저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
TCF4-mediated Transcriptional Regulation of
Kr
üppel-like Factor 4 Gene
by
Min Woong Kim
Major in Neuroscience
Department of Biomedical Sciences
The Graduate School, Ajou University
TCF4-mediated Transcriptional Regulation of
Kr
üppel-like Factor 4 Gene
by
Min Woong Kim
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements for
the Degree of Master in NeuroScience
Supervised by
Myung Ae Lee Ph.D.
Major in Neuroscience
Department of Biomedical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of Min Woong Kim is approved.
SUPERVISORY COMMITTEE
(Sign)
Myung Ae Lee
(Sign)
Hye Sun Kim
(Sign)
Joon Kyu Lee
The Graduate School, Ajou University
June 22nd, 2012
ACKNOWLEDGEMENTS
2
년 간의 석사과정 동안 자상한 조언과 세심한 지도로 저를
이끌어주신 이명애 교수님께 진심으로 감사 드립니다. 아울러 많이
부족하고 모자란 논문이지만 심사에 힘써주신 김혜선 교수님과 이준규
교수님께도 깊은 감사 드립니다.
처음 이 자리에 왔을 때부터 실험실 생활에 대해서 배우고 빨리 익숙해
질 수 있도록 옆에서 든든한 후원자가 되어주신 정용이형, 다정하게
대해주신 현숙 박사님, 같이 실험하면서 많이 정이 들었던 정아, 낯선
한국에 와서 꿋꿋하게 실험하는 Rukhsana, 말 잘 듣고 솔선수범하는
막내 정미 이들이 있어서 대학원 생활을 알차게 유지할 수 있었던
계기이고 원동력이었습니다.
밋밋할 수 있는 실험실 생활에 활력소가 되어주었던 6 층 큰형 동훈이형,
준영이형, 웃음 띈 얼굴이 매력적인 환구형, 항상 열심히 하는 동건이형,
든든한 동주형, 목소리 큰 우영이형, 같이 있어서 즐거웠던 동갑내기
친구들 현정이, 상필이, 진우, 기령이, 정 많은 동생들 두현이, 민정이,
주연이, 기방이 혜영이, 진하 모두모두 고맙다는 말씀 드립니다.
마지막으로 큰 손자 잘 되기를 걱정해주시는 할머니, 뒤에서 묵묵히
응원해주시는 아버지, 항상 파이팅 외쳐주시는 어머니, 형 말 잘 따르는
동생 민혁이 우리 가족에게 이 논문을 빌어 다시 한 번 감사의 말씀
전합니다.
i
- ABSTRACT -
TCF4-mediated Transcriptional Regulation of Krüppel-like Factor
4 Gene
Krüppel-like factor 4 (KLF4) is essential for somatic cell reprogramming. In addition, KLF4 is expressed in a variety of tissues and plays an important role in many physiological processes, including proliferation, terminal differentiation and apoptosis. The function of KLF4 has been thoroughly investigated in normal homeostasis, cell differentiation, and cancer formation. In an effort to reveal transcriptional determinants of this tissue specificity, we recognized 2 putative conserved AT-/TA-CAAAG motifs known to bind transcription factor 4 (TCF4). Transcription factor 7-like 2 also known as TCF7L2 or TCF4 is a protein acting as a transcription factor. In the adult intestine, TCF4 is the primary TCF/LEF family member involved in mediating active Wnt/β-catenin signaling and expressed in the colon epithelium. To investigate if activation of klf4 gene mediated by TCF4, we performed western blotting of TCF4 expression in HCT116 and HEK293 cells. TCF4 proteins are expressed in HCT116 cells, but not HEK293 cells. To determine TCF4-binding activity to 2 putative conserved regions present in human KLF4 promoter, we performed electrophoretic mobility shift assay. In mobility shift assay, TCF4 binding complex bands were observed in ATCAAAG motif, and were not observed in TACAAAG motif. These results indicated that TCF4 might regulate only one motif which is distal region of human KLF4 promoter. Taken together, these data suggests that TCF4 protein plays a critical role in the control of
ii
transcriptional regulation of KLF4 gene.
Key words : Krüppel-like factor 4, TCF4, transcription, HCT116, electrophoretic mobility shift assay
iii
TABLE OF CONTENTS
ABSTRACT ··· ⅰ TABLE OF CONTENTS ··· ⅲ LIST OF FITURES ··· Ⅴ Ⅰ. INTRODUCTION ··· 1Ⅱ. MATERIAL AND METHODS ··· 3
A. Cell culture ··· 3
B. Plaque hybridization ··· 3
C. DNA cloning ··· 4
D. Western blot analysis ··· 4
E. Electrophoretic mobility shift assay (EMSA) ··· 5
Ⅲ. RESULTS ··· 7
A. The transcription factors associated with KLF4 ··· 7
B. KLF4 structure schematic diagram ··· 9
C. Identification of putative conserved motif in the 5′-regulating region of the KLF4 gene ··· 11
iv
D. KLF4 sub-cloned into pGL3-basic vector ··· 13
E. TCF4 protein existed in HCT116 cell, but not HEK293. ··· 15
F. TCF4 protein binds to distal region of KLF4 gene in HCT116 cell. ··· 17
G. Proximal region of KLF4 gene is not activated by TCF4. ··· 19
Ⅳ. DISCUSSION ··· 17
Ⅴ. CONCLUSION ··· 19
REFERENCES ··· 20
v
LIST OF FIGURES
Fig.1. The transcription factors associated with KLF4 ··· 8
Fig.2. KLF4 structure schematic design ··· 10
Fig.3. Identification of putative conserved motif in the 5′-regulating region of the KLF4 gene ··· 12
Fig.4. KLF4 sub-cloned into pGL3-basic vector for luciferase assay ··· 14
Fig.5 TCF4 protein existed in HCT116 cell, but not HEK293. ··· 15
Fig.6 TCF4 protein bind to distal region of KLF4 gene in HCT116 cell. ··· 17
1
Ⅰ
. INTRODUCTION
The Krüppel-like family of zinc-finger transcription factors (KLFs) regulates a range of biological processes, including cell growth, differentiation, embryogenesis and tumorigenesis (Suske et al., 2005; Black et al., 2001). A member of this family, KLF4, also known as gut-enriched Krüppel-like factor/GKLF or epithelial/endothelial zinc finger/EZF, is highly expressed in epithelial cells of different tissues (Garrett-Sinha et al., 1996). Krüppel-like factor 4 (KLF4) is a transcription factor highly expressed in the adult intestine (Shields et al., 1996) and is critical in the process of differentiation (Katz et al., 2002).
KLF4, previously known as gut-enriched Krüppel-like factor or epithelial zinc-finger protein, is a member of the Krüppel-like transcription factor family (Bieker, 2001). Like all Krüppel-like factors, KLF4 has three zinc-finger domains in its C terminus. Depending on the cellular context, KLF4 expression can lead to either transcriptional activation or repression (Ghaleb et al., 2005). KLF4 is expressed in the epithelial cells of the gastrointestinal tract, skin, and several other organs, and its expression is associated with terminal differentiation (Shie et al., 2000; Yoon, 2004). KLF4-null mice die within 15 h of birth, presumably due to severe defects in the late-stage differentiation of skin (Segre et al., 1999). Moreover, KLF4-null mice demonstrate a markedly reduced number of goblet cells in the colon (Katz et al., 2002).
Many transcription factors are involved in ES cell self-renewal and pluripotency regulation. Among them, OCT4, SOX2, and NANOG are thought to be the master regulators
2
of ES cell pluripotency. All three factors have been well studied. OCT4 is a POU homeodomain transcription factor that has been established to be essential in vivo and in
vitro for regulation of early embryonic differentiation and maintenance of pluripotency
(Nichols et al., 1998; Niwa et al., 2000). KLF4 is expressed in a variety of tissues and plays an important role in many physiological processes, including proliferation, terminal differentiation, and apoptosis. Depending on the gene targeted, KLF4 can either activate or repress transcription and, in certain cellular contexts, it can function as either an oncogene or a tumor suppressor (Rowland et al., 2005). The function of KLF4 has been thoroughly investigated in normal homeostasis, cell differentiation, and cancer formation. However, the role of KLF4 in ES cell self-renewal and pluripotency had been neglected until recent studies investigating reprogramming of somatic cells into pluripotent cells highlighted the critical role of KLF4 in remodeling cell fate (Takahashi, 2006; Yamanaka, 2007).
Here we show that Krüppel-like factor 4 (KLF4) expression is transcriptionally activated by TCF4 through distal putative region in HCT116 cell but is not influenced proximal putative region. We report the identification of a functional regulatory DNA site in the promoter of the human KLF4 gene and describe the potential importance of this site in the regulation of the expression of KLF4 gene.
3
Ⅱ. MATERIAL AND METHODS
A. Cell culture
Human embryonic kidney 293 (HEK293) cell line were grown in Dulbecco’s modified Eagle’s medium (DMEM, GIBCO) containing 10% fetal bovine serum(FBS, Hyclone), 10ug/ml penicillin-streptomycin (GIBCO, NY, USA) and human colorectal carcinoma (HCT116) cell line were grown in Rosewel park memorial institute medium 1640 (RPMI Medium 1640, GIBCO) with 10% fetal bovine serum(FBS, Hyclone), 10ug/ml penicillin-streptomycin (GIBCO, NY, USA). These cell lines incubated at 37oC in an incubator of 5%
CO2.
B. Plaque hybridization
The basic procedure has been described elsewhere (Sambrookand Russel, 2001). Hybridization was carried out overnight, at 42oC, in 5 × SSC (1 × SSC was 150 mM NaCl and 15 mM Na-citrate), 0.1% BLOTTO and 50% formamide. Washing was performed in 2 x SSC at 42oC for 30 min, and then in 0.1 x SSC, 0.1% (w/v) SDS at 68oC for 30 min. Plaque hybridization was performed using replica nitrocellulose filters from plates containing 2 × 104 clones each from the human genomic λ EMBL library, using approximately 5 × 108 dpm of the probe, as prepared above. The genomic clones selected in the primary screening were subjected to secondary screening. Inserts from purified positive λ clones were subcloned into pBluescript, and their nucleotide sequence determined.
4
C. Identification of putative conserved motif in the 5′-regulating region of the KLF4 gene
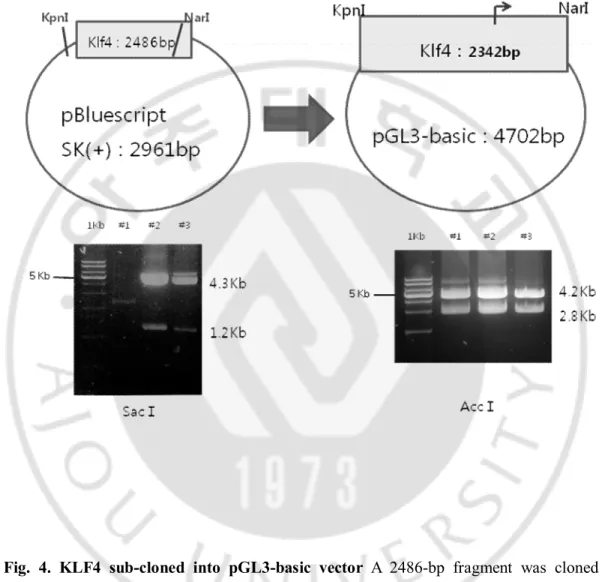
A 2486-bp fragment was cloned in pBluescript SK(+) plasmid vector with SacⅠenzyme in 37℃ incubator for 2 hours. Using enzyme KpnⅠand NarⅠ, a 2342-bp fragment containing sequence – 2221 to +121 was subcloned with ligation on 16℃ incubation for 16 hours for luciferase assay.
D. Western blot analysis
Cells were washed with phosphate-buffered saline (PBS), and directly lysed in RIPA buffer (0.5% sodiumdeoxycholate, 0.1% sodiumdodecyl sulfate, 1% NP-40 and PBS) with PMSF and proteases inhibitors. Protein concentration was determined by Bio-Rad DC assay (Bio-Rad, Hercules, CA, USA). The samples were adjusted to equal protein concentrations, 30㎍ of each protein sample was separated on an SDS-PAGE gel. The protein samples were transferred to immobilon-P membrane (Millipore Corporation, MA, USA). The membrane was incubated in 5% skim milk in TBS-T for 30min at room temperature followed washed three times with TBS-T for 5 min each. Then incubated with TCF4 (1:1000 dilution, Millipore) primary antibody in 5% BSA-TBS-T at 4℃ for overnight. The membrane was then washed three times in TBS-T, 15 min each, and incubated with the secondary anti-rabbit antibody, 1:2000 dilutions (Zymed, CA, USA) for 1 h at room temperature. The membrane was washed three times in TBS-T, and antibody-bound protein was detected by adding ECL Western Blotting Detection reagent (Amersham Pharmacia Biotechnology, buckinghamahire, UK) for 1 min and exposing the membrane to Kodak X-ray film.
5
E. Electrophoretic mobility shift assay (EMSA)
Nuclear extracts from HCT116 and HEK293 cells were prepared essentially as described previously (R.G. Roeder et al., 1983), and protein concentrations were determined using the Bio-Rad Dc protein kit (Bio-Rad). Double-stranded oligonucleotides containing two tandem copies of the KLF4-TCF4 putative binding regions were synthesize, and end-labeled using the T4 Polynucleotide Kinase in the presence of [r-32P] ATP. Free nucleotides were
separated by centrifugation through a G-50 column. DNA-protein binding reactions were carried out in 20㎕ final volume of reaction buffer containing 10mM Tris (pH 7.6), 50mM NaCl, 0.5mM dithiothreitol, 0.5mM EDTA, 1mM MgCl2, 5% glycerol and 250㎍ of poly
(dI-dC) per ml. The nuclear extract (10㎍ of protein) was added to the reaction buffer in the absence or presence of unlabeled competitor DNA and pre-incubated for 15 min on ice. Radioisotope – labeled probe (40,000 cpm) was then added and the mixture was incubated for a further 30 min at room temperature. Electrophoresis to resolve DNA-protein complexes were performed in 4% non-denaturing polyacrylamide gel in 0.25X Tris-borate-EDTA buffer at 200V for 1.5h. The oligolucleotides used were (only sense strand shown) ;
Distal region 5′-TTAGTCATCAAAGGTTTAC-3′ Proximal region 5′-TTTGTATACAAAGGAACTT-3′
Sense and antisense oligonucleotides were annealed and end-labeled with [r-32P] ATP and
T4 polynucleotide kinase according to standard protocol (Schurer, 2002). All of labeled probes were purified on 19% non-denaturing polyacrylamide gel. In standard reactions, proteins and labeled DNA were incubated in a total 20㎕ volumes. Reaction mixtures were
6
pre-incubated for 15min on ice prior to adding labeled DNA and incubated for 30min at room temperature, then loaded onto a 5% non-denaturing polyacrylamide gel. Gels were Fixed, dried and visualized by autoradiography.
7
Ⅲ
. RESULTS
A. The transcription factors associated with klf4
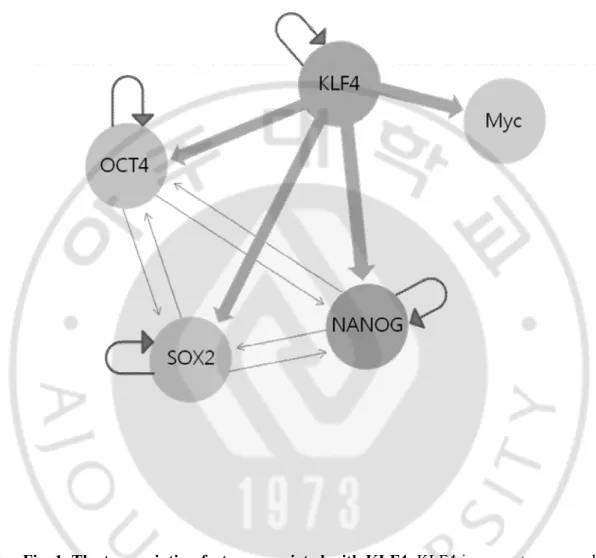
POU5F1/OCT4, SOX2, MYC/c-Myc and KLF4 are the four Yamanaka factors. genes bound by Myc and Rex1 were distinct from those bound by the other factors (Kim et al., 2008), with Myc-association correlating more highly with expressed genes, suggesting that Myc regulation differs from that of the other pluripotency factors. This work also proposes a hierarchy of factors, where KLF4 is an upstream regulator of larger feed-forward loops containing OCT4 and SOX2, a regulator of common targets including NANOG and a potential regulator of Myc, as it binds the c-Myc promoter (Fig. 1).
8
Fig. 1. The transcription factors associated with KLF4. KLF4 is an upstream regulator of larger feed-forward loops containing OCT4 and SOX2, a regulator of common targets including NANOG and a potential regulator of Myc, as it binds the c-Myc promoter. Proposed hierarchy of key pluripotency network transcription factors showing auto-regulatory and cross-auto-regulatory transcriptional loops (arrows).
9
B. KLF4 structure schematic diagram
The two exons of the KLF4 gene are identified by Roman numerals. The translated region or open reading frame is depicted in blue (Fig. 2). A distinct feature of the KLF4 gene is the highly GC-rich nature of the sequence proximal its 5'-end. Thus, the G+C content of the 1000 nt 5'-flanking region is 67% and that of the 5'-untranslated region is 63%. Moreover, the bulk of the GC residues are concentrated in the region between nt -600 and +300 of the gene where the G+C content is 82%. The gene has two exons, each containing a portion of the translated region. The open-reading frame of the KLF4 gene encodes a polypeptide of 513 amino acids with a predicted molecular weight of 53 kDa.
10
Fig. 2. Human KLF4 structure schematic diagram The translated region or open reading frame is depicted in blue. A distinct feature of the KLF4 gene is the highly GC-rich nature of the sequence proximal its 5'-end. The gene has two exons, each containing a portion of the translated region. The open-reading frame of the Klf4 gene encodes a polypeptide of 513 amino acids.
11
C. Identification of putative conserved motif in the 5′-regulating region of the KLF4 gene
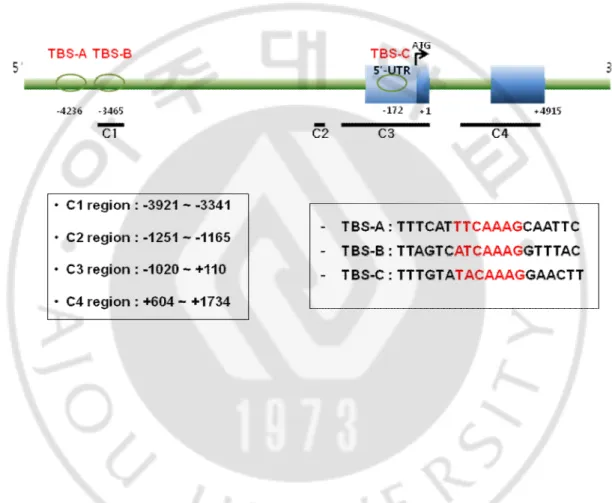
KLF4 is expressed prominently in the differentiating compartment of the gut. In the intestine, KLF4 is expressed in the differentiating cells of the epidermis. This is a key enzyme in the gut and highly expressed in the several cell groups in the intestine and its expression is increased throughout the intestine development, but there is little information on its transcriptional regulation mechanism. To investigate the molecular mechanism by which its expression is tissue-specifically regulated, we have previously characterized the promoter of the human KLF4 gene and reported that transcription factor TCF4 and TCF4 binding regions are regulated for the promoter activity of KLF4 gene.
We identified two motifs of 21 nucleotides with considerable homology to the consensus sequence of the distal and proximal putative region (Fig. 3). This sequence is found in the regulatory region of CAAAG (Hatzis et al., 2008).
12
Fig. 3. Identification of putative conserved motif in the 5′-regulating region of the KLF4 gene This sequence is found in the regulatory region of CAAAG. Distal region is ATCAAG, and proximal region is TACAAAG.
13
D. KLF4 sub-cloned into pGL3-basic vector
We confirmed size of plasmid vector and KLF4 fragment with restriction enzyme (Fig. 4). A 2486-bp fragment was cloned in pBluescript SK(+) plasmid vector with SacⅠenzyme in 37℃ incubator for 2 hours. Using enzyme KpnⅠand NarⅠ, a 2342-bp fragment containing sequence – 2221 to +121 was subcloned with ligation on 16℃ incubation for 16 hours for luciferase assay.
14
Fig. 4. KLF4 sub-cloned into pGL3-basic vector A 2486-bp fragment was cloned in pBluescript SK(+) plasmid vector with SacⅠenzyme in 37℃ incubator for 2 hours. Using enzyme KpnⅠand NarⅠ, a 2342-bp fragment containing sequence – 2221 to +121 was subcloned with ligation on 16℃ incubation for 16 hours for luciferase assay.
15
E. TCF4 protein existed in HCT116 cell, but not HEK293.
To confirm TCF4 expression, we carried out Western blot analysis. We observed TCF4 protein level inside of cells. Western blot result showed that TCF4 existed in HCT116 cell. In contrast, TCF4 is not contained in HEK293 cell ( Fig. 5).
Fig. 5. TCF4 protein existed in HCT116 cell, but not HEK293. Nuclear extracts (30㎍) were prepared from HEK293 and HCT116 cells. Proteins were adjusted as equal amount and performed western blotting.
16
F. TCF4 protein binds to distal region of KLF4 gene in HCT116 cell.
To study whether the human KLF4 sequences interact with TCF4, we performed the electrophoretic mobility shift assays with nuclear extracts from HCT116 and HEK293 cells using the double-stranded oligonucleotides proves containing two putative binding sequences of KLF4 promoter or consensus sequence of the TCF4. We performed competition assay with 20- or 40- or 100-fold molar excess of unlabeled cold KLF4 probes. In distal region, binding was gradually competed by adding cold probes in HCT116 but not by unrelated HEK293 (Fig. 6). And shown as above, the upper complex was specifically disappeared by anti-TCF4 antibody, suggesting that the upper band is KLF4-TCF4 binding complex.
17
Fig. 6. TCF4 protein existed in HCT116 cell, but not HEK293. Incubation of nuclear extracts of HCT116 and HEK293 cells with 32P-labeled KLF4 proves derived from labeled
probe and distal region of KLF4 promoter. For competition assay, an excess of unlabeled oligonucleotides was added at 20X, 40X, 100X molar ratio. Supershift analysis was performed in the presence of anti-Tcf4 antibody.
18
G. Proximal region of KLF4 gene is not activated by TCF4.
We performed competition assay with 20- or 40- or 100-fold molar excess of unlabeled cold KLF4 probes. In proximal region, binding was not exactly completed by adding cold probes in HCT116 (Fig. 7). And shown as above, the upper complex was not specifically disappeared by anti-TCF4 antibody. This result show that TCF4 may not bind to proximal region of KLF4.
19
Fig. 7. TCF4 protein existed in HCT116 cell, but not HEK293. Incubation of nuclear extracts of HCT116 and HEK293 cells with 32P-labeled KLF4 proves derived from labeled
probe and distal region of KLF4 promoter. For competition assay, an excess of unlabeled oligonucleotides was added at 20X, 40X, 100X molar ratio. Supershift analysis was performed in the presence of anti-Tcf4 antibody.
20
Ⅳ
. DISCUSSIN
Krüppel-like factor (KLF)-4 is a C2H2 zinc-finger containing transcription factor that is highly expressed in the gastrointestinal tract (Shields et al., 1996; Ghaleb et al., 2007). It can both activate and repress transcription of different promoters (Ai et al., 2004). It has been proposed to function as a tumor suppressor and an oncoprotein, depending on cellular context. For example, overexpression of KLF4 has been linked to reduced tumorigenicity of colonic and gastric cancer cells in vivo. In addition, specific ablation of KLF4 in the gastric epithelium of mice results in premalignant changes, suggesting that it may be a tumor suppressor (Katz et al., 2005). On the other hand, KLF4 mRNA and protein are over expressed in most squamous-cell carcinomas of the oropharynx and in up to 70% of mammary carcinomas. A role for KLF4 as an oncogene has been further supported by the induction of squamous epithelial dysplasia by ectopic KLF4 expression in mice.
The use of four specific transcription factors, c-myc, Sox2, Oct4 and Klf4, has been shown to induce pluripotency in both human and mouse somatic cells (Takahashi and Yamanaka, 2006; Park et al., 2008). These studies reveal that differentiation is reversible, and provide strong foundation for the development of technologies that will enable the generation of patient-derived pluripotent cells. Immunologically matched pluripotent cells are an ideal source for the generation of cells for transplantation, and patient-derived pluripotent cells may also be useful tools for the study of disease states and drug therapies. Recent analysis of an extended OCT4, SOX2, NANOG transcription factor network in mouse ES cells included
21
the OCT4- or NANOG-interacting proteins Rex1, Dax1, Zpf281 and Nac1 (Wang et al., 2006; Kim et al., 2008) and two pluripotency-associated factors c-Myc (Cartwright et al., 2005), and Klf4 (Jiang et al., 2008). This genome-wide screen of promoters bound by each of the factors found that many promoters (_800) were occupied by multiple, common factors. Moreover, promoters bound by few of the factors were generally inactive, whereas those bound by four or more of the factors were more likely to be active in ES cells and repressed on differentiation (Kim et al., 2008).
we observe that KLF4 and TCF4 are expressed in similar patterns within the large intestine. The pattern of immunohistochemical staining that we found for the TCF4 protein in mouse intestine is consistent with in situ hybridization data looking at expression patterns of TCF4 mRNA that have been previously published. The function of TCF4 in intestinal stem cells has been well studied (Korinek et al., 1998). However, its role in differentiated intestinal epithelial cells is not entirely clear. Given that KLF4 generally functions as a differentiation associated factor within the intestine, we speculate that in the absence of active Wnt signaling, KLF4 interacts with TCF4 in order to maintain the differentiated state. Recently, KLF4 has been found to be one of the four factors that regulate stem cell reprogramming. It will be interesting to examine whether KLF4 inhibits Wnt signaling in inducible pluripotent stem cells (iPS cells) or embryonic stem cells.
22
Ⅴ
. CONCLUSION
Recently, KLF4 has been found to be one of the four factors that regulate stem cell reprogramming. KLF4 is normally active in real embryonic stem cells. In addition, KLF4 seems to play a redundant role along with other KLF family proteins in embryonic stem (ES) cell self-renewal. However, how KLF4 regulates ES cell self-renewal and somatic cell reprogramming is still poorly understood. Here we report that KLF4 is required for both ES cell self-renewal and maintenance of pluripotency by TCF4. In this study, we showed that KLF4 interacts with TCF4. We suggest that TCF4 may be preserved Embryonic Stem (ES) Cell Differentiation by Regulating KLF4 Gene Expression.
23
REFERENCES
1. Ai W, Liu Y, Langlois M, Wang TC. Kruppel-like factor 4 (KLF4) represses histidine decarboxylase gene expression through an upstream Sp1 site and downstream gastrin responsive elements. J Biol Chem 279:8684–8693, 2004 Ghaleb AM, Katz JP, Kaestner KH, Du JX, Yang VW. Kruppel-like factor 4 exhibits antiapoptotic activity following gamma-radiation-induced DNA damage. Oncogene 26: 2365–2373, 2007
2. Bourillot, P. Y., Aksoy, I., Schreiber, V., Wianny, F., Schulz, H., Hummel, O., Hubner, N., and Savatier, P. Stem Cells 27, 1760–1771, 2009
3. Boyer LA, Plath K, Zeitlinger J, Brambrink T, Medeiros LA, Lee TI, Levine SS, Wernig M, Tajonar A, Ray MK et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature, 441:349–353, 2006
4. Brambrink T, Hochedlinger K, Bell G, Jaenisch R. ES cells derived from cloned and fertilize blastocysts are transcriptionally and functionally cloned and fertilized blastocysts are transcriptionally and functionally indistinguishable. Proc Natl Acad Sci USA, 103:933–938, 2006
5. Brons IG, Smithers LE, Trotter MW, Rugg-Gunn P, Sun B, Chuva de Sousa Lopes SM, Howlett SK, Clarkson A, Ahrlund-Richter L, Pedersen RA et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature, 448:191–195, 2007
6. Cartwright P, McLean C, Sheppard A, Rivett D, Jones K, Dalton S. LIF/STAT3 controls ES cell self-renewal and pluripotency by a Myc-dependent mechanism. Development, 132:885–896, 2005
7. Chamberlain SJ, Yee D, Magnuson T. Polycomb repressive complex 2 is dispensable for maintenance of embryonic stem cell pluripotency. Stem Cells, 26:1496–1505, 2008
8. Darr H, Mayshar Y, Benvenisty N. Overexpression of NANOG in human ES cells enables feeder-free growth while inducing primitive ectoderm features. Development, 133:1193–1201, 2006
24
9. Evans, P. M., W. Zhang, X. Chen, J. Yang, K. K. Bhakat, and C. Liu. Kruppel-like factor 4 is acetylated by p300 and regulates gene transcription via modulation of histone acetylation. J. Biol. Chem. 282:33994–34002, 2007
10. Foster, K. W., A. R. Frost, P. McKie-Bell, C. Y. Lin, J. A. Engler, W. E. Grizzle, and J. M. Ruppert. Increase of GKLF messenger RNA and protein expression during progression of breast cancer. Cancer Res. 60: 6488–6495, 2000
11. Ghaleb, A. M., B. B. McConnell, M. O. Nandan, J. P. Katz, K. H. Kaestner, and V. W. Yang. Haploinsufficiency of Kruppel-like factor 4 promotes adenomatous polyposis coli dependent intestinal tumorigenesis. Cancer Res. 67:7147–7154, 2007
12. Goldman, P. S., V. K. Tran, and R. H. Goodman. The multifunctional role of the co-activator CBP in transcriptional regulation. Recent Prog. Horm. Res. 52:103–119; discussion 119–120, 1997
13. Graham, T. A., W. K. Clements, D. Kimelman, and W. Xu. The crystal structure of the beta-catenin/ICAT complex reveals the inhibitory mechanism of ICAT. Mol. Cell 10:563–571, 2002 14. Gregorieff, A., and H. Clevers. Wnt signaling in the intestinal epithelium: from endoderm to cancer.
Genes Dev. 19:877–890, 2005
15. Huber, O., R. Korn, J. McLaughlin, M. Ohsugi, B. G. Herrmann, and R. Kemler. Nuclear localization of beta-catenin by interaction with transcription factor LEF-1. Mech. Dev. 59:3–10, 1996
16. Jen, J., S. M. Powell, N. Papadopoulos, K. J. Smith, S. R. Hamilton, B. Vogelstein, and K. W. Kinzler. Molecular determinants of dysplasia in colorectal lesions. Cancer Res. 54:5523–5526, 1994
17. Jiang, J., Chan, Y. S., Loh, Y. H., Cai, J., Tong, G. Q., Lim, C. A., Robson, P., Zhong, S., and Ng, H. H. Nat. Cell Biol. 10, 353–360, 2008
18. Katz, J. P., N. Perreault, B. G. Goldstein, C. S. Lee, P. A. Labosky, V. W. Yang, and K. H. Kaestner. The zinc-finger transcription factor Klf4 is required for terminal differentiation of goblet cells in the
25 colon. Development 129:2619–2628, 2002
19. Kim, M. K., T. J. McGarry, P. O’Broin, J. M. Flatow, A. A. Golden, and J. D. Licht. An integrated genome screen identifies the Wnt signaling pathway as a major target of WT1. Proc. Natl. Acad. Sci. U. S. A. 106:11154–11159, 2009
20. Korinek, V., N. Barker, P. Moerer, E. van Donselaar, G. Huls, P. J. Peters, and H. Clevers. Depletion of epithelial stem-cell compartments in the small intestine of mice lacking Tcf-4. Nat. Genet. 19:379– 383, 1998
21. Kramps, T., O. Peter, E. Brunner, D. Nellen, B. Froesch, S. Chatterjee, M. Murone, S. Zullig, and K. Basler. Wnt/wingless signaling requires BCL9/legless-mediated recruitment of pygopus to the nuclear beta-catenin- TCF complex. Cell 109:47–60, 2002
22. Levy, L., Y. Wei, C. Labalette, Y. Wu, C. A. Renard, M. A. Buendia, and C. Neuveut. Acetylation of beta-catenin by p300 regulates beta-catenin- Tcf4 interaction. Mol. Cell. Biol. 24:3404–3414, 2004 23. Liu, P., M. Wakamiya, M. J. Shea, U. Albrecht, R. R. Behringer, and A. Bradley. Requirement for
Wnt3 in vertebrate axis formation. Nat. Genet. 22:361–365, 1999
24. Ma, H., C. Nguyen, K. S. Lee, and M. Kahn. Differential roles for the coactivators CBP and p300 on TCF/beta-catenin-mediated survivin gene expression. Oncogene 24:3619–3631, 2005
25. Miyabayashi, T., J. L. Teo, M. Yamamoto, M. McMillan, C. Nguyen, and M. Kahn. Wnt/beta-catenin/CBP signaling maintains long-term murine embryonic stem cell pluripotency. Proc. Natl. Acad. Sci. U. S. A. 104:5668–5673, 2007
26. Morin, P. J., A. B. Sparks, V. Korinek, N. Barker, H. Clevers, B. Vogelstein, and K. W. Kinzler. Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science 275:1787–1790, 1997
26 neoplasia in the mouse. Science 247:322–324, 1990
28. Nowak, D. E., B. Tian, and A. R. Brasier. Two-step cross-linking method for identification of NF-kappaB gene network by chromatin immunoprecipitation. Biotechniques 39:715–725, 2005
29. Ohnishi, S., S. Ohnami, F. Laub, K. Aoki, K. Suzuki, Y. Kanai, K. Haga, M. Asaka, F. Ramirez, and T. Yoshida. Downregulation and growth inhibitory effect of epithelial-type Kruppel-like transcription factor KLF4, but not KLF5, in bladder cancer. Biochem. Biophys. Res. Commun. 308:251–256, 2003 30. Poy, F., M. Lepourcelet, R. A. Shivdasani, and M. J. Eck. Structure of a human Tcf4-beta-catenin
complex. Nat. Struct. Biol. 8:1053–1057, 2001
31. Ramalho-Santos M, Yoon S, Matsuzaki Y, Mulligan RC, Melton DA. “Stemness”: transcriptional profiling of embryonic and adult stem cells. Science 2002;298:597–600
32. Roose, J., G. Huls, M. van Beest, P. Moerer, K. van der Horn, R. Goldschmeding, T. Logtenberg, and H. Clevers. Synergy between tumor suppressor APC and the beta-catenin-Tcf4 target Tcf1. Science 285:1923–1926, 1999
33. Shields, J. M., R. J. Christy, and V. W. Yang. Identification and characterization of a gene encoding a gut-enriched Kruppel-like factor expressed during growth arrest. J. Biol. Chem. 271:20009–20017, 1996
34. Takahashi, K., and S. Yamanaka. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126:663–676, 2006
35. Thompson, B., F. Townsley, R. Rosin-Arbesfeld, H. Musisi, and M. Bienz. A new nuclear component of the Wnt signalling pathway. Nat. Cell Biol. 4:367–373, 2002
36. Wolf, D., M. Rodova, E. A. Miska, J. P. Calvet, and T. Kouzarides. Acetylation of beta-catenin by CREB-binding protein (CBP). J. Biol. Chem. 277:25562–25567, 2002
27
37. Yang, Y., B. G. Goldstein, H. H. Chao, and J. P. Katz. KLF4 and KLF5 regulate proliferation, apoptosis and invasion in esophageal cancer cells. Cancer Biol. Ther. 4:1216–1221, 2005
38. Zhang, W., X. Chen, Y. Kato, P. M. Evans, S. Yuan, J. Yang, P. G. Rychahou, V. W. Yang, X. He, B. M. Evers, and C. Liu. Novel cross talk of Kruppel-like factor 4 and beta-catenin regulates normal intestinal homeostasis and tumor repression. Mol. Cell. Biol. 26:2055–2064, 2006
28 -국문요약-
TCF4 단백질에 의한 Krüppel-like factor 4 유전자의 전사 조절
아주대학교 대학원 의생명과학과 김 민 웅 ( 지도교수: 이 명 애 )Krüppel-like factor 4 (KLF4)은 체세포에서 reprogramming 하는 역할에 필수적이다. 또한 KLF4은 다양한 조직에서 발연하고 세포증식, terminal differentiation, 세포사멸과 같은 많은 생리적 과정에서 중요한 역할을 수행한다. 그 기능으로는 일반적 항상성, 세포 분화, 그리고 암 형성에 밀접하게 관여한다. 이런 조직 특이성에 대한 전사적 결정을 드러내도록 노력하기 위해 우리는 transcription factor 4 (TCF4) 단백질이 binding 하는 것으로 알려진 두 개의 AT-/TA-CAAAG conserved motif를 확인하였다. Transcription factor 7-like 2(TCF7L2)로 알려진 TCF4은 transcription factor의 하나이고, 성인의 장에서 Wnt/β-catenin 신호 전달 활동을 중재하고, 결장 표면에서 발현되는 primary TCF/LEF family member이다. KLF4 유전자의 변화활동이 TCF4에 의해서 중재되는지 알아보기 위해, Western blot 실험을 통해 HCT116 세포와 HEK293 세포에서 TCF4 발현을 관찰하였다. TCF4 단백질이 HCT116 세포에서는 발현 되었지만 HEK293 세포에서는 나타나지 않았다. 인간 KLF4 promoter에
29
존재하는 두 개의 conserved 지역에 TCF4 단백질이 binding 하는지 알아보기 위해, electrophoretic mobility shift assay를 수행하였다. ATCAAAG 지역에는 TCF4 binding complex band가 관찰되었지만 TACAAAG 지역에서는 관찰되지 않았다. 이 결과는 TCF4 단백질이 KLF4 promoter 지역의 먼 곳에서 조절할 것이라는 사실을 보여준다. 위의 결과들로 미루어볼 때 TCF4 단백질이 KLF4 유전자에 대한 전사 과정을 조절하는데 있어 중요한 역할을 할 것이라 여겨진다.
핵심어 : Krüppel-like factor 4, TCF4, transcription, HCT116, electrophoretic mobility shift assay