Prevotella nigrescens의 추출물, 지질다당질 및 세균 배양액에 대한 독성의 특성
원광보건대학 임상병리과․원광대학교 의과대학 진단검사의학교실1
김은숙․김신무․조지현1
Characteristics on the Toxicity of Sonicated Bacterial Extracts, Lipopolysaccharides, and Bacterial Growth Supernatants of
Prevotella nigrescens
Kim, Eun Sook., Kim, Shin Moo., Cho, Ji Hyun1
Dept. of Clinical Laboratory Science, Wonkwang Health Science College, Iksan, Korea Dept. of Laboratory Medicine, Wonkang University College of Medicine, Iksan, Korea1
The purpose of this study was to investigate the toxicities and toxic activities of bacterial preparations from Prevotella nigrescens. Supernatants, sonicated bacterial extracts, and lipopolysaccharides(LPS) were used to compare the possible specific sensitivities of human red blood cells(RBCs) toxicity against P.
nigrescens ATCC 33592 isolates. The clinical isolates of P. nigrescens were cultured from infected root canals and identified using polymerase chain reactions of 16S rRNA primers and an ATCC type strain(33592). The toxicity of bacterial growth medium supernatants, sonicated bacterial extracts, and LPSs on RBCs were expressed as a percentage of the positive control(distilled water plus erythrocytes), which was considered to be 100% lysis. The toxicity of RBCs in bacterial growth supernatant detected P. nigrescens ATCC 33592 at 30.5% and P. nigrescens clinical isolate at 27.1%. Sonicated bacterial extracts had concentrated toxicity at dosages of 30㎍/ml, 50㎍/ml, and 100㎍/ml. The LPS fractions had no toxicity.
These results suggest that the characteristics of the toxicity of RBCs against P. nigrescens causes lysis of erythrocytes in sonicated bacterial extract.
Key Words: Prevotella nigrescens, PCR, Toxicity, LPS 임상병리검사과학회지 : 35권 제1호, 14-21, 2003
1)I. 서 론
1697년에 레벤훅크(Leeuwenhoek)는 우식이 심한 치아 의 근관내에서 세균감염 가능성에 대해 처음으로 언급하 였다(Spangberg, 1990). 그로부터 200년이 지나 Miller
교신저자 : 김신무, (우)570-750 전북 익산시 신룡동 344-2, 원광보 건대학 임상병리과
Tel : 063-840-1211 E-mail : [email protected]
(1890)에 이르러서야 현미경을 이용하여 괴사된 치수조직 에서 다양한 형태의 미생물이 존재함을 입증하였다 (Miller, 1890). Henrici와 Hartzell(1919)은 정상적인 치수 는 원래 무균상태이며 근관치료는 이미 존재하고 있는 근관내 감염요소를 모두 제거하여 조직의 재감염을 방지 하는 것이라고 발표하여, 근관치료학에서의 세균의 중요 성을 강조하였을 뿐 아니라 현대적 개념의 근관치료에 대해서도 언급하였다(Henrici와 Hartzell, 1919). 그 이후에
행해진 연구들에서도 치수 및 치근단 질환이 발병되거나 진행될 때 미생물이 중요한 역할을 한다는 것이 강조되 었다(Kakehashi 등, 1965; Griffe 등, 1980; Zavistoski 등, 1980). 최근에는 감염 근관내 세균분포의 변화가 일정한 양상을 보이는 것으로 밝혀졌는데, 즉 감염 초기에는 통 성혐기성 세균인 streptococci나 enterococci 등이 주로 나 타나지만, 치수가 괴사되어 근관이 혐기적 상태로 변해감 에 따라 치근단 부위에서 혐기성 세균이 크게 증가하는 것으로 보고되고 있다(Dahl 등, 1992; Haapasalo, 1993).
1970년대 이전의 연구에서는 그 방법이 부적절하고 배양 기술이 미숙하여 감염 근관내에서 혐기성 세균의 역할이 중요시 되지 않았으나(Baumgartner와 Falker, 1991) 1976 년, Sundqvis가 외상으로 인해 치수괴사가 일어난 전치의 근관내에서 혐기성 세균이 많이 나타나며 이들 혐기성 세균이 근관 감염에 중요한 역할을 한다고 보고한 이후, 혐기성 세균과 이것의 배양 방법, 혐기성 세균과 호기성 세균 및 통성혐기성 세균의 상승 효과에 대한 연구가 활 발히 진행되었다(Sundqvis, 1976). Hashioka 등(1992)은 임 상증상이 없는 경우보다는 있는 경우에 혐기성 세균의 수와 종류가 증가한다고 보고하였다(Hashioka 등, 1992).
진행된 감염근관에서 발견되는 혐기성 세균의 대부분은 그람음성혐기성 간균으로, 혈액한천배지에서 검정색 또 는 갈색의 집락을 형성한다. 그 중에서도 Prevotella intermedia는 치주질환과 연관되어있고, 급성 증상이 있는 근관감염에서도 많이 발견되는 균으로, 최근에는 P.
intermedia에서 다른 DNA구조를 가진 P. nigrescens를 분 리하였다. 1992년 Shah 등은 P. intermedia균주들을 multilocus 효소 전기영동 한 결과 두가지 다른 형태의 전 기영동 프로필 (profile)을 보이며, 하나는 ATCC25611과 같고, 다른 하나는 ATCC33563과 같은 것을 발견하여 ATCC33563과 hybridize되는 유전적인 특징을 가진 균주 들을 P. nigrescens라 명명하였다(Shah와 Gharbia, 1992).
P. nigrescens가 P. intermedia로부터 재분류된 후 근관감 염에서 P. nigrescens는 black-pigmented bacteria 중에서 발 현율이 가장 높게 관찰되었다(Shah와 Gharbia, 1992;
Baumgartner와 Falker, 1991). 그런데 P. intermedia와 P.
nigrescens가 구강내에 동일하게 존재하며 P. intermedia는 치주가 건강한 성인과 아동에서는 발견되지 않고 치주염 이 심화될수록 검출이 증가함으로써 치주질환과 관련되
어 있다고 생각되는 반면 P. nigrescens는 건강한 치주조 직에서 뿐 만 아니라 감염 근관에서 P. intermedia보다 3 배 이상 검출되었다는 보고가 있다(Gharbia 등, 1994;
Matto와 Saarela, 1996).
이러한 Prevotella가 생성하는 용혈소는 또한 다양한 병 인의 중요한 독성인자로 간주된다(Boerlin 등, 1999;
Maslow 등, 1999). 검은색소 생성 혐기성균과 같은 세균 은 성장하기 위해 철이 필요하다. 적혈구의 용혈성 파괴 결과 혈색소와 헤민(hemin)은 구강에서 세균성장의 영양 소인 exogenous iron의 중요한 자원일 수 있다(Leung 등, 1998; Amano 등, 1995).
그 동안 치주질환과 관련된 P. intermedia의 toxicity activity와 용혈소(hemolysin)에 관한 연구는 많이 되어있 으나 근래에 새롭게 분류된 P. nigrescens의 용혈소에 대 한 연구는 거의 없는 편이다. 임상검체에서 분리된 P.
nigrescens의 적혈구에 대한 독성을 알아보고자 본 연구를 시도하였다.
II. 재료 및 방법
1. PCR 검사에 의한 P. nigrescens 특이 16S r-RNA 검출
Qiagene DNeasy kit (Qiagene, Germany)를 이용해서 DNA를 추출하여 P. nigrescens 특이 16S r-RNA gene을 검출하기 위하여 Dahlen 등이 고안한 시발체(sense primer, 5’-GTG TTT CAT TGA CGG CAT CCG ATA TGA AAC-3’ ; antisense primer, 5’-CCA CGT CTC TGT GGG CTG CGA-3’, 828 bp)를 사용하였다.
PCR 산물의 확인은 1.2 % agarose gel electrophoresis를 시행하여 분석하였다
2. 세균배양액
독성 실험을 위한 세균배양액은 BM broth(1% tryptone Difco, Detroit, MI, USA), 1% proteose peptone(Difco), 0.5% yeast extract(Difco), 0.5% NaCl, 5 ㎍/ml hemin, 0.5
㎍/ml menadione) 5 ml 당 1 ml의 현탁액을 넣고 7일간 혐
기적으로 배양하였다. 배양된 균액은 3,000 rpm에서 30분 간 원침한 후 상청액을 준비하였다. 상청액은 여과멸균 (0.22 ㎛)하여 즉시 실험할 때까지 -20℃에 보관하였다.
3. 세균초음파 추출 단백질
세균을 PBS 완충액으로 3회 세척하고 완충액으로 균 부유액을 만들었다. 균 부유액은 열 변성을 방지하기 위 하여 얼음속에 묻어두고 1분씩 파쇄기(BIO-RAD, USA)로 30회 파쇄시켰다. 그 결과 얻어진 상청액을 여과멸균하였 다. 초음파 추출 단백질의 일정농도를 맞추기 위하여 Bradford Protein Assay Kit (Bio-Rad Laboratories, CA, USA)를 이용하여 단백질 농도를 측정하였다. 최종농도를 완충액으로 희석하여 -20℃ 냉동고에 보관하였다.
4. Lipopolysaccharide (LPS) 준비
LPS는 P. nigrescens ATCC 33563과 임상분리균주를 각 각 1 g씩을 Eidhin과 Mouton의 방법에 의해 추출하였다.
LPS추출물을 확인하기위하여 12% acrylamide gel이 포함 된 SDS-PAGE로 확인하였다 (Eidhin 등, 1993).
5. 적혈구의 준비
적혈구는 모두 사람 혈액을 사용하였다.
6. 독성 실험
적혈구는 PBS 완충액으로 세척하였으며 원침 (3,000 rpm, 15분)후 상청액이 투명해질 때까지 계속 세척하였 다. 세척된 혈구는 1% 혈구 부유액을 만들어 사용하였다.
균배양 상청액의 독성 실험은 적혈구 부유액 0.3 ml에 동 량의 균배양 상청액을 넣고 실험하였다. 그 반응액을 잘 혼합한 후 37℃에서 1시간 배양한 뒤 10,000 rpm에서 5분 간 원침 한 후에 유리된 혈색소의 농도를 545 ㎚의 흡광 도로 측정하였다.
세균초음파 추출 단백질과 LPS의 독성을 알아보기 위 해서 적혈구 부유액 0.3 ml에 농도별로 용량을 조절하여 위 실험과정과 동일한 방법으로 측정하였다. 독성효과를
알아보기 위해 적혈구의 용혈정도를 측정하였다. 용혈정 도는 증류수로 완전용혈 된 흡광도를 100%로 하여 각 실 험조건에 따른 용혈정도를 상대적으로 계산하였다. 용혈 에 대한 sodium azide (NaN3)와 dithiothreitol (DTT)의 영 향을 보기 위한 실험에서는 NaN3와 DTT농도가 상청액에 2 mM, 10 mM, 50 mM, 세균초음파 추출 단백질에서는 50 mM이 되도록 조절하여 관찰하였으며, 동일한 조건하 에서 상청액과 초음파 추출 단백질을 넣지 않은 군을 대 조군으로 하였다.
III. 결 과
1. PCR 검사에 의한 P. nigrescens 특이 16S rRNA 의 검출
임상에서 분리된 균주는 828bp 크기의 P. nigrescens 특 이 16S rRNA를 관찰 할 수 있어 P. intermedia가 아니라 P. nigrescens임을 확인 할 수 있었다(Fig. 1).
(bp)
Fig. 1. Agarose gel electrophoresis of the PCR products from the endodontitis of patients. Lane SM, Size marker; Lane 1, PCR product of P. nigrescens ATCC 33563; Lane 2, PCR product of P.
nigrescens clinical isolate.
2. LPS 추출물의 전기영동 확인
P. nigrescens의 LPS를 추출하여 12% acrylamide gel이 포함된 SDS-PAGE를 실시한 결과 Prevotella속의 전형적 인 36 kDa위치에서 ladder모양의 LPS추출물이 관찰되었 다(Fig. 2).
Fig. 2. Silver-stained SDS-PAGE of LPS prepared from P.
nigrescens. S, SDS-PAGE size marker; a, P. nigrescens ATCC 33563; b, P. nigrescens clinical isolate.
3. 세균배양 상청액의 온도변화에 따른 독성비교
세균배양 상청액에 대한 37℃에서의 용혈정도는 증류 수로 완전 용혈시킨 대조군에 비하여 실험군인 표준균주 (P. nigrescens ATCC 33536)는 30.5%, 임상 분리균주(P.
nigrescens)는 27.1% 관찰되었다. 95℃에서의 독성은 증류 수로 완전 용혈시킨 대조군에 비하여 실험군인 표준균주 는 22.7%, 임상 분리균주(P. nigrescens )는 14.7% 관찰되 었다(Fig. 3).
4. 세균초음파 추출 단백질 농도에 따른 독성비교
적혈구 부유액 0.3 ml에 대한 세균초음파 추출 단백질 30 ㎍농도에서 표준균주는 11.4%, 임상 분리균주는 22%
이고, 50 ㎍농도에서 표준균주는 17.4%, 임상 분리균주는
0.0 5.0 10.0 15.0 20.0 25.0 30.0 35.0
37℃ 95℃
bacteria
% of control
ATCC33563 Clinical isolate
Fig. 3. Comparison of toxicity against bacterial growth supernatant of P. nigrescens.
0 20 40 60 80
30㎍ 50㎍ 100㎍
bacteria
% of contro
ATCC33563 Clinical isolate
Fig. 4. Comparison of toxicity against sonicated bacterial extract(SBE) of P. nigrescens.
34.1%이며, 100 ㎍농도에서 표준균주는 46.7% 임상 분리 균주는 71.6%로 나타났다(Fig. 4).
5. LPS 추출물에서 농도에 따른 독성 비교
적혈구 부유액 0.3 ml에 대한 추출된 LPS의 각 농도, 200 ㎍, 400 ㎍, 1000 ㎍농도에서 전부 용혈을 볼 수가 없 었다.
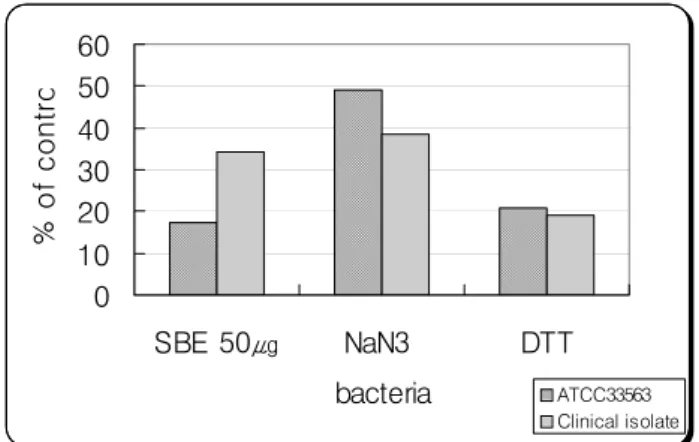
6. NaN3와 DTT의 독성 억제효과
세균초음파 추출 단백질 50 ㎍농도에서 NaN3 억제효과 를 보기위해서 50 mM의 농도를 첨가했을 때 표준균주는 49.1%, 임상 분리균주는 38.6%였고 DTT억제효과를 보기 위해 50 mM의 농도를 첨가했을 때 표준균주는 20.8%, 임
0 10 20 30 40 50 60
SBE 50㎍ NaN3 DTT bacteria
% of contro
ATCC33563 Clinical isolate
Fig. 5. Effect of sodium azide(NaN3) and dithiothreito(DTT) on toxicity against SBE of P. nigrescens.
상 분리균주는 19.2%로 관찰되었다(Fig. 5).
IV. 고 찰
흑색 색소를 형성하는 그람음성 혐기성 세균은 급성 임상 증상을 가진 환자의 근관에서 자주 발견되는 세균 으로서 세균 및 세균의 성분과 산물이 치근단 병소의 생 성과 밀접하게 연관된 것으로 알려져 있다.
Hillman 등은 치주질환을 가진 환자의 3/4이 그들의 치 은연하 플라그에서 용혈성 균주가 1개 또는 그 이상으로 차이가 보인다고 서술하였다(Hillman 등, 1993). 이러한 용혈성균주는 Actinomyces. Streptococcus, Staphylococcus, Prevotella 및 Actinobacillus로 관찰되었으며 이 5균종 중 에 절대혐기성 균주인 Prevotella는 용혈이 적혈구의 종류 에 따라 다르게 나타났다. P. intermedia경우는 토끼와 사 람 적혈구에서 강한 적혈구 응집을 나타내며 면양적혈구 에서는 용혈이 약하게 나타났다(Leung 등, 1989; Dahlen 등, 1996). 이와 같이 Prevotella에 대한 적혈구의 특이성 으로 Prevotella는 토끼 적혈구를 강하게 용혈하였으며 사 람 적혈구에 대한 용혈성은 보통으로 나타나고 면양적혈 구에서는 용혈이 나타나지 않았다(Beem 등, 1998). 본 연 구에서는 Prevotella에 의해서 용혈이 나타나는 사람적혈 구를 사용하여 P. nigrescens의 독성정도를 확인하였다.
용혈은 기능면에서 볼 때 독력인자로 고려 될 수 있다 (Marre 등, 1989; Mobley 등, 1990). 이 독력기능은 상주균
이나 침입하는 세균이 in vivo에서 자라고 대사를 하는데 필요한 철의 획득 가능성을 증가시킴으로써 성취되는 것 같다. 숙주에서 발견 할 수 있는 철의 모두는 세포속에 있고 세포밖에 남아있는 것은 알부민, 합토글로빈, 히모 펙신, 트란스피린 또는 락토페린에 결합하기 때문에 세균 이유효한 철을 획득하기 위한 경쟁은 병변을 유지하는 데 긴요한 것이다(Bullen, 1973; Payne과 Finkelstein, 1978). 따라서 세균의 감염이 일어나려면 침입하는 세균 이 숙주에서 발견되는 매우 낮은 농도의 철을 얻기 위하 여 성공적으로 경쟁할 수 있어야 한다(Payne과 Finkel- stein, 1978). 이 경쟁은 siderophore의 작용으로 일어나거 나 혹은 숙주의 철-결합성 단백에서 철을 떼어 낼 수 있 는 세균이 소유한 철-결합성 단백의 작용을 통하여 일어 난다(Konopka 등, 1982; Bramanti와 Holt, 1990). 용혈소는 감염이 일어나는 국소적 부위에 있는 적혈구를 용해하고 세균이 획득할 수 있도록 헤모글로빈을 유리시킴으로써 철의 농도를 증가시킨다. 세균에게 독력 잠재력을 제공하 는 것은 즉 이용할 수 있는 철(매우 낮은 농도이지만)의 존재인 것이다(Linggood와 Ingram, 1982; Lebek와 Gruenig, 1985). 일반적으로 용혈소는 적혈구 이외에도 다 른 여러 종류의 세포에 작용하여 세포막의 투과성을 변 화시키며, 분비 등의 세포활성에 영향을 미쳐 exocytosis 를 연구하는 데 중요한 도구로 사용되어 왔다(Hilger 등, 1985; Bader 등, 1986; Scheffer 등, 1988).
LPS는 그람음성 세균의 주요 세균벽 성분이며 염증도 발 요소 중 하나로서 그람양성균에서 발견되지 않는다.
치주질환 진행에서 LPS의 역할은 명확하게 규명되지 않 았으나 세균벽의 성분인 LPS보다는 다른 요소인 lipoteichoic acid와 mutein (peptodoglycan)이 주로 뼈(bone) 의 재흡수를 자극하는 것으로 나타났다(Hausmann 등, 1975; Lensgraf 등, 1979; Dewhirst, 1982). 마찬가지로 본 실험에서도 P. nigrescens 세균벽에서 추출한 LPS는 용혈 을 일으키지 않았다. 용혈에 관여된 용혈소는 LPS와는 무관하게 생각할 수도 있으나 강한 세포독성을 일으킨다 는 이전의 논문을 참고하면 LPS를 추출하는 과정에서 95
℃에서의 가열과정에서 LPS성분이 변형을 일으킨 것이 아닌가를 추론 할 수 있다.
세균의 에너지 대사과정에서 전자전달계의 차단제인 NaN3의 용혈에 대한 이전의 논문을 보면 Fusobacterium
nucleatum균주에서 NaN3의 농도가 50 mM까지 증가 할수 록 용혈이 210%까지 증가하는 것을 관찰 할 수 있었다는 보고와 Porphyromonas gingivalis의 용혈성에 있어서 NaN3가 용혈을 감소시킨다는 Chu 등(1991)의 보고가 있 었다. 저자들 연구에서도 F. nucleatum 균주와 같이 증가 함을 볼 수가 있었다.
-SH기는 일반적으로 용혈소를 활성화 시키거나 또는 불활성화 시킨다(Kay 등, 1990; Chu 등, 1991). 본 연구에 서는 -SH기를 갖는 DTT를 이용하여 P. nigrescens 균주의 용혈성을 관찰한 결과 50 mM농도에서 표준균주는 DTT 를 첨가하지 않을 경우와 별 차이가 없었으나 임상 분리 균주의 약 50% 정도 억제효과가 나타난 것으로 보아 -SH 기의 차이가 표준균주와 임상 분리균주가 용혈성능에 차 이가 나타나는 것으로 본다. -SH기에 의하여 활성화되는 이러한 용혈소는 보통 세포막에 있는 콜레스테롤 (cholesterol)과 반응하여 세포를 파괴시키는 것으로 보고 되었고(Haque 등, 1992) 또한 적혈구 세포 표면의 콜레스 테롤이 수용성 용혈소의 수용기로 작용하여 외부로부터 콜레스테롤을 넣어주면 세포 표면의 콜레스테롤과 경쟁 관계가 되어 용혈을 억제 시킨다고 보고 되었다 (Shinoda 등, 1985). 상기 실험들은 주로 P. nigrescens를 가지고 서 로간의 독성의 특성을 알아보고자 실험하였다. P.
nigrescens 균주가 배지에서 성장할 때 용혈소를 세포밖으
로 유리하는지를 알아보기 위해 배양 상청액으로 용혈을 측정한 결과 용혈을 볼 수 있었다. 또한 P. nigrescens 세 균자체가 강하게 용혈성을 나타내므로 그 용혈소가 세포 내 물질과 지질다당체에 있는지를 알아보기 위해 세포분 획을 얻었다. 먼저 표준균주와 임상 분리균주의 지질다당 체를 추출하여 용혈을 측정한 결과 지질다당체에 의한 용혈은 보이지 않았다. 세균초음파 추출 단백질에서 용혈 이 나타나는 것은 용혈소가 단백질이며 다른 단백질들과 마찬가지로 세포질내에서 만들어지며 세포 외막쪽으로 이동하리라는 것을 추정할 수 있다.
Prevotella melaninogenica, P. intermedius, P. oralis와 다 른 그람 음성균들과 같이 서로 가깝게 연관된 혐기성균 들은 protohaem이 절대적으로 요구되는 비슷한 기능을 가 지고 있다(Shah와 Williams, 1987). 그러나 P. gingivalis의 cysteine protease와 같은 유사한 효소는 없고, 없어도 되는 단백분해 활성을 가지는 것으로(Slots 등, 1982; Seddon
등, 1989) 보고되었다.
세균초음파 추출단백질에 의한 용혈이 강하게 나타나 며 배양상청액의 95℃에서 독성효과는 감소하는 것으로 보아 독성의 물질이 단백질임을 추정할 수 있다.
V. 결 론
1. 세균초음파 추출 단백질 농도를 측정한 결과 표준균주 (P. nigrescens ATCC 33536)는 2.52 mg/ml, 임상 분리 균주 (P. nigrescens)는 1.724 mg/ml의 농도이었다.
2. LPS추출 결과 표준균주는 3.3 mg, 임상 분리균주는 3.1 mg이었다.
3. 상청액의 37℃에서의 독성은 표준균주는 30.5%, 임상 분리균주는 27.1%로 나타났다.
4. 상청액의 95℃에서의 독성은 표준균주에서 22.7%, 임 상 분리균주에서 14.7%로 나타났다.
5. 세균초음파 추출 단백질의 독성효과는 농도 30 ㎍에서 표준균주는 11.4%, 임상분리균주는 22%이고, 50 ㎍에 서 표준균주는 17.4%, 임상 분리균주는 34.1%이며, 100㎍에서 표준균주는 46.7% 임상 분리균주는 71.6%
로 나타났다.
6. LPS는 200 ㎍, 400 ㎍, 1000 ㎍의 농도에서 독성은 모 두 볼 수가 없었다.
7. 세균초음파 추출 단백질 50 ㎍농도에서 NaN3 독성억 제효과를 보기위해서 50mM의 농도를 첨가했을 때 표 준균주는 49.1%, 임상분리균주는 38.6%이고 DTT억제 효과를 보기위해 50 mM의 농도를 첨가했을때 표준균 주는 20.8%, 임상 분리균주는 19.2%로 나타났다.
이 연구 결과 세균초음파 추출단백질이 독성을 일으키는 것으로 추론할 수 있으며 독성효과의 차이는 -SH기를 갖는 구조적 차이임을 시사 할 수 있다.
참 고 문 헌
1. Amano A, Kuboniwa M, Kataoka K, Tazaki K, Inoshita E, Nagata H, Tamagawa H, Shizukuishi S. Binding of hemoglobin by Porphyromonas gingivalis. FEMS
Microbiol Lett 134:63-67, 1995
2. Bader M.F, Thierse D, Aunis D. Characterization of hormone and protein release from α-toxin-permeabilized chromaffin cells in primary culture. J Biol Chem 261:
57-77, 1986
3. Baumgartner C, Falker WA. Bacteria in the apical 5 mm of infected root cannals. J Endodon 17:380-383, 1991
4. Beem, JE, Nesbitt WE, Leung, KP. Identification of hemolytic activity In Prevotella intermedia Oral Microbiol Immunol 13:97-105, 1998
5. Boerlin, P, Mc"Ewen, SA, Boerlin-Petzold, F, Wilson, JB, Johnson, RP, Gyles, CL. Associtions between virulence factors of shiga toxin-producing Escherichia coli and disease in humans. J Clin Microbiol 37:497- 503, 1999
6. Bramanti TE, Holt SC. Iron regulate outer membrane proteins in the periodontopathic bacterium, Bacteroides gingivalis. Biochem. biophys. Res Commume 166:
1146-1154, 1990
7. Chu L, Bramanti TE, Ebersole JL, Holt SC. Hemolytic activity in the periodontopathogen Porphyromonas gingivalis: kinetics of enzyme release and localization.
Infect Immun 59:1932-1940, 1991
8. Dahln G, Moller AJR. Microbiology of endodontic infections. p444-475, 1992
9. Dahlen, GG, Johnson JR, Gmur R. Prevotella intermedia and Prevotella nigrescens serotypes, ribotypes and binding characteristics. FEMS Microbiol Lett 138:89-95, 1996
10. Dewhirst FE. N-acetyl muramyl dipeptide stimulation of bone resorption in tissue culture. Infec Immun 35;
133-137, 1982
11. Eidhin DN, Mouton CA. Rapid method for preparation of rough and smooth lipopolysaccharide from Bacter- oides, Porphyromonas and Prevotella FEMS Microbiol Lett 110:133-138, 1993
12. Gharbia SE, Hapaasalo M, Shah HN. Characterization of Prevotella intermedia and Prevotella nigrescens
isolates from periodontic and endodontic infections. J Peridontol 65:56-61, 1994
13. Griffe MB, Patterson SS, Miller CH, Kafrawy AH, Newton CW. The relationship of Bacteroides melanino- genicus to symptoms associated with pulpal necrosis.
Oral Surg 50:457-461, 1980
14. Haapasalo M. Black-pigmented gram-negative anaero- bes in endodontic infections. FEMS Immunol Med Microbiol 6:213-218, 1993
15. Haque A, Sugimoto N, Horiguchi Y, Okabe T, Miyata T, Iwanaga S, Matsuda M. Prodution, purification,and characterization of botulinolysin, a thiol-activated hemo- lysin of Clostridium botulinum Infect Immun 60:71-78, 1992
16. Hashioka K, Yamasaki M, Nakane A, Horiba N, Nakamura H. The relationship between clinical symptoms and anaerobic bacteria from infected root canals. J Endodon 18:558-561, 1992
17. Hausmann E, Luderitz O, Knox K, Weinfeld N.
Structural requirements for bone resorption by endo- toxin and lipoteichoic acid. J Dent Res 54:B94-9, 1975 18. Henrici AT, Hartzell TB. The bacteriology of vital
pulp. Res J 1:419-422, 1919
19. Hilger GA, Bhakdi S, Gratzel M. Minimal requirements for exocytosis: A study using PC12 cells permeabilized with staphylococcal α-toxin. J Biol Chem 260:127-130, 1985
20. Hillman, JD, Maiden, MF, Pfaller, SP, Martin, L, Duncan, MJ, Socransky SS. Characterization of hemo- lytic bacteria in subgingival plaque. J Periodontal Res 28:173-179, 1993
21. Kakehashi SM, Stanley HR, Fitzgerald RJ. The effects of surgical exposures of dental pulps in germ-free and coventional laboratory rats. Oral Surg Oral Med and Oral Pathol 20:340-349, 1965
22. Kay HM, Birss AJ. Smalley JW. Haemagglutinating and haemolytic activity of the extracellular vesicles of Bacteroides gingivalis W50. Oral Microbiol Immunol 5:
269-274, 1990
23. Konopka K, Binderief A, Neilands JB. Aerobactin- mediated utilization of transferrin iron. Biochemistry 21:
6503-6508, 1982
24. Lebek G, Gruenig H. Relation between the hemolytic property and iron metabolism in Escherichia coli. Infect Immun 50:682-686, 1985
25. Lensgraf EJ, Greenblatt JJ, Bawden JW. Effect of group A streptococcal peptidoglycan and group A streptococcal cell wall on bonein tissue culture. Infect Arch Oral Biol 24:495-498, 1979
26. Leung KP, Fukushima H, Sagawa H, Walker CB, Clark WB. Surface appendages, hemagglutination, and adherence to human epithelial cells of Bacteroides intermedius. Oral Microbiol Immunol 4:204-210, 1989 27. Leung KP, Subramanian PS, Okamoto M, Fukushima
H, Lai CH. The binding and utilization of hemoglobin by Prevotella intermedia. FEMS Microbiol Lett 162:
227-233, 1998
28. Linggood M, Ingram PL. The role of alpha-hemolysin in the virulence of Escherichia coli for mice. J Med Microbiol 15:23-30, 1982
29. Marre R, Hacker J, Braun V. The cell-bound hemolysin of serratia marcscens contributes to uropathogencity.
Microb Patholg 7:153-156, 1989
30. Maslow JN, Dawson D, Calrlin, EA, Holland SM.
Hemolysin as a virulence factor for systemic infection with isolates of Mycobacterium avium complex. J Clin Microbiol 37:445-446, 1999
31. Matto J, Saarela M. Distribution and genetic analysis of oral Prevotella intermedia and Prevotella nigrescens.
Oral Microbiol Immunol 11:96-102, 1996
32. Miller WD. Microorganisms of the human mouth. S.S.
White Dental Co Philadelphia 1890
33. Mobley HLT, Green DM, Trifillis AL, Johson DE, Chippendale GR, Lockatell V, Jons BD, Warren JW.
Pyelonephritogenic Escherichia coli and killing of cultured human renal proximal tubular epithelial cells:
role of hemolysin in some strains. Infect Immun 58:
1281-1289, 1990
34. Neilands JB. Iron absorption transport in micro- organisms. Annu Rev Nutr 1:27-46, 1981
35. Payne SM, Finkelstein RY. The critical role of iron in host-bacterial interactions. J Clin Invest 64:1428-1440, 1978
36. Scheffer J, Koning W, Goebel W. Comparison of four hemolysin producing organisms(Escherichia coli, serratia marcscens, Aeromonas hydrophila, and Listeria monocytogenes) for release of inflammatory mediators from various cells. J Clin Microbiol 26:544, 1988 37. Shah HN, Gharbia SE. Biochemical and chemical
studies on strains designated Prevotella intermedia and proposal of a new pigmented species, Prevotella nigrescens sp. nov. Int J Syst Bacteriol 42:542-546, 1992
38. Shah, HN, Williams RAD. Curr Microbiol 15:313-318, 1987
39. Shinoda S, Miyoshi S, Yamanaka H, Nakahara NM.
Some properties of Vibrio vulnificus hemolysin.
Microbiol Immunol 29:583-590, 1985
40. Slots, J, Genco RJ, Mergenhagen SE. In Host-Parasite Interations in Periodontal Diseases(.,eds.) American Society for Microbiology, Washingt- on DC p.27-45, 1982
41. Slots and MA. Taubman. Contemporary oral microbio- logy and Immunology, Mosby-Year book. Inc St Louis 1992
42. Spangberg LSW. Experimental endodontics. In CRC Press INc p132 1990
43. Sundqvis G. Bacterilogical studies of necrotic dental pulps. Ume University Odontology, Dissertation No 7, 1976
44. Zavistoski J, Dzink JBS, Onderdonk A and Bartlett J.
Quantitative bacteriology of endodontic infections. Oral Surg 49:171-174, 1980