10 This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
J. Mushrooms 2020 March, 18(1):10-19 http://dx.doi.org/10.14480/JM.2020.18.1. 10 Print ISSN 1738-0294, Online ISSN 2288-8853
© The Korean Society of Mushroom Science
Si Young Ha(Complete a doctorate), Ji Young Jung(Research professor), Jai Hyun Park(Graduate student), Dong Hwan Lee(Graduate student), Ji Won Choi(Graduate student), Jae-Kyung Yang(Professor)
*Corresponding author E-mail : [email protected]
Tel : +82-55-772-1862, Fax : +82-55-772-1869 Received January 17, 2020
Revised March 17, 2020 Accepted March 24, 2020
Effect of light-emitting diodes on cordycepin production in submerged Cordyceps militaris cultures
Si-Young Ha, Ji-Young Jung, Jai-Hyun Park, Dong-Hwan Lee, Ji-Won Choi, and Jae-Kyung Yang*
Division of Environmental Forest Science/Institute of Agriculture and Life Science, Gyeongsang National University, Jinju, 52828, Republic of Korea
ABSTRACT:
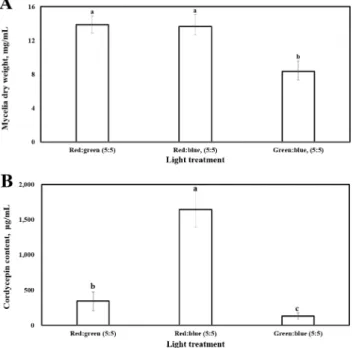
Light is an important factor for cordycepin production in Cordyceps militaris. We investigated the effects of different light-emitting diode (LED) conditions including various LED wavelengths and their combinations on cordycepin production in Cordyceps militaris cultivated in submerged culture. The results of our study showed that the combinations of LED wavelengths were more beneficial than single LED sources for cordycepin production. Among the three tested wavelength combinations, the greatest effects for cordycepin production were observed for the red:blue light combination at the wavelength ratio of 5:5 or 3:7.
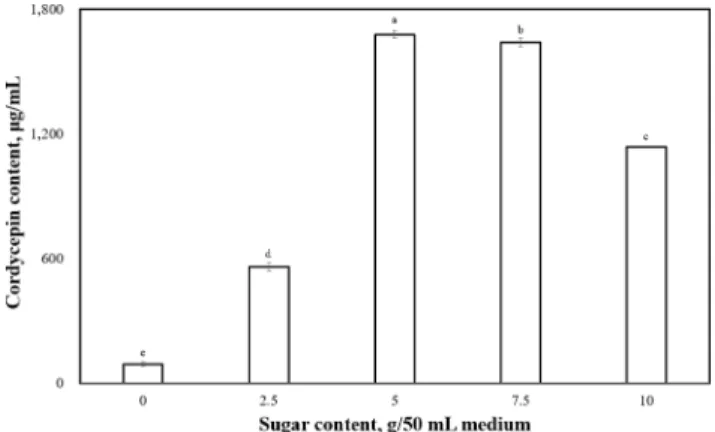
The optimal culture conditions were 19.2278 h/day of illumination time; 9.19497 g/50 mL of glucose content in the media; and 53.112 h of cultivation time. Our model predicted a maximum yield of 2860.01 μg/mL cordycepin. Finally, to verify the calculated maximum, we performed experiments in the culture media representing the obtained optimum combination and the cordycepin yield of 2412.5 μg/mL.
KEYWORDS: