml EGb 761.
- 22 -
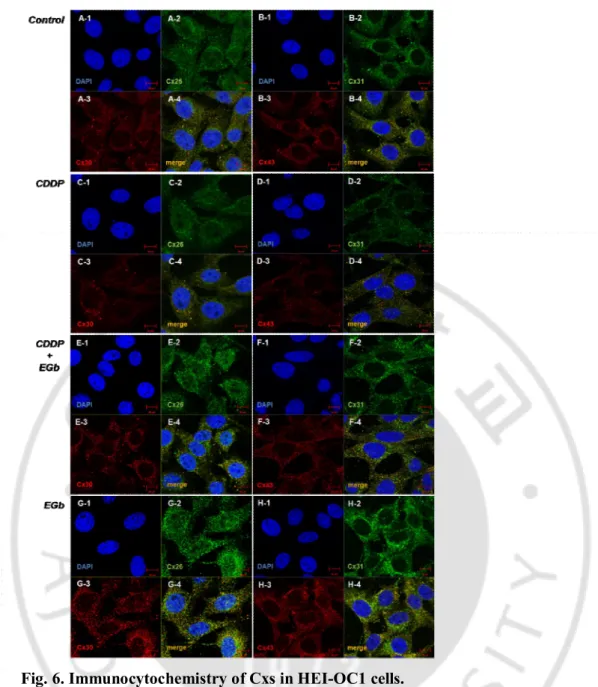
As shown in Fig 4, immunocytochemical analysis revealed the expression in the control group of Cx26, 30, 31, and 43. These Cxs displayed a granular or punctuate shape and were distributed exclusively in the cell membrane as well as cytoplasm (Fig. 4A, Fig. 4B). Compared with the control cells, cells treated with 50 μM cisplatin showed much less condensed Cxs in the cell membrane and much lower brightness of Cxs, and the expression of Cxs tended to aggregate just around the nuclei (Fig. 4C, Fig. 4D). However, in the cells treated with EGb 761 and CDDP, the expression of Cxs was increased and distributed wider in the cell membrane and cytoplasm like those of the control group (Fig. 4E, Fig. 4F).
- 23 -
Fig. 6. Immunocytochemistry of Cxs in HEI-OC1 cells.
The control group showed the expression of Cx26 (A-2), 30 (A-3), 31(B-2), and 43 (B-3) in a granular or punctuate shape and distributed in the cell membrane as well as cytoplasm. The expression of four kinds of Cxs (C-2, C-3, D-2, D-3) of HEI OC1 cells treated with 50 μM cisplatin was reduced and tended to aggregate just around the nuclei. However, the cells (E-2, E-3, F-2, F-3) treated with EGb 300 μg/ml and cisplatin 50 μM showed well preserved Cxs with similar patterns to the control group. Scale bar = 10 μm.
- 24 -
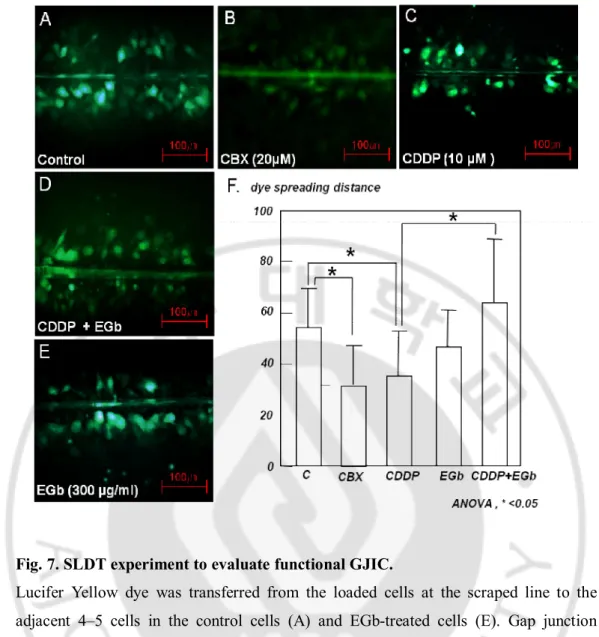
In the SLDT experiment (Fig. 5), normal control HEI-OC1 cells were well coupled by gap junctions. Lucifer Yellow dye was transferred from the loaded cells at the scraped line to the adjacent cells over 4–5 gradients (Fig. 5A). This dye transfer was significantly reduced when gap junction blocker, carbenoxolone (20 μM, 24 h) was treated on cells (Fig. 5B). Cells exposed to cisplatin (10 μM, 24 h) also showed a reduced capacity for dye transfer through three or fewer rows like carbenoxolone (Fig. 5C). These results indicated that cisplatin functionally inhibited GJIC in HEI-OC1 cells. However, EGb 761 up-regulated the inhibited dye transfer up to five or more rows of the adjacent cells, like the control group, in HEI-OC1 cells treated with cisplatin (Fig. 5D). Only EGb-treated cells show similar dye transfer to that in the control cells (Fig. 5E). This preventive effect of EGb 761 against cisplatin-induced down-regulation of GJIC was significantly prominent compared to the cisplatin group (Fig. 5F).
- 25 -
Fig. 7. SLDT experiment to evaluate functional GJIC.
Lucifer Yellow dye was transferred from the loaded cells at the scraped line to the adjacent 4–5 cells in the control cells (A) and EGb-treated cells (E). Gap junction blocker, Carbenoxolone (20 μM, 24 h) and cisplatin (10 μM, 24 h) significantly reduced the dye transfer (B and C). However, EGb 761 up-regulated the inhibited dye transfer up to five or more layers of the adjacent cells (D). ANOVA test was performed for statistical comparison (F). SLDT; Scrape-loading dye transfer, CBX; Carbenoxolone. Y axis: μm.
- 26 -
B. Ex vivo experiments - in vitro oC culture
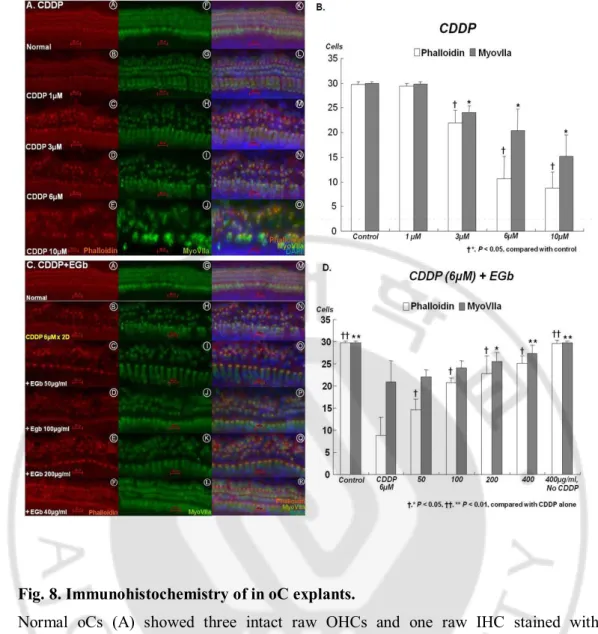
In immunohistochemical analysis (Fig. 6A), normal oCs showed the intact three row OHCs and one row IHCs stained with the hair cell-specific markers, phalloidin and MyoVIIa. As the concentration of cisplatin increased from 1–10 μM, the HC damage in the cochlea increased. At the low concentration of 1 μM cisplatin, the loss of stereocila was mainly detected in the first row OHCs. This damage spread to the second and third OHCs, and finally to IHCs depending on the concentration of cisplatin. There was a significant difference in the number of intact OHCs between the control group and the cisplatin group treated with ≥3 μM (P < 0.005) (Fig. 6B). The phalloidin staining was less than that of MyoVIIa in the same situation, suggesting that actin structures were easily damaged prior to myosin by cisplatin.
In the consequent study, we chose the concentration of 6 μM cisplatin because this concentration showed the definite change of oC hair cells but did not lead to complete destruction. As the concentration of EGb was increased from 50–400 μg/ml, the preservation of HCs in the cochlea was more readily identified (Fig. 6C). The minimal concentration of EGb that displayed a significant preventive effect for hair cells compared to the cisplatin group was 50 μg/ml in phalloidin staining, but 200 μg/ml in both phalloidin and MyoVIIa immunohistochemistry (Fig. 6D). EGb 761 efficiently prevented cisplatin-induced hair cell death in in-vitro oC culture.
- 27 -
Fig. 8. Immunohistochemistry of in oC explants.
Normal oCs (A) showed three intact raw OHCs and one raw IHC stained with phalloidin and MyoVIIa. As the concentration of cisplatin increased from 1–10 μM, the damage of hair cells in the cochlea increased. (B) There was a significant difference of the number of intact OHCs between the control group and the cisplatin group treated with ≥3 μM (P < 0.005). (B) The preservation of hair cells in the cochlea was more readily identified depending on the concentration of EGb (C, D). Scale bar = 20 μm.
- 28 -
C. In vivo experiments
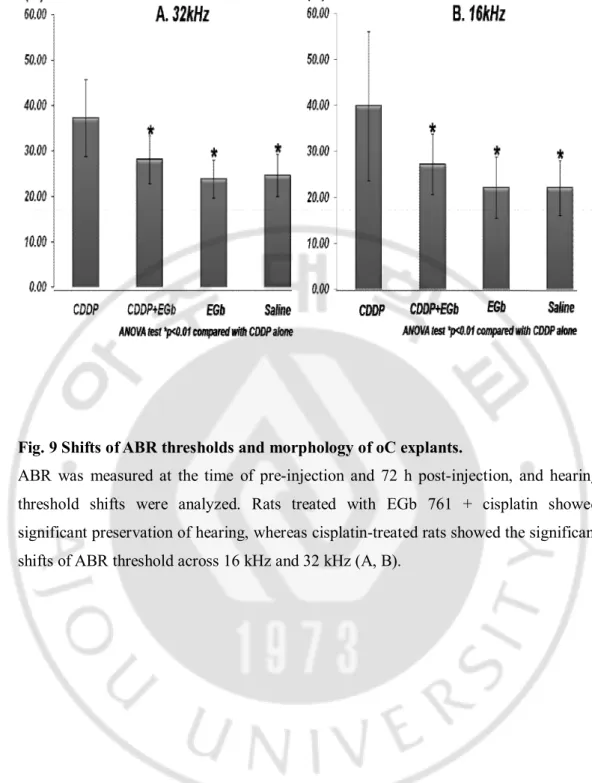
1. Shift of ABR thresholds.
The ABR thresholds of 16 and 32 kHz were 22.0 (± 5.9) dB and 24.5 (± 4.6) dB in the control group, 39.8 (± 16.2) dB and 37.1 (± 8.4) dB in the cisplatin group, 22.1 (± 6.6) dB and 23.8 (± 4.2) dB in the EGb 761 group, and 27.1 (± 6.5) dB and 28.1 (± 5.3) dB in the EGb + cisplatin group, respectively (Figs. 7A and 7B). Rats treated with EGb 761 + cisplatin showed significant preservation of hearing, whereas cisplatin-treated rats showed the significant shifts of ABR threshold across 16 kHz and 32 kHz (P < 0.001) (Figs. 7A and 7B).
- 29 -
Fig. 9 Shifts of ABR thresholds and morphology of oC explants.
ABR was measured at the time of pre-injection and 72 h post-injection, and hearing threshold shifts were analyzed. Rats treated with EGb 761 + cisplatin showed significant preservation of hearing, whereas cisplatin-treated rats showed the significant shifts of ABR threshold across 16 kHz and 32 kHz (A, B).
- 30 -
2. Cochlear morphology - phalloidin staining and SEM
Both the IHCs and the OHCs of the control group and the EGb 761 group showed normal appearance of stereocilia in every turn of the cochleae in SEM and phalloidin staining studies (Figs. 7C, 7G, 7K, 7F, 7J, and 7N). However, for the rats treated with cisplatin, OHC damage was noted primarily in the basal and middle turns of the cochlea. Most of the damage and scar change seemed to affect the first and second rows of OHCs in the basal and middle turns (Figs. 7H and 7L). In contrast to the cisplatin group, a relatively normal appearance of OHCs was identified in the cochleae of rats treated with EGb 761 + cisplatin (Figs. 7E, 7I, and 7M). IHCs maintained their morphologic integrity throughout the cochlea, even in the presence of cisplatin. However, Fig. 7I image looked a little disorganization of stereocilia of IHCs. It might be from the error of technical procedures or damage from cisplatin. These data offer strong evidence supporting the potential benefits of EGB 761 against cisplatin-induced hearing loss.
- 31 -
Fig.10. Phalloidin staining and SEM of cochlear hair cells.
The control group (C, G, K) and the EGb 761 group (F, J, N) showed normal appearance of stereocilia in every turn of the cochleae in the phalloidin staining (C - F) and SEM (G – N) studies. However, the damage of OHCs was noted primarily in the basal and middle turns of the cochlea in the cisplatin group (D, H, L) but it was prevented by EGb 761 (E, I, M). ABR: Auditory Brainstem Response. Y axis (dB) in figs. A and B means hearing threshold of ABR.
- 32 -
IV. Discussion
The role of GJIC in hearing, for the purpose of understanding the phenomena responsible for ototoxic drug-induced hearing loss, is poorly studied. Gap junctions are channels connecting adjacent cells, composed of Cx hexamers in the plasma membrane. These channels mediate electrical and chemical coupling between cells. GJIC is important in maintaining tissue homeostasis, normal cell growth, differentiation, and development (Loewenstein, 1979). So far, 21 Cx isoforms have been described, of which Cx43 is a widespread short-lived protein (Laird, 2006; Sohl et al., 2004). Cxs are regulated by multiple mechanisms, including mitogen-activated protein kinase (MAPK) dependent phosphorylation (Jo et al., 2005; Lampe et al., 2004; Wang et al., 2000), as well as lysosomal and proteasomal degradation (Leithe et al., 2004; Qin et al., 2003).
Cx mutations, with a carrier frequency of approximately 3%-4% have been established as the most common cause of non-syndromic hearing loss, responsible for up to 20% of cases of childhood deafness (Cohn et al., 1999; Denoyelle et al., 1997; Estivill et al., 1998; Gasparini et al., 2000; Sobe et al., 2000). In one study (Kenna et al., 2001), 30% of 99 probands were detected as GJB2 mutations, which included 18% of biallelic mutations and 12% of single mutations. Another study of pediatric and adult patients with severe-to-profound hearing loss has reported an even higher incidence of GJB2 mutations (Gasparini et al., 2000). The GJB2 gene codes for the gap junction channel protein connexin 26 (Cx26). It has been suggested that gap junctions assist in potassium homeostasis in the cochlea and that mutations in the Cx26 protein impair the endolymph potassium concentrations that are required for auditory signal transduction (Lautermann et al., 1999; Frenz et al., 2000; Xiz et al., 1999). Also, amelioration of gap junction by toxic agents is known to cause abnormal function and cause acute or chronic diseases in tissues and to affect tumor promotion. Since the discovery of gap junction inhibition by various agents has been reported supporting the hypothesis that
- 33 -
gap junction inhibition is the cellular mechanism underlying cellular promotion and that prevention of the down-regulation of gap junction is critical for preventing toxic effect (Yotti et al., 1979; Murray et al., 1979). Thus, even though the main mechanism of cisplatin ototoxicity is related with ROS, we should not overlook the change of Cx function or expression in cisplatin ototoxicity.
The mechanism of cisplatin-induced ototoxicity has been intensively investigated. There is compelling evidence that the formation of ROS, particularly superoxide anion, occurs, leading to depletion of intracellular antioxidants, such as glutathione or superoxide dismutase (SOD), from cochlear tissues (Rybak et al., 2007). This may allow lipid peroxidation, as shown by increased concentrations of malondialdehyde (Rybak et al., 2007), and toxic lipid peroxides and aldehydes, such as 4-hydroxynonenal and peroxynitrite (Lee et al., 2004). These can increase calcium influx and apoptosis in cells of the cochlea (Alam et al., 2000; Clerici et al., 1995; Lee et al., 2004).
One of the enzymes that can produce superoxide radicals is an isoform of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, NOX-3, which is unique to the cochlea. Cisplatin has been shown to activate this enzyme, leading to a dramatic increase in superoxide production (Banfi et al., 2004) not only in cochlear cell lines but also in the cochlea of rats exposed to cisplatin (Mukherjea et al., 2006). Superoxide can lead to the formation of hydrogen peroxide. Superoxide can also react with nitric oxide to form peroxynitrite which reacts with proteins to form nitrotyrosine. Cochlear hair cells of cisplatin-treated guinea pigs demonstrate immunoreactivity for 4-hydroxynonenal, but auditory neurons were found to be immunopositive for both 4-hydroxynonenal and nitrotyrosine (Lee et al., 2004).
ROS can stimulate Bax in the cytosol, which translocates to the mitochondria causing leakage of cytochrome C into the cytosol. This, in turn, can stimulate caspases-9 and -3,
- 34 -
leading to breakdown of DNA by caspase activated deoxyribonuclease (Watanabe et al., 2003) and cleavage of fodrin within the cuticular plate of damaged OHCs by caspase-3, leading to apoptosis (Wang et al., 2004). Furthermore, p53 plays an important role in cisplatin-induced cell death. A study of OC-k3 cells exposed to cisplatin suggested that the MAPK cascade may be involved in induction of cell death. Experiments with cisplatin-exposed cells demonstrated an increased phosphorylation of extracellular regulated kinase (ERK) 1/2 that was inhibited by PD98059 and suramin. Both of these inhibitors protected these cells from cisplatin-induced cytotoxicity. Activated extracellular signal-regulated kinase 1/2 may be a main effecter of cell death, causing nuclear fragmentation, actin cytoskeleton rearrangement, and cell death. Cell death from cisplatin occurred in a p53 and caspase-independent manner, because these cell lines are functionally depleted of p53 (Previati et al., 2007). Cisplatin-induced permanent hearing loss has been associated with the degeneration of cochlear OHCs and cells within the stria vascularis (Comis et al., 1986; Meech et al., 1998). In our study, cisplatin induced the loss of OHCs in the basal and middle turns of the cochlea.
Inflammatory cytokines may mediate cisplatin ototoxicity. These cytokines (tumor necrosis factor-a(TNF-a), interleukin (IL)-1b and IL-6) may be upregulated by the activation of ERK and nuclear factor kB (NFkB). This was demonstrated in HEIOC1 cells in vitro. Neutralization of these cytokines by antibodies and pharmacologic inhibition of ERK significantly reduced the death of these cells exposed to cisplatin. These in-vitro studies were confirmed in rats. TNF-α was immunolocalized to the spiral ligament, spiral limbus and the organ of Corti. This suggests that proinflammatory cytokines, especially TNF-α, play a significant role in cochlear damage caused by cisplatin (So et al., 2007).
Despite the absence of definitive understanding of the mechanism of the link between the functional status of Cxs and regulation of apoptosis in auditory cells,
- 35 -
emerging data suggests that cisplatin can affect the function of GJIC. Previous work, using other connexins, other cell types and othercytotoxic agents, has shown that GJIC can enhance and propagatetoxic effects and apoptosis (Kalvelyte et al., 2003; Krutovskikh et al.,2002; Udawatte and Ripps, 2005; Wang et al., 2007). Xuhui et al. described that heteromeric gap junctions composed of Cx26 and Cx32 in HeLa cells enhance the cytotoxicity of cisplatinas assessed by proliferative capacity, cell survival and induction of specific apoptotic caspases (Xuhui et al., 2013). Reduced GJIC and down-regulated expression of Cx43 have been described in the senescent endothelial cells (Xie et al., 1994) and in the senescent human fibroblasts (Statuto et al., 2002). In addition, a study reported hydrophobic platinum complex showed time- and dose-dependent decrease in GJIC in liver epithelial cells of rats, and this change was linked to rapid hyperphosphorylation of Cx43 and the disappearance of Cx clusters from cell membranes (Procházka et al., 2007). Treatment with cisplatin (10mM) can block cell growth cycle at G1 and induce premature senescence in human fibroblasts (Zhao et al., 2004). The premature senescence changes include high frequency of apoptosis, elevation of p53 expression, and loss of membranous gap junctions and reduction of dye-transfer capacity. Cisplatin and oxaliplatin inhibit the activity of Cx channels by direct interaction with Cx protein and by reducing the level of Cx expression (Wang et al., 2010). In our study, like previous reported results, cisplatin also induced the loss of hair cells in basal and middle turn of cochlea, and down-expression of Cx26, 30, 31, and 43 in HEI-OC1 cells. Despite the uncertainty of the relationship between down-regulation of Cxs and loss of hair cells by cisplatin, we may surmise that ROS generation causes down-expression of Cxs in the supporting cells indirectly resulting in hair cell death. Therefore, the down-regulation of GJIC and inhibition of Cx expression may be one of the important mechanisms in cisplatin-induced ototoxicity.
Although the important roles of gap junctions in the controlling of cellular apoptosis have been suggested by several authors, different kinds of experiments have led to different results in cancer cells (Krutovskikh et al., 2002; Sai et al., 2001; Watanable et
- 36 -
al., 1995; Yasui et al., 2000). Peterson-Roth et al. (2009) reported that cisplatin toxicity is enhanced by the presence of functional gap junctions between the target cells. Tanaka et al. (2001) also described that in bladder cancer lines gap junction expression acts additively with cisplatin in promoting apoptosis, cell cycle arrest, and down-regulation of bcl-2. Therefore, the different regulation of GJIC and Cxs by cisplatin depending on target cells, normal cells or cancer cells requires attention and may justify the local delivery of the EGb 761 to the inner ear.
Protection against cisplatin may be provided by endogenous molecules, by exogenous agents or by a combination of exogenous agents that trigger endogenous protective mechanisms. Endogenous mechanisms, however, do not seem to be able to overcome the toxic effect of cisplatin when the dose exceeds a certain threshold. Therefore, many exogenous protective agents have been studied in experimental and/or clinical research to prevent cisplatin-induced ototoxicity (Rybak et al., 2003; Rybak et al., 2005). Protection against cisplatin ototoxicity has been observed with the intracochlear perfusion of inhibitors of caspase-3 and caspase-9. These agents dramatically reduced the extent of hearing loss and apoptosis of hair cells (Wang et al., 2004). The application of the p53 inhibitor, pifithrin-a, to organotypic organ of Corti cultures also protected the hair cells from damage. The protection was correlated with a reduction in p53 expression and caspase activation (Zhang et al., 2003). Numerous attempts have been made at upstream protection of the cochlea with a variety of antioxidant compounds before death pathways. Among them, several antioxidants containing thiol groups attenuate cisplatin ototoxicity including sodium thiosulfate, diethyldithiocarbamate, D or L-methionine, methylthiobenzoic acid, lipoic acid, N-acetylcysteine, tiopronin, glutathione ester and amifostine (Rybak et al., 2005). Other agents that may function as free radical scavengers and also protect the cochlea from cisplatin damage and hearing loss in experimental animals include a-tocopherol (alone or in combination with tiopronin), aminoguanidine, D and L-methionine, and sodium salicylate (Rybak et al., 2005). Ebselen (alone or in combination with allopurinol, an inhibitor of xanthine oxidase) has also been shown to protect rats against cisplatin
- 37 -
ototoxicity (Lynch et al., 2005).
Therefore, we selected Gingko biloba, a typical antioxidant, which is considered to be the world’s oldest living tree species containing unique chemical substances unknown in other living things. EGb 761, a standardized extract of G. biloba, has potent antioxidant activity (Marcocci et al., 1994). It acts as a free radical scavenger by preventing lipid peroxidation, increasing GSH levels and GR activity, and possessing SOD activity. The flavonoids contained in EGb 761 act mainly on hydroxyl radicals and inhibit the lipid peroxidation of cell membranes. Interestingly, EGb 761 has been shown to protect brain tissue by inhibiting NOS in gerbils (Calapai et al., 2000). EGb 761 also has anti-inflammatory and antiapoptotic properties in normal tissues (Christen et al.,n 2002). The combined activity of all the EGb 761 components is required for optimal therapeutic action (Christen et al.,n 2002). The finding that EGb 761 protected against cisplatin induced ototoxicity was consistent with a Japanese study which showed that a standardized G. biloba extract (100 mg/kg, orally, for 10 d) administered 90 minutes before administration of cisplatin (1 mg/kg x 10 day) could ameliorate drug-induced auditory threshold shifts and protect renal function without any attenuation of the anticancer effect of cisplatin in Fisher rats (Fukaya and kanno, 1999). Xu et al. (2004) reported EGb 761 (14 mg/kg, i.p. 2 days before, for 7 days) could effectively reduce cisplatin-induced ototoxicity (cisplatin at 2 mg/kg, i.p. for 5 days) in guinea pigs, attenuating cisplatin-induced changes in serum SOD activity and malondialdehyde levels. Huang et al. (2007) also reported EGb 761 (200 mg/kg, i.p.) protects against cisplatin-induced ototoxicity (13 mg/kg, i.p.). As well, in the present study, EGb 761 significantly prevented hearing loss in rats injected i.p. with cisplatin. EGb 761, as a free radical scavenger, may play an important role of the protective effect on cisplatin-induced ototoxicity.
Furthermore, EGb 761 could affect the function of GJIC in cisplatin-treated cells. Although there has not been a study of the relationship between EGb 761 and GJIC in