96
Copyright © 2015 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
해조류는연안생태계에서광합성을하는일차생산자로서지 구온난화로인한수온상승과해양산성화
(
해수의CO
2농도증 가),
그리고인간활동에따른부영양화등에대한생리적반응 이다르므로환경요인의변화에따라종다양성과군집구조가 변화된다(Connell and Russell, 2010; Nikinmaa, 2013).
인간 의화석연료사용량의증가에따라대기중에CO
2는지속적으로증가되고있으며
,
해양은그중약30%,
즉시간당백만톤(1 million tons)
이상의CO
2를흡수하는것으로알려져있으며(Feely et al., 2004; Gao and Zheng, 2010; Connell and Rus- sell, 2010; Xu and Gao, 2012),
해양의CO
2 농도는현재의380 ppm
에서2050
년에는970 ppm
까지증가될것으로예견하고 있다(Lee et al., 2006).
대부분의해조류는해수의CO
2농도가 증가할때생장과생산량이증가하지만,
조직에CaCO
3을함유 한유절및무절산호조류는가입,
생장및CaCO
3축적에부정해수의 CO2와 영양염 농도 및 조도가 구멍갈파래(Ulva australis) 배아, 유엽과 성체의 생장에 미치는 영향
전다빈·나연주·유옥환
1·최한길*
원광대학교 생명과학부/기초자연과학연구소, 1한국해양과학기술원 해양생태계연구부
Effects of varying CO
2, Nutrient and Light Irradiance Levels on the Growth of Ulva australis at Germling, Juvenile, and Adult Stages
Da Vine Jeon, Yeon Ju Na, Ok Hwan Yu1 and Han Gil Choi*
Faculty of Biological Science and Institute for Basic Science, Wonkwang University, Iksan 570-749, Korea
1Marine Ecosystem Research Division, Korea Institute Ocean Science Technology, Ansan 426-744, Korea
The effects of CO
2concentration, nutrient levels, and irradiance on the growth of germlings and juveniles, and on the photosynthesis of adults were examined in a green tide alga, Ulva australis . We used a factorial experimental design with two CO
2concentrations (380 and 750 ppm), two nutrient levels (control and PES medium), and two irradiance levels (50 and 100 μmol photons m
-2s
-1). Germlings grew best (664.15±61.45 μm in length) under conditions of 750 ppm, PES, and 100 μmol photons m
-2s
-1after 10 days in culture. Relative growth rates (RGR) of the juveniles were greatest (4.41% day
-1) under conditions of 750 ppm, PES, and 50 μmol photons m
-2s
-1after 5 days in culture.
Photosynthetic efficiency (F
v/F
m) of the adult discs was 0.73 ± 0.05 before the experiment and reached a maximum (0.83±0.01) under conditions of 750 ppm, control, and 50 μmol photons m
-2s
-1after 5 days in culture. Growth (germ- lings and juveniles) and photosynthesis (adult discs) of Ulva australis increased when CO
2levels were 750 ppm.
Additionally, the optimal irradiance for growth and photosynthesis differed among stages, wherein germlings grew best at 100 μmol photons m
-2s
-1, juveniles grew best at 50 μmol photons m
-2s
-1, and adults photosynthesized most at 50 μmol photons m
-2s
-1. The performance of Ulva australis at all examined life stages was enhanced under the PES nutrient treatment. In conclusion, the physiological responses of U. australis to varying CO
2, nutrient, and irradiance levels differed slightly among life stages. However, growth and photosynthesis always increased with elevated CO
2and nutrient concentrations. These results indicate that U. australis green tide blooms might occur more frequently in coastal areas if CO
2and nutrient concentrations increase.
Key words: CO
2, Growth, Irradiance, Nutrient, Photosynthesis, Ulva australis
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/)which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2015.0096 Kor J Fish Aquat Sci 48(1) 096-103, February 2015
Received 27 Januaty 2015; Revised 6 February 2015; Accepted 10 February 2015
*Corresponding author: Tel: +82. 63. 850. 6579 Fax: +82. 63. 857. 8837 E-mail address: [email protected]
해수의 CO2, 영양염 농도 및 조도와 구멍갈파래 생장
97
적인반응을보였다
(Kuffner et al., 2007; Hall-Spencer et al., 2008; Martin and Gattuso, 2009; Connell and Russell, 2010;
Ragazzola et al., 2012).
또한
,
최근에우리나라를포함한전세계연안에서빈번하게 일어나는구멍갈파래류,
염주말류,
그리고대마디말류와같은 녹조류대발생(green tide)
은 인간활동으로인한부영양해역 이나교란된해역에서주로관찰된다(Nelson et al., 2003; Hi- raoka et al., 2004; Pinedo et al., 2007; Liu et al., 2009; Yabe et al., 2009; Kim et al., 2011; Deng et al., 2012).
녹조대발생은 해조류군집구조를단순화시키고종다양성을위협하고연안 생태계의변화를교란시키는나쁜영향을준다(Carlton, 2000;
Taylor et al., 2001; Occhipinti-Ambrogi and Savini, 2003).
녹 조류구멍갈파래(Ulva australis)
는영양염이풍부한연안에서 빠른생장을보이는기회종이며,
이들의대발생은관광산업피 폐와대기와해양사이에생물적인장벽을만들어광투과및산 소공급이억제되어다른해양생물의생장과생식에영향을준 다(Taylor et al., 2001; Robic et al., 2009; Nelson et al., 2010;
Renjun et al., 2013; Park, 2014).
우리나라연안에서구멍갈파래는암반에서식하는대표적인 종으로서 연안생태계의생산자이며 다양한해양생물의 먹이 장과생육장으로서중요한역할을하는것으로알려져있지만
(Han et al., 2003),
이들의대발생은연안생태계및관광산업에 나쁜영향을주고있다(Park, 2014).
구멍갈파래류의대발생의 원인은인간활동에의한연안역의부영양화로생각하고있으 나,
해양의CO
2 증가와부영양화의상호작용이구멍갈파래의 대발생과관련이있는지에대한연구는수행되지않았다.
구멍 갈파래는유주자(zoospore),
배아(germling),
유엽(juvenile)
과 성체(adult)
단계의생활사를가진다.
중금속과자외선(UV)
에 노출된구멍갈파래의생리적반응에대한연구(Han and Choi, 2005; Han et al., 2008)
와다양한환경요인(
온도,
광도와영양 염등)
에대한연구가수행되었다(Floreto et al. 1994).
구멍갈파 래에대한선행연구는대부분성체단계에서수행되었으며,
배 아와유엽단계에관한연구는미흡한상태이다(Coelho et al., 2000).
하지만,
해조류의배아와유엽은성체에비해환경변화 에훨씬민감하게반응하므로(Coelho et al., 2000), CO
2 농도증 가와영양염등의환경요인에대한생활사별생장반응을확인 하는것은녹조대발생을이해및관리하는데매우중요한정보 라고할수있다(Choi et al., 2010; Coelho et al., 2000; Deng et al., 2011; Olischläger et al., 2012).
따라서,
본연구는생태학적 으로중요한역할을하지만,
녹조대발생의원인종의하나인구 멍갈파래의배아,
유엽과성체단계에서의CO
2 증가와영양염 에대한반응을확인하기위하여수행되었다.
재료 및 방법
본연구를위한구멍갈파래는전남완도군정도리
(34° 18'N, 126° 41'E)
에서2013
년11
월2
일에채집하였으며,
현장해수와함께플라스틱샘플병에담아서실험실로운반하였다
.
운반된 엽체에착생한동·
식물과미세조류를제거하고멸균해수와유 화용붓을사용하여수회세척한후실험에사용하였다. 배아와 유엽 생장
배아및유엽의생장실험을위하여
,
세척한성숙한엽체를멸 균해수가담긴플라스틱바트에넣어20℃
의연속광으로설정 된배양기에서유주자를방출받았다.
유주자액(4 mL)
은슬라 이드글라스 조각(2.5×2.5 cm)
과 멸균해수가들어있는Petri dish (Ø 12 cm)
에주사기로고르게접종한후유주자가착생 하도록24
시간실온에보관하였다(Fig. 1A).
다음날배아가부 착된슬라이드조각3
개를배양액50 mL
이담긴밀폐된배양 용기에넣고다양한CO
2 농도(380, 750 ppm),
영양염(Control, PES)
과조도(50, 100 μmol photons m
-2s
-1)
가조합된조건에서10
일간배양하였고배양액은2
일간격으로전량교체하였다.
해수의CO
2농도는Gas Concentration Controller (CO
2조절 장치)
로조절하였고,
대조구(control)
는 멸균해수(Autoclaved seawater)
이고PES
는Provasoli's
의 배양액(Provasoli, 1968)
이다.
배양기의광원은형광등을사용하였고조도는100 μmol photons m
-2s
-1이었으며, 50 μmol photons m
-2s
-1는 배양용기 에 검은색 망을 씌워 확보하였으며 디지털조도계(DX-200, TAKEMURA)
로확인하였다.
이때배양기의온도(20℃)
와광 주기는12:12 h L:D (Light: Dark)
로일정하게유지하였다.
배 양10
일후배아는광학현미경(BX53F, OLYMPUS, JAPAN)
에부착된디지털카메라로촬영하였고Image J
프로그램으로 길이를측정하였다.
구멍갈파래 배아는 실내배양에서 유엽으로 생장하였으
며
,
실험에사용된유엽 길이는4.05±0.20 mm
였고면적은0.28±0.02 cm
2(mean±SE, n=180)
였다(Fig. 1B).
구멍갈파 래유엽의실내배양은배아와동일한CO
2 농도,
영양염농도와 조도에서수행되었다.
마개가달린각배양용기(10 mL)
에유엽5
개와배양액을넣고밀폐시켜인큐베이터에서5
일동안배양 하였고배양액은2
일간격으로전량교체하였다.
배양개시5
일 후에유엽을디지털카메라로촬영하여Image J
프로그램에서 길이를측정한후,
상대생장률(RGR, relative growth rate)
을아 래의식으로계산하였다(Ichiki et al., 2000).
RGR (% day
-1) = 100 (lnL
2- lnL
1)/T
2– T
1L
1,
실험개시때의유엽길이; L
2는측정시유엽길이, T
1, T
2 는배양일수성체 광합성효율
해수의
CO
2와영양염농도,
그리고조도가구멍갈파래성체 의광합성에미치는영향을파악하기위하여,
채집한건강한엽 체를멸균해수로수회세척한후, cork borer
로원형디스크(Ø
1 cm)
를만들어24
시간정치배양한후,
탈색되지않은디스크를실험에사용하였다
(Fig. 1C, 1D).
배양기의광주기(12:12 h
LD)
와온도(20℃)
는동일하게세팅하였고밀폐된배양용기에CO
2 농도(380, 750 ppm)
와영양염농도(Control, PES)
가다른 배양액50 mL
과원형디스크5
개씩을넣은후, 50
과100 μmol photons m
-2s
-1의조도에서5
일동안배양하였고배양액은2
일 간격으로전량교체하였다.
성체 디스크의 광합성효율
(F
m/F
v)
은 배양 개시 전(before)
과배양5
일후(after)
에15
분간암적응시킨후에Phyto-PAM (Walz, Effecltrich, Germany)
을사용하여측정하였다. Phyto- PAM
의세팅조도는8, 16, 32, 64, 90, 120, 210, 295, 350, 435 μmol photons m
-2s
-1로설정하여20
초간격으로측정하였다.
광 합성효율측정은아래의식으로계산하였다.
F
v/F
m= (F
m-F
o)/F
mF
m=
최대형광효율, F
o=
최소형광효율형광변수
(F
v)
는암적응후최대형광과최소형광의차이를나 타내며(Jesus et al., 2006),
광양자수율(quantum yield, F
m/F
v)
은광합성명반응의반응중심중하나인광계2 (photosystemII,
PSII)
의광화학반응에대한양자수율또는양자효율을의미한다
.
측정한빠른광반응곡선은광양자수율을바탕으로전자전 달률(electron transport rate, ETR)
을계산하고,
이를이용해광 합성-
조도곡선을추정하여상대적인광합성변수(α)
를구할수 있다(Schreiber et al., 1994).
E
k= ETR
max/α
E
k=
광포화계수, ETR
max=
최대전자전달률결 과
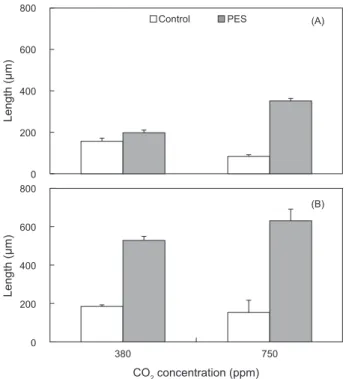
배아생장
구멍갈파래유주자가접종되어착생시킨후에배아의길이는
24.63±0.57 μm (mean±SE, n=90)
였으며,
해수의CO
2농도 가750 ppm
인PES
배양액,
그리고100 μmol photons m
-2s
-1에 서635.53±61.45 μm
로최대생장을보였으며, 750 ppm
의멸 균해수와50 μmol photons m
-2s
-1에서가장느린생장을보였 으며,
이때길이는79.33±0.07 μm
였다(Fig. 2).
낮은실험조 도인50 μmol photons m
-2s
-1에서, CO
2농도가380
과750 ppm
범위에서는영양염농도가증가할수록생장이촉진되었다.
또 한,
배아는고조도인100 μmol photons m
-2s
-1에서도750 ppm
까지적당한영양염증가
(PES)
조건에서빠른생장을보여조도
,
영양염과CO
2농도가증가할때배아는최대생장을보였다(Fig. 2).
유엽 생장률
구멍갈파래유엽의길이는배양개시전에
4.05±0.09 mm (mean±SE, n=180)
였고,
상대생장률은750 ppm
의PES
배양 액,
그리고50 μmol photons m
-2s
-1에서4.41 % day
-1로최대였 고, 380 ppm
의멸균해수와50 μmol photons m
-2s
-1에서최소(0.33 % day
-1)
로나타났다(Table 1).
유엽의생장은380 ppm
에비해750 ppm
에서50
과100 μmol photons m
-2s
-1에서모 두빠른생장을보였으며,
최대생장은영양염이풍부한PES
와750 ppm
에서4.41% day
-1로나타났으며,
멸균해수의경우 에도CO
2농도가증가한750 ppm
에서대조구에비해높은생 장을보였다(Table 1).
Fig. 1. Growth stage of Ulva australis: A, zygotes; B, germlings; C, juvenile; D, E, adult.
해수의 CO2, 영양염 농도 및 조도와 구멍갈파래 생장
99
성체 광합성
구멍갈파래 성체의 광합성효율
(F
m/F
v)
은 배양 개시 전에0.73±0.05
였으며,
배양5
일후에750 ppm
의멸균해수와50
μ mol photons m
-2s
-1에서0.83±0.01
로최대였고750 ppm
의 멸균해수와100 μmol photons m
-2s
-1에서0.63±0.03
으로최소 였으며초기값에비해낮았다(Fig. 3).
성체의광합성효율은고 조도(100 μmol photons m
-2s
-1)
의380 ppm
에서평균0.68,
그 리고750 ppm
에서0.65
로서배양개시전(0.73)
에비해감소하 였다.
저조도인50 μmol photons m
-2s
-1에서는배양개시전에 비해모든실험구에서높게나타났고광합성효율은750 ppm
에서380 ppm
에비해높았다(Fig. 3)
저조도에서광합성효율은 영양염농도와관계없이해수의CO
2 농도가380
에서750 ppm
으로증가할때정비례로높아졌다(Fig. 3).
최대전자전달률
(ETR
max)
은실험개시전에122.45±3.71
였 으며,
배양5
일후모든CO
2농도에서저조도인50
에서고조도 인100 μmol photons m
-2s
-1에비해모두높았으며, 380
과750
Fig. 2. Effects of CO2 concentration and nutrient levels on thegrowth of Ulva australis germlings. Germlings were cultured for 10 days under 50 (A) and 100 μmol photons m-2s-1 (B). Vertical bars indicate standard errors (n = 3 replicates).
CO2 concentration (ppm)
(A)
(B)
Length (μm)Length (μm)
380 800
600 400 200 0 800 600 400 200 0
750
Control PES
CO2 concentration (ppm)
(A)
(B)
Yield (Fv/ Fm)Yield (Fv/ Fm)
380 1.0
0.8 0.6 0.4 0.2 0.0 1.0 0.8 0.6 0.4 0.2 0.0
750
Control PES
Table 1. Average relative growth rate (% day-1) of Ulva australis juveniles grown under two CO2, nutrient and irradiance levels. Ju- veniles grew for 5 days at 20 ℃ and a photoperiod of 12:12 h LD.
Values are means ± SE (n=5 replicates) Irradiance
Nutrient CO2 concentration (ppm)
(μmol photons m-2s-1) 380 750
50
Control 0.33±0.22 3.28±0.46 PES 0.91±0.76 4.41±1.57 Mean 0.62±0.29 3.85±0.56
100
Control 0.63±0.58 1.38±0.41 PES 2.09±0.25 3.42±0.35 Mean 1.36±0.73 2.40±1.02 PES, Provasoli's medium.
Table 2. Effects of CO2 concentration and nutrient levels on the photosynthesis parameters of Ulva australis adult discs, which were cultured for 5 days under 50 and 100 μmol photons m-2s-1, 20 ℃, and 12:12 h LD. Values are mean ± SE (n=5 replicates)
Parameter Irradiance CO2 concentration Nutrient concentration
(μmo lphotons m-2s-1) (ppm) Control PES
ETRmax
50 380 119.91±11.99 178.56±37.90
750 118.72±18.75 160.88±13.83
100 380 86.61±8.72 125.31±34.19
750 56.24±6.69 59.26±10.65
Ek
50 380 159.39±11.31 242.10±72.65
750 182.70±24.83 240.35±24.25
100 380 170.75±5.32 204.47±41.51
750 109.54±4.49 131.72±4.38
α
50 380 0.75±0.03 0.77±0.07
750 0.64±0.03 0.68±0.02
100 380 0.50±0.03 0.60±0.06
750 0.52±0.05 0.44±0.08
PES, Provasoli's medium; ETR, electron transport rate.
전다빈
ㆍ
나연주ㆍ
유옥환ㆍ
최한길100
ppm
에서는PES
배양액에서최대였다(Table 2).
최소ETR
max 값은멸균해수에CO
2농도가750 ppm
그리고고조도인100 μmol photons m
-2s
-1에서나타났다.
광포화계수
(E
k)
는실험개시전에185.95±5.63 (n= 5 rep- licates)
이었으며,
배양5
일후750 ppm
의PES
배양액과50 μmol photons m
-2s
-1에서240.35±24.25(380 ppm
의PES
배양 액과50 μmol photons m
-2s
-1에서242.10±72.65)
로서최대였 다(Table 2). E
k값은일반적으로50 μmol photons m
-2s
-1에서높 았으며,
영양염농도가낮은멸균해수와CO
2농도가높은750 ppm
에서낮은값을보였다(Table 2).
구멍갈파래성체의배양전기울기
(α)
값은0.66±0.00 (n=5 replicate)
였으며, 5
일 후에는380 ppm
의PES
배양액과50 μmol photons m
-2s
-1에서0.77±0.07
로서최대였고750 ppm
의PES
와100 μmol photons m
-2s
-1에서최소였다(Table 2).
기울 기값은저조도인50 μmol photons m
-2s
-1에서CO
2농도는380
과750 ppm
에서높은값을보였다.
고 찰
구멍갈파래배아의길이는배양
10
일후에50
과100 μmol photons m
-2s
-1에서각기276.21±17.11 μm
와329.83±29.00 μm
로고조도에서빠른생장을보였고,
해수의CO
2농도가750
ppm,
그리고고영양염상태에서도생장은촉진되었다.
켈프종인
Macrocystis pyrifera
의배우체는해수의CO
2농도가산업혁 명이전인280 ppm
에비해820 ppm
에서약32%
빠르게생장 한것으로확인되었다(Roleda et al., 2012).
하지만,
경제종인Saccharina japonica
의암배우체는배양10
일후에현재해수의CO
2 농도인380 ppm
에서750
과1,000 ppm
에비해2-3
배빠 른생장을보이는것으로확인되었다(Oh, 2014).
따라서,
지구 온난화에의한해수의CO
2 농도증가는현미경적단계인배아 의생장에영향을주는것으로확인되었으며,
종에따른차이를 보이는것으로확인되었다.
해수의CO
2 농도의증가는아직먼 미래라고생각할지모르지만,
켈프숲에서CO
2 농도는하루에80-530 ppm
의변화를보이고있고(Delille et al., 2000),
구멍갈 파래성체에서방출된유주자는조간대암반에착생및생장하 므로온난화에의한CO
2 농도와인간활동에의한영양염증가 는구멍갈파래배아의생장을촉진하는것으로나타남으로써 녹조대발생의빈도는증가될것으로사료된다.
본연구에서구멍갈파래유엽의생장과성체의광합성은
50
과100 μmol photons m
-2s
-1에서증가된CO
2농도와영양염(PES)
에서촉진되었다.
녹조류인가시파래(Ulva prolifera)
는현재의 해수CO
2농도인390 ppm
에비해1,000 ppm
에서빠른생장을 보여본연구결과와유사하게CO
2농도와생장은정비례관계 를보였다(Xu and Gao, 2012).
해수중의CO
2농도가증가하면 생장이촉진되는종은홍조류의Pyropia yezoensis, Gracilaria spp.
와Lomentaria articulata (Gao et al., 1991, 1993; Kubler et al., 1999)
와갈조류의Sargassum fusiforme (Zou, 2005)
가 있으나,
반면에Gracilaria tenuistipitata, Pyropia leucostica
와Porphyra linearis
종은CO
2 농도가증가하면생장이감소되었 다(Garcia-Sanchez et al., 1994; Israel et al., 1999; Mercado et al., 1999).
갈파래류인Ulva lactuca
는CO
2 농도가432
와635 ppm
에서생장이촉진되었으나884 ppm
에서감소를보였고, Ulva intestinalis
는 용존무기탄소(DIC, dissolved inorganic carbon)
가증가할때생장이감소되었고(Andria et al., 2001), Ulva clathrata
는884 ppm
에서감소되었으나(Kerrison et al., 2012), Ulva prolifera
는1,000 ppm
에서증가하는패턴을보임 으로써(Xu and Gao, 2012)
동일한속(genus)
에서도CO
2 농도 증가에따른생리적반응은다르게나타났다.
해수의CO
2 농도 가높고pH
가낮은자연생태계(
예,
화산활동이활발한해역)
에 서는홍조류의산호말류,
녹조류의Halimeda spp.
와갈조류의Padina spp.
와같이CaCO
3이필요한해조류는풍도가감소되 고높은CO
2 농도에서잘자라는해초(seagrass)
와해조류(
옥덩 굴,
대마디말,
모자반류)
가번무한다고하였다(Hall-Spencer et al., 2008).
따라서,
지구온난화에의한수온상승과CO
2 농도증 가에따른해양산성화,
그리고인간활동에의한영양염의증가 가연안생태계의일차생산자인해조류의군집구조의변화와종 다양성에영향을미칠것으로사료된다.
구멍갈파래 성체의 광합성은 조도와영양염과
CO
2 농도에 Fig. 3. Average yield (Fv/Fm) of Ulva australis frond discs culturedfor 5 days under three CO2 concentration and nutrient levels, and at 50 (A) and 100 μmol photons m-2s-1 (B). Vertical bars indicate standard errors (n = 3 replicates).
CO2 concentration (ppm)
(B)
Length (μm)Length (μm)
380 800
600 400 200 0 400 200 0
750
CO2 concentration (ppm)
(A)
(B)
Yield (Fv/ Fm)Yield (Fv/ Fm)
380 1.0
0.8 0.6 0.4 0.2 0.0 1.0 0.8 0.6 0.4 0.2 0.0
750
Control PES
해수의 CO2, 영양염 농도 및 조도와 구멍갈파래 생장
101
따라다르게나타났으며
, 50
에서100 μmol photons m
-2s
-1에 비해높았고, PES
배양액과750 ppm
에서 최대였다.
녹조류Cladophora sp.
의광합성(F
v/F
m)
은360
과1,000 ppm
에서각기22.25
와40.08%
증가하였고(Ping et al., 2014), Ulva rigida
는 해수의CO
2농도가증가(93%)
할때광합성효율이감소하여 본연구의구멍갈파래(Ulva australis)
와생리적반응이유사하 였다(Gordillo et al., 2003).
한편,
해조류중에서CaCO
3를세포 벽에가지는유절·
무절산호조류인홍조류Amphiroa sp.
의광 합성은1,000 ppm
에서는40%
감소였고, Lithophyllum sp.
는550 ppm
에서도감소하였으며,
최근에Gao and Zheng (2010)
은높은CO
2농도에서Corallina sessilis
의생장이억제된다고 하였다.
따라서,
해수의CO
2농도의증가는탄산칼슘을가지는 종의광합성에나쁜영향을미치며,
갈파래류와대마디말류는 종에따라차이를보이지만750
에서1,000 ppm
까지광합성이 증가하고나아가본연구결과에서보여준것처럼CO
2농도와 영양염농도가동시에증가한연안에서향후대발생에적신호 가나타나고있어대책마련이시급한실정이다.
해조류의 생활사에서현미경적인단계
(
예,
유주자,
배아,
접 합자,
포자등)
는무생물적환경(
수온,
건조,
영양염등)
과생물 학적환경(
경쟁,
초식등)
에매우민감하게반응하므로개체군 증감은생활사의초기에결정되며(Coelho et al., 2000),
환경 에대한생리적반응도생활사단계에따라다른것으로알려 져있다(Kim et al., 2011).
예를들면,
녹조류Ulva linza
의광 합성은적절한온도와조도범위에서는생활사초기단계(early stage)
에있는개체가후기단계(late stage)
에있는엽체에비 해높았으며,
열악한조건에서는반대로후기단계에서광합성 이높게나타났다(Kim et al., 2011).
해수의CO
2농도증가는 갈조류Laminaria hyperborea
의 생활사에서 암배우체의 난(oogonium)
과아포체(juvenile sporophyte)
형성및생장을촉 진시켰으나,
포자체의광합성에서는큰차이를보이지않았다(Olischläger et al., 2012).
최근, Saccharina japonica
의암배우 체는CO
2농도가380 ppm
인실험구에서750
과1,000 ppm
실 험구에비해2-3
배의빠른길이생장을보였으나,
성체의광합성은
750 ppm
에서높게나타났으며수온과상호작용이있음을확인하였다
(Oh, 2014).
본연구에서구멍갈파래의배아와유엽 의생장은380 ppm
에비해750 ppm
의CO
2농도에서확연하 게증가하였으나,
성체의광합성은380
과750 ppm
에서유의차 가없었다.
하지만, Ulva prolifera
는배아생장과성체광합성 이1,000 ppm
에서390 ppm
에비교하여확연하게좋았음이확 인되었다(Xu and Gao, 2012).
따라서,
본연구에서는생활사의 생장단계에따라CO
2농도,
영양염농도,
그리고이들의상호 작용이다름이확인되었으며,
이결과는배아가환경변화(
특히 해양산성화,
수온상승,
영양염등)
에민감하므로배아의생리적 반응을밝히는것이향후기후변화에대응하는중요한정보임 을시사한다고볼수있다.
사 사
이논문은
2011
년도정부(
교육과학기술부)
의재원으로한국 연구재단의기초연구사업지원을받아수행된연구(
과제번호NRF-2011-0012519)
로서이에감사드립니다.
또한,
이논문 은해양수산부의재원으로해양생명공학기술개발사업연구개 발비지원에의해수행되었습니다.
References
Andria JR, Brun FG, Perez-Llorens JL and Vergara JJ. 2001.
Acclimation responses of Gracilaria sp. (Rhodophyta) and Enteromorpha intestinalis (Chlorophyta) to changes in the external inorganic carbon concentration. Bot Mar 44, 361- 370. http://dx.doi.org/10.1515/BOT.2001.046.
Carlton JT. 2000. Global change and biological invasions in the oceans. In: Invasive Species in a Changing World. Mooney HA and Hobbs RJ, eds. Island Press, Washington DC, USA, 31-53.
Choi TS, Kang EJ, Kim J and Kim KY. 2010. Effect of salin- ity on growth and nutrient uptake of Ulva pertusa (Chlo- rophyta) from an eelgrass bed. Alage 25,17-26. http://dx.doi.
org/10.4490/algae.2010.25.1.017.
Coelho SM, Rijstenbil, JW and Brown MT. 2000. Impacts of anthropogenic stresses on the early development stages of seaweeds. J Aquar Ecosyst Stress Recovery 7, 317-333.
http://dx.doi.org/10.1023/A:1009916129009.
Connell SD and Russell BD. 2010. The direct effects of in- creasing CO2 and temperature on non-calcifying organ- isms: increasing the potential for phase shifts in kelp forests.
Proc R Soc B 277, 149-1415. http://dx.doi.org/10.1098/
rspb.2009.2069.
Delille B, Delille D, Fiala M, Prevost C and Frankignoulle M.
2000. Seasonal changes of pCO2 over a subantarctic Mac- rocystis kelp bed. Polar Biol 23, 706-716. http:// dx.doi.org/
10.1007/s003000000142.
Deng Y, Tang X, Huang B and Ding L. 2011. Life history of Chaetomorpha valida (Cladophoraceae, Chlorophyta) in culture. Bot Mar 54, 551-556. http://10.1515/ BOT.2011.066.
Deng Y, Tang X, Huang B and Ding L. 2012. Effect of tempera- ture and irradiance on the growth and reproduction of the green macroalga, Chaetomorpha valida (Cladophoraceae, Chlorophyta). J Appl Phycol 24, 927-933. http://dx.doi.
org/10.1007/s10811-011-9713-0.
Feely RA, Sabine CL, Lee K, Berelson W, Kelypas JA, Fabry VJ and Millero FJ. 2004. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 305, 362-366.
http://dx.doi.org/10.1126/science.1097329.
Floreto EAT, Hirata H, Yamasaki S and Castro SC. 1994. Ef- fect of salinity on the growth and fatty acid composition of Ulva pertusa Kjellman (Chlorophyta). Bot Mar 37,151-155.