세포벽 분해효소 처리가 현미의 페놀화합물 함량과 항산화 활성에 미치는 영향
조동화1․이석기1․박지영1․박혜영1․최혜선1․최인덕1․한상익1․정현정2․정두연2․오세관1
1농촌진흥청 국립식량과학원 중부작물부
2전남대학교 식품영양과학부
Effect of Cell Wall Degrading Enzyme Treatment on the Phenolic Content and Antioxidant Activity of Brown Rice
Dong-Hwa Cho
1, Seuk-Ki Lee
1, Jiyoung Park
1, Hye-Young Park
1, Hye-Sun Choi
1, Induck Choi
1, Sang-Ik Han
1, Hyun-Jung Chung
2, Duyun Jeong
2, and Sea-Kwan Oh
11
Department of Central Area Crop Science, National Institute of Crop Science, Rural Development Administration
2
Department of Food and Nutrition, Chonnam National University
ABSTRACT Enzymatic treatments of brown rice flour (BRF) using commercial cell wall degrading enzymes (Celluclast 1.5 L, Pectinex Ultra SP-L) to enhance the phenolic acid content and antioxidant property were investigated.
The phenolic acid composition and content were changed substantially by a treatment with commercial cell wall degrad- ing enzymes. These enzymes increased the sum of free phenolic acids by 8.4∼11.2 times, especially for ferulic and p-coumaric acid. Celluclast 1.5 L appeared to be more effective in increasing the total phenolic content than Pectinex Ultra SP-L. The GABA content of BRF with the Celluclast 1.5 L treatment was approximately 43 times higher than that of the control (32.7 vs. 0.76 mg/100 g sample). The ABTS radical scavenging activities of the enzyme treated samples were significantly higher than those of the control. In particular, the Celluclast 1.5 L treated sample was approximately 2.5 times higher than that of the control sample. Enzymatic hydrolysis of the cell wall polysaccharides in BRF could be used as an effective procedure for increasing not only the phenolic acid and GABA content, but also the antioxidant activity.
Key words: cell wall degrading enzyme, brown rice flour, phenolic acids, antioxidant properties, γ-aminobutyric acid
Received 21 November 2017; Accepted 17 May 2018
Corresponding author: Sea-Kwan Oh, Crop Post-harvest Technol- ogy Research Division, Department of Central Area Crop Science, National Institute of Crop Science, Rural Development Administra- tion, Suwon, Gyeonggi 16429, Korea
E-mail: [email protected], Phone: +82-31-695-0610
서 론
쌀은 한국을 비롯한 다양한 나라에서 주식으로 이용되는 식량 작물로 연간 약 7억 5천만 톤이 생산되며 밀, 옥수수와 함께 세계에서 가장 많이 생산되는 곡류 중 하나이다(1). 전 세계 쌀 생산 및 소비량은 꾸준히 증가하는 추세로 지난 50 년간 세계 쌀 소비량은 2배 이상 증가하였다(1). 이와 대조 적으로 한국 및 일본의 쌀 소비량은 점진적으로 감소하는 추세이다. 1979년 우리나라 국민 1인당 연간 양곡 소비량은 135.6 kg이었으나 점차 감소하여 2016년에는 61.9 kg에 불과하였다(2,3).
쌀은 주로 백미 형태로 주로 이용되었으나, 2000년대 이 후 생활 습관병 환자 증가, 웰빙(well-being) 트렌드 유행과
함께 현미에 대한 관심 및 이용이 증가하였다. 현미는 도정 을 거치지 않은 쌀로 영양 및 기능성이 풍부한 미강과 쌀눈 을 가지고 있기 때문에 백미보다 페놀산, 비타민, 감마아미 노낙산(γ-aminobutyric acid, GABA), 감마오리자놀(γ- oryzanol) 등을 비롯한 기능성 성분 함량이 높다(4).
페놀산(phenolic acid)은 다양한 식물체에서 발견되는 식 물화학물질(phytochemical)로 항산화, 항염증, 항당뇨, 항 비만, 항암, 심혈관계 질환 예방 등 다양한 생리활성을 가지 고 있다(5). 페놀산은 벤조산(benzoic acid)과 신남산(cin- namic acid)에 하나 이상의 치환기를 가지고 있는 형태의 화 합물을 총칭하는 것으로, 그 형태에 따라 유리 페놀산(free phenolic acid)과 불용성 페놀산(bound phenolic acid)으 로 나뉜다(5). 대부분 페놀산은 불용성 형태로 존재하는데, 이들은 주로 식물 세포벽의 탄수화물과 에스터 결합(ester bond)을 하고 있다(6,7). 현미에서 페놀산의 80~90%는 불 용성 형태로 존재하며, 이들 대부분은 현미의 배유가 아닌 미강층에 존재한다(7). 쌀에는 ferulic acid,
p
-coumaric acid, vanillic acid, gallic acid, caffeic acid 등 다양한 페놀 산이 있지만, 특히 ferulic acid와p
-coumaric acid의 함량이 높다(7,8). Ferulic acid와
p
-coumaric acid는 벼과 식물 에서 주로 많이 존재하는 페놀산으로 주로 식물의 과피(al- eurone layer)와 호분층(pericarp)의 세포벽, 특히 아라비 노자일란(arabinoxylan)과 에스터 결합을 이루고 있다. 하 지만 페놀산의 생리활성은 불용성 형태보다 유리형태에서 강하게 나타난다. 따라서 쌀에 풍부하게 존재하는 불용성 페놀산을 유리형태로 바꿀 수 있다면 쌀 및 쌀 가공식품의 건강 기능성을 높일 수 있을 것으로 생각한다.식물의 세포벽은 셀룰로스(cellulose), 헤미셀룰로스(he- micellulose), 펙틴(pectin), 리그닌(lignin) 등으로 구성되 어 있으며, 이들은 다시 구성하고 있는 당류에 따라 xylan, arabinoxylan, mannan, galactomannan 등으로 나뉠 수 있 다(9). 이들 대부분은 사람이 소화할 수 없는 식이섬유의 일 종이기 때문에 이들과 결합하고 있는 불용성 페놀산 또한 체내에서 흡수 및 활용이 힘들다. 세포벽 분해효소는 다양한 세포벽 구성 다당류를 분해할 수 있는 효소로 cellulase, pectinase, β-glucanase, hemicellulase, xylanase, ara- banase 등이 있다. 상업적으로 이용되는 세포벽 분해효소는 보통 미생물에서 얻어지며, 이들은 곡류뿐만 아니라 다양한 식물체의 단백질 및 지방의 추출 효율을 높이는 데 주로 사 용된다(10,11). 또한, 세포벽 분해효소 처리는 세포벽과 결 합하고 있는 불용성 페놀산을 유리 페놀산으로 전환시켜줄 수 있을 것으로 예상한다. 하지만 세포벽 분해효소 처리에 따른 식물체, 특히 식량작물의 건강 기능성 및 생리활성 변 화에 대한 연구는 일부만 진행되어 있다. 따라서 본 연구는 세포벽 분해효소를 현미 가루에 처리함으로써 현미 가루의 기능성을 높이고자 하였으며, 이를 위하여 페놀산 함량, GABA 함량 및 항산화 활성을 조사하였다.
재료 및 방법
실험재료
본 실험에는 2016년 농촌진흥청 국립식량과학원에서 재 배 및 수확된 삼광벼(
Oryza sativa
cv.Samkwang
)가 사용 되었으며, 2016년 3월에 제현한 현미를 4°C에서 냉장 보관 하며 실험에 사용하였다. 효소 처리를 위한 현미 가루는 분 쇄 후 100 메시 체(pore size 150 μm)에 통과시켜서 일정 크기 이하의 분말로 얻었다. 실험에 사용된 세포벽 분해효소 는 Pectinex Ultra SP-L(Pectinase fromAspergillus aculeatus
, declared activity 3,800 units/mL)과 Cellu- clast 1.5 L(cellulase fromTrichoderma reesei
, declared activity 750 EGU g-1)로 Novozyme A/S(Bagsvaerd, Denmark)에서 구입하여 사용하였다.세포벽 분해효소 처리
밀폐된 용기에 현미 가루 15 g(dry basis)과 1%(v/v) 세 포벽 분해효소(Pectinex Ultra SP-L, 혹은 Celluclast 1.5 L) 수용액(10 mL)을 넣고 완전히 혼합하여 주었다. 혼합
후 즉시 이를 인큐베이터(40°C)에서 24시간 동안 반응시키 고 반응이 완료된 현미 가루는 꺼내어 페트리 접시에 옮겨 담아 열풍건조(50°C, 8시간)하였다. 완전히 건조된 시료는 막자사발로 분쇄하고 100 메시 체(pore size 150 μm)에 통과시켜 일정 크기 이하의 현미 가루로 얻었다. 대조군은 3차 증류수 10 mL를 동일한 조건으로 처리하였다.
현미 가루의 페놀산 분획 추출
시료(2 g, dry solids)에 100 mL의 70% 에탄올을 가한 다음 3시간 동안 상온에서 추출하고 원심분리 하였다. 침전 물에 다시 100 mL의 70% 에탄올을 가하여 동일한 방법으 로 2회 더 추출하였다. 얻어진 상등액은 진공회전농축기를 이용하여 감압 농축한 후 10% 에탄올(20 mL)에 재분산하 고 1 M HCl을 이용하여 pH를 2.0으로 조정하였다. 산성화 된 추출물에 에틸아세테이트 : 에틸에테르(1:1, v/v) 혼합액 을 가하고 30분간 교반 후 원심분리 하여 에틸아세테이트 : 에틸에테르 혼합액 분획층만 수거하였으며, 남은 잔사에 다시 에틸아세테이트 : 에틸에테르(1:1, v/v) 혼합액을 가하 여 동일한 방법으로 총 5회 추출하였다. 얻어진 추출물은 진공회전농축기를 이용하여 감압 농축한 후 메탄올 3 mL에 재분산하고 이후 실험에 사용하였다.
현미 가루의 총 페놀 함량 및 유리 페놀산 분석
총 페놀 화합물 함량은 Folin-Ciocalteu 방법(12)을 일부 수정하여 분석하였다. 페놀산 분획 추출물 0.1 mL에 2%
Na2CO3 2 mL를 첨가하여 혼합한 후 실온에서 3분간 반응 시킨 다음 50% Folin 시약을 첨가하였다. 암소에서 30분 동안 방치한 후 750 nm에서 흡광도를 측정하였다. Gallic acid로 표준 검량 곡선을 작성한 후 페놀산 분획 추출물의 총 페놀 화합물 함량은 gallic acid equivalent(mg GAE/
100 g sample)로 나타냈다.
시료의 페놀산 구성 및 함량은 reversed-phase high- performance liquid chromatography(RP-HPLC)를 이용 하여 측정하였다. 페놀산 분획 추출물을 0.25 μm syringe filter로 여과한 후 HPLC UltiMate 3000 system(Thermo Fisher Scientific, Madison, WI, USA)으로 분석하였으며, 분석 칼럼은 Capcell-pak C18 column(4.6 mm i.d.×250 mm length, 5 µm, Shiseido, Tokyo, Japan)을 사용하였다.
칼럼 온도와 시료 주입량은 각각 40°C, 1 μL로 설정하였다.
이동상은 A(0.02% trifluoroacetic acid in deionized wa- ter)와 B(0.02% trifluoroacetic acid in methanol)를 0.4 mL/min 유속으로 흘려주었으며, 기울기 용리조건(gradient elution)은 다음과 같다: 0~30 min, 0~15% B; 30~45 min, 15~25% B; 45~60 min, 25~50% B; 60~65 min, 50~100
% B; 65~67.5 min, 100% B; 67.5~70 min, 100~0% B;
70~75 min, 0% B. 페놀산 종류에 따라 검출 UV 파장을 달리하였는데, 하이드록시벤조산(hydroxybenzoic acid) 계 열은 280 nm, 하이드록시남산(hydroxycinnamic acid) 계
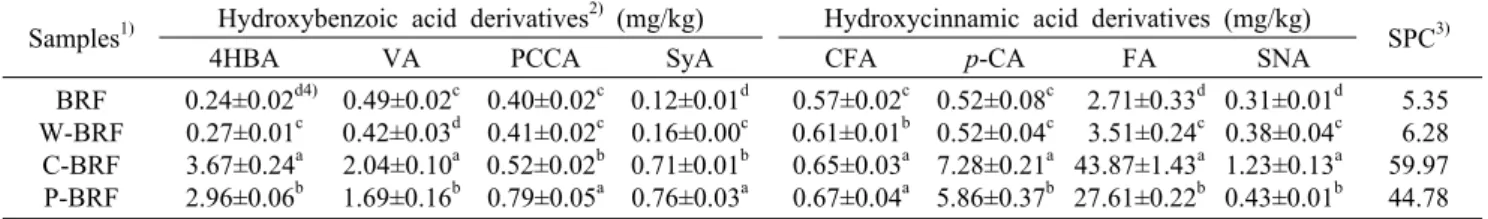
Table 1. Phenolic acid composition of enzyme treated brown rice flours
Samples1) Hydroxybenzoic acid derivatives2) (mg/kg) Hydroxycinnamic acid derivatives (mg/kg)
SPC3)
4HBA VA PCCA SyA CFA
p-CA
FA SNABRF W-BRF
C-BRF P-BRF
0.24±0.02d4) 0.27±0.01c 3.67±0.24a 2.96±0.06b
0.49±0.02c 0.42±0.03d 2.04±0.10a 1.69±0.16b
0.40±0.02c 0.41±0.02c 0.52±0.02b 0.79±0.05a
0.12±0.01d 0.16±0.00c 0.71±0.01b 0.76±0.03a
0.57±0.02c 0.61±0.01b 0.65±0.03a 0.67±0.04a
0.52±0.08c 0.52±0.04c 7.28±0.21a 5.86±0.37b
2.71±0.33d 3.51±0.24c 43.87±1.43a 27.61±0.22b
0.31±0.01d 0.38±0.04c 1.23±0.13a 0.43±0.01b
5.35 6.28 59.97 44.78
1)BRF: brown rice flour, W-BRF: water treated BRF, C-BRF: Celluclast 1.5 L treated BRF, P-BRF: Pectinex Ultra SP-L treated
2)4HBA: 4-hydroxybenzoic acid, VA: vanillic acid, PCCA: protocatechuic acid, SyA: syringic acid, CFA: caffeic acid, p-CA: p-couma-BRF.
ric acid, FA: ferulic acid, SNA: sinapic acid.
3)SPC: sum of individual phenolic acids content.
4)Values (dry weight basis) with different letters (a-d) within a column are different with statistical significance (P<0.05).
열은 325 nm 파장에서 측정하였다.
현미 가루의 감마아미노낙산 함량 분석
감마아미노낙산과 글루탐산(glutamic acid) 분석을 위하 여 시료 1 g과 3% TCA 5 mL를 혼합한 다음 3시간 동안 추출하였다. 이후 원심분리(3,000×
g
, 15 min, 4°C)를 통하 여 얻어진 상층액을 0.25 μm syringe filter로 여과하였다.여과된 시료(20 μL)는 아미노산 분석기(L-8800, Hitachi Ltd., Tokyo, Japan)로 분석하였다.
현미 가루의 항산화 활성 측정
페놀산 분획 추출물의 항산화력은 1,1-diphenyl-2-pic- rylhydrazyl(DPPH) 라디칼 소거능, 2,2’-azino-bis(3- ethylbenzothiazoline-6-sulfonic acid)(ABTS) 라디칼 소 거능 방법을 이용하여 측정하였다. DPPH 라디칼 소거능은 Bondet 등(13)의 방법을 일부 수정하여 수행하였다. 0.2 mM DPPH 용액 0.8 mL에 페놀산 분획 추출물 0.2 mL를 첨가하고 암소에서 30분 동안 보관한 후에 흡광도(abs. 520 nm)를 측정하였다.
ABTS 라디칼 소거능은 Re 등(14)의 방법을 일부 수정하 여 수행하였다. ABTS(7.4 mM) 용액과 potassium persul- fate(2.6 mM) 용액을 1:1의 비율로 섞어 하루 동안 암소에 방치한 후, 혼합액의 흡광도 값(abs. 735 nm)이 1.4가 되도 록 메탄올로 희석하였다. 희석된 ABTS 용액 1 mL에 페놀 산 분획 추출액 20 μL를 첨가하고 암소에서 30분 동안 보관 한 후에 흡광도(abs. 735 nm)를 측정하였다. 항산화 실험의 활성은 Trolox equivalent antioxidant capacity(TEAC, mg TE/100 g sample)로 표현하였다.
통계분석
모든 실험은 3회 반복하였으며, 얻어진 측정값은 SPSS 통계 package program(Statistical Package for the Social Sciences, version 11.0, SPSS Inc., Chicago, IL, USA)을 이용하여 분산분석(ANOVA test)을 실시하였고, 다중범위 시험법(Duncan’s multiple range test)으로 유의성을 검증 하였다.
결과 및 고찰
세포벽 분해효소 처리 현미 가루의 유리 페놀산 함량
세포벽 분해효소 처리에 따른 현미 가루의 유리 페놀산의 조성 및 함량 변화는 Table 1에 나타냈다. 고성능 액체 크로 마토그래피로 분석한 결과 현미 가루에는 4-hydroxyben- zoic acid, vanillic acid, protocatechuic acid, syringic acid, caffeic acid,p
-coumaric acid, ferulic acid, sina- pinic acid 등의 유리 페놀산이 존재하였다. 이들 중에서p
- coumaric acid, ferulic acid는 현미 가루의 주된 페놀산으 로 전체 페놀산 함량의 약 60%를 차지하였다. 이와 같은 결과는 기존의 보고와 일치하였다(8,15). Butsat와 Siria- mornpun(15)은 유리 페놀산이 쌀의 배유가 아닌 미강에 주 로 존재하고 ferulic acid와p
-coumaric acid의 함량이 다 른 페놀산보다 높다고 하였으며, Tian 등(8)도 이와 유사한 결과를 보고하였다.Celluclast 1.5 L(cellulase)과 Pectinex Ultra SP-L (pectinase) 처리는 현미 가루의 페놀산 함량을 유의적으로 증가시켰다(Table 1). 증류수 처리 현미 가루(대조군)의 페 놀산 함량은 무처리군(현미 가루)과 유사하였다. 하지만 Celluclast 1.5 L과 Pectinex Ultra SP-L 처리군의 총 페놀 산 함량은 무처리군과 비교하여 각각 11.2배, 8.4배 높았으 며, 특히 ferulic acid 함량은 각각 16.2배, 10.2배 높았다.
이는 효소 처리에 의해 세포벽 구성성분과 결합하고 있는 불용성 페놀산이 떨어져 나와 유리형태로 전환되기 때문이 다. 현미 페놀산의 80~90%는 불용성 형태로 존재하는데 (7), 이들이 분해됨에 따라 유리 페놀산의 함량이 급격하게 증가한 것이다. Kim과 Lim(16)은 탄수화물 분해효소 처리 가 미강의 총 페놀산 함량을 2.5~3.0배 증가시켰으며, 특히 ferulic acid와
p
-coumaric acid의 함량이 두드러지게 증가 했다고 보고하였다. Moore 등(17)은 세포벽 가수분해효소 를 이용한 고체 발효가 밀기울(wheat bran)의 페놀산에 미 치는 영향을 조사하였으며, 세포벽 가수분해효소 처리에 의 해 밀기울의 불용성 페놀산 함량은 감소한 반면에 유리 페놀 산 함량은 증가했으며, 이는 효소 처리가 세포벽 구성요소와 불용성 페놀산 사이의 화학적 결합을 끊어주기 때문이라고Table 2. DPPH/ABTS radical scavenging activities and total phenolic content of the enzyme treated brown rice flours
Samples1) ABTS radical scavenging activity(mg TE/100 g sample) DPPH radical scavenging activity
(mg TE/100 g sample) TPC
(mg GAE/100 g sample) BRF
W-BRF C-BRF P-BRF
90.8±1.4c2) 89.1±1.4c 227.5±11.5a 153.0±13.0b
39.1±2.2c 36.3±2.2d 73.6±0.9a 53.2±1.5b
136.5±3.3c 132.4±6.3c 232.6±11.6a 190.2±5.4b
1)BRF: brown rice flour, W-BRF: water treated BRF, C-BRF: Celluclast 1.5 L treated BRF, P-BRF: Pectinex Ultra SP-L treated
2)BRF.Values (dry weight basis) with different letters (a-d) within a column are different with statistical significance (P<0.05).
보고하였다. Alrahmany 등(18)은 세포벽 분해효소 처리가 귀리 겨(oat bran)의 유리 페놀산 함량을 증가시킨다고 보고 하였다.
유리 페놀산 함량의 증가는 세포벽 분해효소의 작용에 의 한 것만은 아니다. 세포벽 분해효소가 단일로 작용할 경우 세포벽 구성성분을 분해할 수 있지만 페놀산-세포벽 구성성 분의 에스터 결합을 끊을 수 없다. 세포벽 구성성분과 에스 터 결합을 하고 있는 페놀산을 유리형태로 바꾸기 위해서는 phenolic acid esterase가 필요하다. Phenolic acid ester- ase의 활성이 없으면 불용성 페놀산은 완전히 유리되지 못 하고 당류가 붙어있는 conjugated form으로 세포벽에서 분 리될 것이다. Xue 등(19)은 다양한 미생물에서 분리된 cel- lulase 처리가 볏짚(rice straw)의 유리 페놀산 조성에 미치 는 영향을 조사하였으며, feruloyl esterase 활성을 가진 효 소가 유리페놀산 함량을 증가시키는 데 효과적이라고 하였 다. 본 실험에 사용된 Celluclast 1.5 L과 Pectinex Ultra SP-L은 phenolic acid esterase 활성을 가지고 있는 것으 로 보고되었다(20-22). Donaghy와 McKay(22)는 상업적 으로 이용되는 탄수화물 분해효소 10종의 phenolic acid esterase 활성을 분석한 결과, Pectinex Ultra SP-L에서 그 활성이 가장 높다고 하였다.
미생물에서 분리 및 정제된 상용 세포벽 분해효소는 phe- nolic acid esterase 활성을 가지고 있기 때문에(20-23), 본 실험에서 사용된 두 효소(Pectinex Ultra SP-L, Cellu- clast 1.5 L) 외의 다른 상용 세포벽 분해효소 또한 현미 가루의 페놀산 함량 및 기능성을 증진시킬 수 있을 것이라 생각되며, 이에 대한 추가 연구가 필요할 것이다.
세포벽 분해효소 처리 현미 가루의 총 페놀 화합물 함량
세포벽 분해효소 처리에 따른 현미 가루의 총 페놀 화합물 함량은 Folin-Ciocalteu 법으로 측정되었으며, 결과는 Table 2에 나타냈다. 증류수 처리 현미 가루(대조군)의 총페놀 화 합물 함량은 무처리군(현미 가루)보다 약간 감소하였지만 유의적인 차이를 보이지 않았다. 현미 가루의 총 페놀 화합 물 함량은 136.5 mg GAE/100 g으로 기존 보고들과 유사하 였다(24-27). Kim 등(24)은 한국에서 재배되는 7품종 현미 의 총 페놀 화합물 함량을 분석한 결과 유색미인 흑진주와 흑광의 총 페놀 화합물 함량은 다른 품종보다 약 4배 이상 높았지만, 다른 품종(고아미4호, 일품, 큰눈벼, 하이아미)의총 페놀 화합물 함량은 약 100~130 mg GAE/100 g으로 유사하였다. 현미의 페놀 화합물은 미강과 배유에 주로 존재 하는 것으로 알려져 있다(24-26). 현미의 총 페놀 화합물 함량은 백미보다 4배 이상 높으며 이는 페놀 화합물이 페놀 산과 마찬가지로 배유가 아닌 미강과 배아에 편중되어 있기 때문이다(25). 페놀 화합물은 자유 라디칼(free radical)을 안정화시킬 수 있는 페놀 고리(phenolic ring)를 가지고 있 기 때문에 뛰어난 항산화 활성을 가진다. 따라서 백미보다 총 페놀 화합물 함량이 높은 현미를 취반밥 및 쌀 가공식품 에 이용하는 것이 국민의 건강 유지에 긍정적일 것이다. 또 한, 쌀밥 및 가공식품은 호화 후 섭취하는 것이 일반적인데, 비록 호화에 의해 현미의 총 페놀 화합물 함량은 감소하지만 여전히 백미와 비교해서 그 함량이 여전히 높았다(25).
세포벽 분해효소 처리는 현미 가루의 총 페놀 화합물 함량 을 유의적으로 증가시켰다. Celluclast 1.5 L과 Pectinex Ultra SP-L 처리에 따라 총 페놀 화합물 함량은 각각 1.7배, 1.4배 증가하였다. 비록 효소 처리에 따른 증가 경향은 앞서 살펴본 유리 페놀산 함량과 달랐지만(Table 1), 총 페놀 화 합물 함량도 효소 처리에 의해 유의적으로 증가하였다. 이는 세포벽의 구성성분과 결합하고 있는 페놀산이 세포벽 분해 효소 처리에 의해 추출하기 쉬운 형태로 전환되었기 때문으 로 생각된다. 효소에 의해 불용성 페놀산은 conjugated phenolic acid로 전환될 것이며, 물 및 에탄올에 대한 용해 도가 증가할 것이다. 저분자 탄수화물과 페놀 화합물이 결합 된 형태의 페놀산을 conjugated phenolic acid라고 하는데, 효소 처리에 따른 conjugated phenolic acid 구성 및 함량 변화에 대한 연구는 아직 진행되지 않고 있으며 이에 대한 연구가 진행되면 좀 더 명확하게 효소 처리에 따른 페놀 화 합물 변화 메커니즘을 구명할 수 있을 것이다.
또한, 이에 대한 간접적인 증거로는 효소 처리에 따른 추 출 효율 및 총당을 측정함으로써 알 수 있을 것이다. Kim과 Lim(16)은 Celluclast 1.5 L과 Pectinex Ultra SP-L을 포 함한 6종의 효소 처리가 탈지 미강 추출물의 추출효율 및 총당, 환원당, 총 페놀 화합물 함량을 유의적으로 증가시킨 다고 보고하였다. 이는 세포벽 분해효소 처리에 의해 고분자 탄수화물 및 불용성 페놀산이 유리화됨을 의미한다. 볏짚 (rice straw)과 밀(wheat)에서도 이와 유사한 결과가 보고 되었다(19,27). 또한, 앞서 살펴본 것과 같이 phenolic acid esterase에 의한 단순 페놀산 함량 증가도 총 페놀 화합물의
Table 3. Glutamic acid and GABA content in enzyme treated
brown rice floursSamples1) Glutamic acid
(mg/100 g sample) GABA (mg/100 g sample) BRF
W-BRF C-BRF P-BRF
11.76±0.10a2) 4.84±0.12b 3.26±0.04d 3.82±0.09c
0.76±0.01d 6.15±0.23c 32.71±1.31a 22.40±0.90b
1)BRF: brown rice flour, W-BRF: water treated BRF, C-BRF:
Celluclast 1.5 L treated BRF, P-BRF: Pectinex Ultra SP-L treated BRF.
2)Values (dry weight basis) with different letters (a-d) within a column are different with statistical significance (P<0.05).
함량 증가에 영향을 미칠 것이다. 효소 처리에 따라 유리페 놀산 함량과 총 페놀 화합물 함량은 모두 증가하였지만, 두 값 사이에서 정의 상관관계를 관찰할 수 없었다(Table 1, Table 2). 이는 단순 페놀산, conjugated phenolic acid뿐 만 아니라 쌀에는 탄닌(tannin), 플라보노이드(flavonoid) 등 다양한 페놀 화합물들이 존재하기 때문이다(28,29).
또한, 총 페놀 화합물 함량의 증가는 실험에서 효소 처리 온도에 영향을 받을 수 있다. 열처리는 열에 약한 항산화 물질의 함량을 떨어트릴 수도 있지만, 용매에 의한 추출효율 을 높일 수도 있기 때문이다. Kwak 등(28)은 열처리 온도에 따른 현미 에탄올 추출물의 총 페놀 화합물 함량을 조사하였 다. 그들은 40°C 처리가 현미의 추출효율 및 총 페놀 화합물 함량을 증가시키며, 열처리에 의한 총 페놀 화합물 함량 변 화는 식품을 구성하고 있는 불용성 페놀산의 열반응성에 따 라 달라질 수 있다고 하였다.
세포벽 분해효소 처리 현미 가루의 GABA 함량
세포벽 분해효소 처리에 따른 GABA 함량 변화는 Table 3에 나타냈다. 현미 가루의 GABA 함량은 0.76 mg/100 g으 로 기존 보고와 유사하였다. 기존 연구에 따르면 현미의 GABA 함량은 1.4~54.2 mg/100 g으로 보고되었다(4). Lee 등(30)은 한국에서 생산되는 현미 10품종(삼광, 큰눈, 홍진 주, 흑광, 수원595호, 미소미, 고아미4호, 단미, 친들, 현품) 의 GABA 함량을 조사한 결과 0.07~28.24 mg/100 g 범위 이며, 이는 품종, 재배 환경 등 다양한 요소에 영향을 받는다 고 하였다. Saikusa 등(31)은 현미의 배아, 미강, 배유의 GABA 함량이 각각 25.4, 15.9, 3.0 mg/100 g으로 현미의 GABA는 배아와 미강에 편중되어 있다고 하였다.
GABA는 비단백계 아미노산의 하나로 포유류 대뇌피질 에서 억제성 신경전달물질로 작용하여 뇌세포 대사 기능 촉 진, 신경 안정작용, 스트레스 완화, 기억력 증진, 우울증 완 화, 기억력 증진 등의 다양한 효과가 있기 때문에(32), 현미 및 쌀 부산물에서 GABA 함량을 증진시키고자 하는 연구가 다양하게 진행되었다. 발아(germination)는 현미의 GABA 함량을 증가시킬 수 있는 가장 효과적인 방법으로 그동안 많이 연구되었다(4). Ohtsubo 등(33)은 발아에 의해 현미의 GABA 함량이 25배 증가하였다고 보고하였다. 또한, 발아
침지 용액에 글루탐산 및 키토산을 첨가하면 GABA 함량 증진에 효과적인 것으로 보고되었다(34). Kim 등(35)은 전 해수 처리가 미강의 GABA 함량을 증진시킨다고 보고하였 다. 이외에도 다양한 스트레스 처리(기계적 상처, 열처리, 혐기처리 등)가 식물체의 GABA 함량을 증진시키는 데 효과 적인 것으로 보고되었다(36).
증류수 처리 현미 가루(대조군)의 글루탐산과 GABA 함 량은 각각 4.84, 6.15 mg/100 g으로 무처리군(현미 가루)과 비교하여 글루탐산의 함량은 약 2배가량 감소한 반면에 GABA 함량은 약 8배 증가하였다. 이는 현미 자체가 가지고 있는 glutamate decarboxylase(GAD)가 증류수 처리에 의 해 활성화되었기 때문으로 생각된다. GAD는 L-glutamate 의 탈탄산화를 유도하여 GABA를 생성하는 효소이다. GAD 는 현미의 미강과 배아에 존재하며 GAD 활성화에 적합한 조건을 갖추면 간단하게 GABA 생합성을 유도할 수 있다 (37,38). Kim 등(35)은 미강을 40°C의 물에 침지하는 것만 으로 GABA 함량을 9배 이상 증가시킬 수 있다고 보고하였 다. Ohtsubo 등(37)은 GABA의 전구체인 glutamate를 인 위적으로 공급해주면 100 g의 배아로 29 g의 GABA를 합성 할 수 있다고 하였다.
Celluclast 1.5 L과 Pectinex Ultra SP-L 처리 현미 가루 의 GABA 함량은 각각 32.7, 22.4 mg/100 g으로 대조군보 다 각각 43.0, 29.5배 높았다(Table 3). 반면 GABA 생합성 의 전구체인 glutamic acid의 함량은 효소 처리에 의해 급격 하게 감소하였다. 효소 처리에 따라 GABA 함량이 유의적으 로 증가한 것은 현미 자체가 가지고 있는 glutamate de- carboxylase(GAD)와 외부에서 넣어준 세포벽 분해효소의 복합적 작용에 의한 것으로 생각된다. 효소 처리는 세포벽의 연화 및 파괴를 유도하여 수분흡수 및 GAD 효소 작용을 촉진할 수 있을 것으로 생각된다. 상업적으로 이용 및 시판 되는 효소 제품은 단일의 효소로 존재하는 것이 아니라 주요 작용을 하는 효소와 함께 일부 다른 작용을 하는 효소가 혼 입되어 있을 수 있다. Celluclast 1.5 L과 Pectinex Ultra SP-L에 단백질 분해효소가 일부 혼입되어 있다면 이들이 현미 단백질을 분해하여 GABA 전구체인 glutamic acid를 형성할 것이다.
세포벽 분해효소 처리 현미 가루의 항산화 활성
항산화 활성 측정에는 ABTS 라디칼 소거능과 DPPH 라 디칼 소거능 측정법이 사용되었으며, 그 결과는 Table 2에 나타냈다. 증류수 처리는 현미 가루의 항산화 활성에 큰 영 향을 미치지 못한 것과 대조적으로 Celluclast 1.5 L과 Pec- tinex Ultra SP-L 처리는 현미 가루의 항산화 활성을 유의적 으로 증가시켰다. Celluclast 1.5 L과 Pectinex Ultra SP-L 처리군의 ABTS 라디칼 소거능은 각각 227.5, 153.0 mg TE/100 g sample로 대조군(90.8 mg TE/100 g sample)과 비교하여 2.5배, 1.7배 높았으며, DPPH 라디칼 소거능은 각각 73.6, 53.2 mg TE/100 g sample로 대조군(39.1 mg
TE/100 g sample)과 비교하여 1.9배, 1.4배 높았다. 효소 처리에 따른 항산화 활성의 증가는 유리페놀산 및 총 페놀 화합물 함량 증가(Table 1, Table 2)에 기인한다고 생각된 다. 페놀 화합물은 수소 공여능, 금속 킬레이팅, 자유 라디칼 소거 등의 효과를 가지고 있으며, 쌀을 비롯한 다양한 곡류 의 주요 항산화 물질로 알려져 있다(39,40). 또한, 효소 처리 가 항산화 물질의 추출 효율을 증가시켰기 때문일 수 있다.
가수분해효소 처리는 용매에 대한 고형분 추출 효율을 높인 다(10,11,16).
현미 가루의 항산화 활성은 효소 처리에 의해 증가하였지 만 페놀산 및 총 페놀 화합물 함량과 뚜렷한 상관관계를 가 지고 있지 않았다(Table 1, Table 2). 이는 페놀 화합물 외 의 다양한 항산화 물질이 현미 가루에 존재하기 때문으로 생각된다. 쌀에는 페놀 계열 화합물(페놀산, 플라보노이드, 탄닌, 안토시아닌 등)뿐만 아니라 다양한 항산화 물질(γ- oryzanol, tocopherol, tocotrienol, phytic acid 등)이 있다 (4). 이들의 항산화력은 비록 페놀산에 비해 떨어지지만 그 들의 함량은 페놀 화합물과 비교될 정도로 많은 양이 현미에 존재한다(40). Goufo와 Trindade(40)는 현미에 페놀 화합 물, tocols 및 γ-oryzanol이 각각 102.6 mg/kg, 60.18 mg/
kg, 413.3 mg/kg 존재하며, 페놀 화합물이 현미에서 항산화 력을 나타내는 주성분이기는 하지만 다른 항산화 물질 (tocols, γ-oryzanol, phytic acid 등)도 현미의 항산화력을 결정하는 데 중요한 역할을 한다고 하였다.
요 약
쌀은 전 세계에서 주요한 에너지 공급원으로 이용되고 있는 식량작물로, 특히 우리나라 국민에게 있어 매우 중요한 식량 작물 중 하나이다. 하지만 최근 서구화된 식문화와 함께 국 민 1인당 쌀 소비량은 점차 감소하고 있는 추세이며, 이를 해결하고자 쌀 가공품 및 특수미 품종 개발 등의 다양한 연 구가 진행되고 있다. 특히 2000년 이후의 웰빙 트렌드와 함께 건강 기능성 식품에 대한 관심이 증가함에 따라 천연 유래의 건강기능성 식품 소재에 대한 관심이 증가하였다.
따라서 우리나라 국민들이 가장 많이 섭취하는 식량작물인 쌀의 영양성 및 건강기능성에 대한 관심 및 요구도 커질 수 밖에 없어졌다. 본 연구에서는 이런 요구를 충족할 수 있는 쌀 2차 가공품 개발을 위하여 건강기능성이 향상된 현미 가 루를 개발하고자 하였다. 효소 처리는 세포벽 구성성분과 강하게 결합하고 있는 페놀산을 유리화하여 현미 가루의 유 리 페놀산 및 총 페놀 화합물 함량을 유의적으로 증가시켰 다. 항산화력 또한 효소 처리에 의해 유의적으로 증가하였 다. 하지만 페놀 화합물 함량과 항산화 활성 사이에는 뚜렷 한 상관관계가 없었으며, 이는 현미에는 페놀 화합물 이외의 다양한 항산화 물질이 존재하기 때문이다. 효소 처리에 의해 GABA 함량도 유의적으로 증가하였는데, 이는 현미에 있는 glutamate decarboxylase(GABA 합성 효소)가 세포벽 분
해효소 처리조건에서 활성화되기 때문이다. 세포벽 분해효 소 처리는 친환경적이고 간편한 처리 방법임에도 불구하고 현미의 생리활성 물질 함량 및 항산화 활성을 높이는 데 효 과적이었다. 따라서 앞으로 효소 처리 현미 가루의 가공적성 평가 및 생산 공정 최적화에 대한 연구가 심도 깊게 이루어 진다면 쌀가루의 상업적 이용 및 소비에 기여할 수 있을 것 으로 생각된다.
REFERENCES
1. FAO Rice Market Monitor (RMM). http://www.fao.org/eco nomic/est/publications/rice-publications/rice-market-mon itor-rmm/en/ (accessed Oct 2017).
2. Park JW, Chae S, Yoon S. 2009. The effects of steeping and cooking pressure on qualities of cooked brown rice. Korean
J Food Cult 24: 69-76.
3. Oh SK, Kim DJ, Cheun AR, Yoon MR, Hong HC, Choi IS, Oh YJ, Oh KB, Kim YK. 2010. Quality evaluation of
Juanbyeo as aseptic-packaged cooked rice. Korean J Food Sci Technol 42: 721-726.
4. Cho DH, Lim ST. 2016. Germinated brown rice and its bio- functional compounds. Food Chem 196: 259-271.
5. Heleno SA, Martins A, Queiroz MJ, Ferreira IC. 2015.
Bioactivity of phenolic acids: metabolites versus parent com- pounds: A review. Food Chem 173: 501-513.
6. Lopez-Martinez LX, Oliart-Ros RM, Valerio-Alfaro G, Lee CH, Parkin KL, Garcia HS. 2009. Antioxidant activity, phe- nolic compounds and anthocyanins content of eighteen strains of Mexican maize. LWT
- Food Sci Technol 42: 1187-1192.
7. Zhou Z, Robards K, Helliwell S, Blanchard C. 2004. The distribution of phenolic acids in rice. Food Chem 87: 401- 406.
8. Tian S, Nakamura K, Kayahara H. 2004. Analysis of phenolic compounds in white rice, brown rice, and germinated brown rice. J Agric Food Chem 52: 4808-4813.
9. Heredia A, Jiménez A, Guillén R. 1995. Composition of plant cell walls. Z Lebensm Unters Forsch 200: 24-31.
10. Hourigan AJA, Chesterman CF. 1997. Application of carbo- hydrases in extracting protein from rice bran. J Sci Food
Agric 74: 141-146.
11. Hanmoungjai P, Pyle DL, Niranjan K. 2002. Enzyme-as- sisted water-extraction of oil and protein from rice bran.
J Chem Technol Biotechnol 77: 771-776.
12. Adom KK, Liu RH. 2002. Antioxidant activity of grains.
J Agric Food Chem 50: 6182-6187.
13. Bondet V, Brand-Williams W, Berset C. 1997. Kinetics and mechanisms of antioxidant activity using the DPPH・free radical method. LWT
- Food Sci Technol 30: 609-615.
14. Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice- Evans C. 1999. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol
Med 26: 1231-1237.
15. Butsat S, Siriamornpun S. 2010. Antioxidant capacities and phenolic compounds of the husk, bran and endosperm of Thai rice. Food Chem 119: 606-613.
16. Kim SM, Lim ST. 2016. Enhanced antioxidant activity of rice bran extract by carbohydrase treatment. J Cereal Sci 68: 116-121.
17. Moore J, Cheng Z, Su L, Yu LL. 2006. Effects of solid-state enzymatic treatments on the antioxidant properties of wheat bran. J Agric Food Chem 54: 9032-9045.
18. Alrahmany R, Avis TJ, Tsopmo A. 2013. Treatment of oat bran with carbohydrases increases soluble phenolic acid con- tent and influences antioxidant and antimicrobial activities.
Food Res Int 52: 568-574.
19. Xue Y, Wang X, Chen X, Hu J, Gao MT, Li J. 2017. Effects of different cellulases on the release of phenolic acids from rice straw during saccharification. Bioresour Technol 234:
208-216.
20. Faulds CB, Sancho AI, Bartolomé B. 2002. Mono- and di- meric ferulic acid release from brewer’s spent grain by fun- gal feruloyl esterases. Appl Microbiol Biotechnol 60: 489- 494.
21. Spanikova S, Biely P. 2006. Glucuronoyl esterase-Novel carbohydrate esterase produced by Schizophyllum commune.
FEBS Lett 580: 4597-4601.
22. Donaghy JA, McKay AM. 1994. Novel screening assay for the detection of phenolic acid esterases. World J Microbiol
Biotechnol 10: 41-44.
23. Di Gioia D, Sciubba L, Setti L, Luziatelli F, Ruzzi M, Zani- chelli D, Fava F. 2007. Production of biovanillin from wheat bran. Enzyme Microb Technol 41: 498-505.
24. Kim DJ, Oh SK, Yoon MR, Chun AR, Choi IS, Lee DH, Lee JS, Yu KW, Kim YK. 2011. The change in biological activities of brown rice and germinated brown rice. J Kore-
an Soc Food Sci Nutr 40: 781-789.
25. Kim SY, Seo BY, Park E. 2013. The impact of cooking on the antioxidative and antigenotoxic effects of rice. J Korean
Soc Food Sci Nutr 42: 1370-1377.
26. Kim DJ, Oh SK, Yoon MR, Chun AR, Hong HC, Lee JS, Kim YK. 2010. Antioxidant compounds and antioxidant ac- tivities of the 70% ethanol extracts from brown and milled rice by cultivar. J Korean Soc Food Sci Nutr 39: 467-473.
27. Sorensen HR, Meyer AS, Pedersen S. 2003. Enzymatic hy- drolysis of water-soluble wheat arabinoxylan. 1. Synergy be- tween α-L-arabinofuranosidases, endo-1,4-β-xylanases, and β-xylosidase activities. Biotechnol Bioeng 81: 726-731.
28. Kwak J, Oh SK, Kim DJ, Lee JH, Yoon MR, Kim HW, Lee JS. 2013. Effects of heat-treated brown rice on total phenolics and antioxidant activities. J Korean Soc Food Sci
Nutr 42: 534-541.
29. Oh SK, Choi HC, Cho MY, Kim SU. 1996. Extraction
method of anthocyanin and tannin pigments in colored rice.
Agric Chem Biotechnol 39: 327-331.
30. Lee K, Ham H, Kim HJ, Park HY, Sim EY, Oh SK, Kim WH, Jeong HS, Woo KS. 2016. Functional components and radical scavenging activity of germinated brown rice accord- ing to variety. Korean J Food Nutr 29: 145-152.
31. Saikusa T, Horino T, Mori Y. 1994. Distribution of free amino acids in the rice kernel and kernel fractions and the effect of water soaking on the distribution. J Agric Food
Chem 42: 1122-1125.
32. Mody I, De Koninck Y, Otis TS, Soltesz I. 1994. Bridging the cleft at GABA synapses in the brain. Trends Neurosci 17: 517-525.
33. Ohtsubo KI, Suzuki K, Yasui Y, Kasumi T. 2005. Bio-func- tional components in the processed pre-germinated brown rice by a twin-screw extruder. J Food Compos Anal 18:
303-316.
34. Oh SH, Lee IT, Park KB, Kim BJ. 2002. Changes in the levels of water soluble protein and free amino acids in brown rice germinated in a chitosan/glutamic acid solution.
Korean J Biotechnol Bioeng 17: 515-519.
35. Kim HS, Lee EJ, Lim ST, Han JA. 2015. Self-enhancement of GABA in rice bran using various stress treatments. Food
Chem 172: 657-662.
36. Kinnersley AM, Turano FJ. 2000. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit Rev Plant Sci 19: 479-509.
37. Ohtsubo S, Asano S, Sato K, Matsumoto I. 2000. Enzymatic production of γ-aminobutyric acid using rice (Oryza sativa) germ. Food Sci Technol Res 6: 208-211.
38. Wang L, Xu DX, Lv YG, Zhang H. 2010. Purification and biochemical characterisation of a novel glutamate decarbox- ylase from rice bran. J Sci Food Agric 90: 1027-1033.
39. Balasundram N, Sundram K, Samman S. 2006. Phenolic compounds in plants and agri-industrial by-products: Antiox- idant activity, occurrence, and potential uses. Food Chem 99: 191-203.
40. Goufo P, Trindade H. 2014. Rice antioxidants: phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci Nutr 2:
75-104.