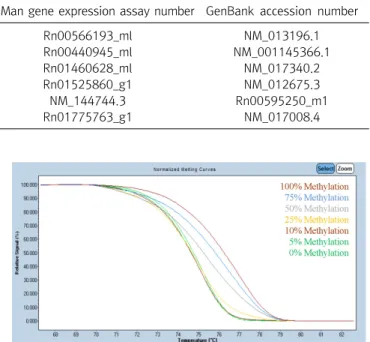
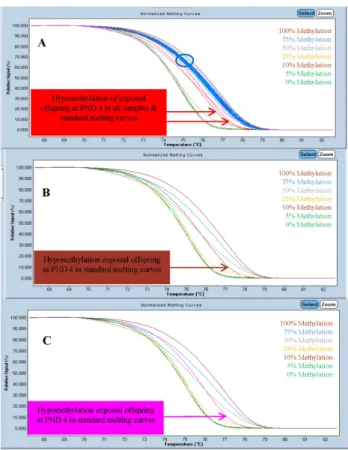
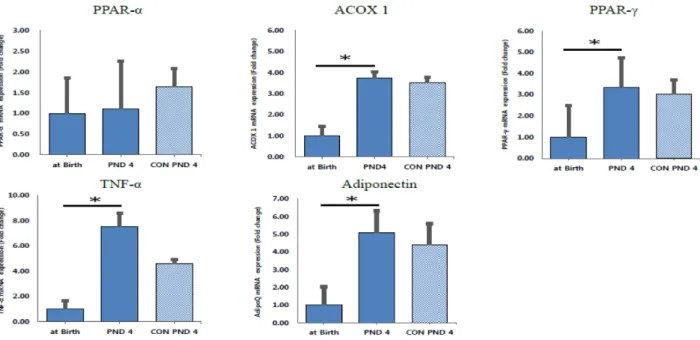
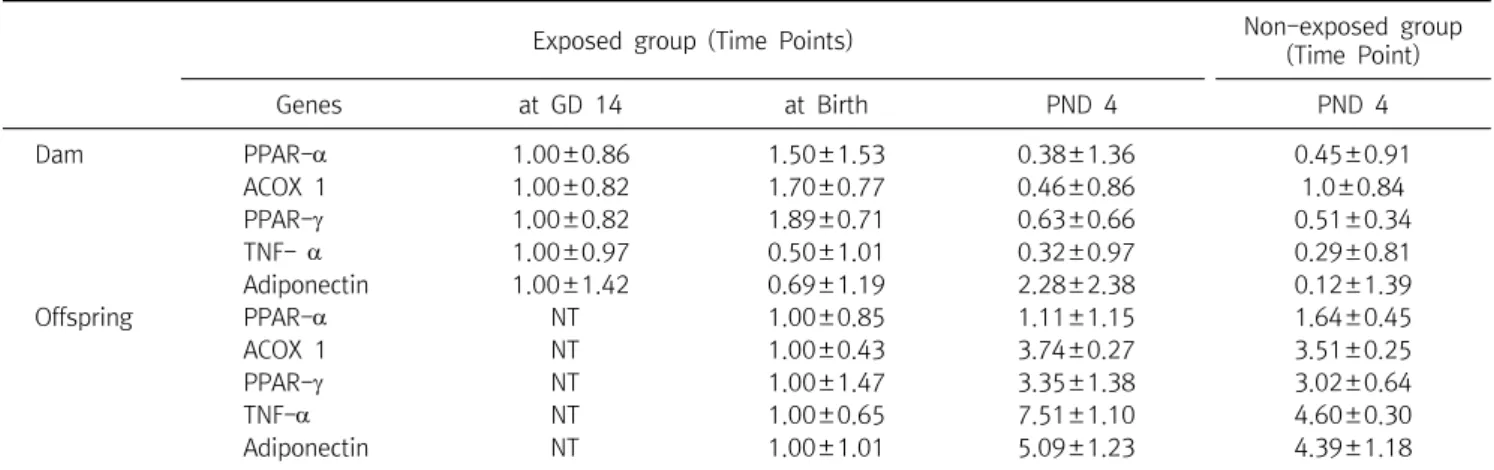
Global DNA Methylation Patterns and Gene Expression Associated with Obesity-Susceptibility in Offspring of Pregnant Sprague-Dawley Rats Exposed to BDE-47 and BDE-209
12
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
수치

+2
관련 문서
GDP impact of COVID-19 spread, public health response, and economic policies. Virus spread and public
3) ECDC, 2020. Interim guidance for environmental cleansing in non-healthcare facilities exposed to 2019-nCoV 4) NEA, 2020. Interim list of household products and active
Micro- and nano-sized pores were formed on the surface of the alloy using PEO and anodization methods, and the pore shape change according to the Zr
There was a significant increase in new bone formation in the group in which toothash and plaster of Paris and either PRP or fibrin sealants were used, compared with the groups
Mean and standard deviation of Labial margin, Lingual margin, and Incisal margin gaps at the three groups (unit : ㎛ )..
Methods: Eighty ASA physical status I and II pediatric patients (1~7 years) were randomly divided into four groups: Group I (Lactated Ringer`s solution, n=20), Group II
Mortality and clinical signs in SD rats about single oral administration after 3 weeks with callus extract ..... Hematological values in SD rats administered orally
E and prickly pear cactus in Sprague Dawley rats fed fish oil by examining erythrocyte Na-leak, platelet aggregation, TBARS production and plasma
