기후변화에 따른 한반도 난온대 상록활엽수의 잠재 생육지 변화 예측
1a윤종학2*․中尾勝洋3․박찬호2․이병윤2․오경희2
Change Prediction for Potential Habitats of Warm-temperate Evergreen Broad-leaved Trees in Korea by Climate Change
1aJong-Hak Yun2*, Katsuhiro Nakao3, Chan-Ho Park2, Byoung-Yoon Lee2, Kyoung-Hee Oh2
요 약
본 연구는 기후변화에 따른 한반도 난온대 상록활엽수의 생육지 변화를 예측하기 위하여 CT-model을 이용하여 현재기후(1961~1990)와 3종류의 미래기후(2081~2100) 시나리오에서의 잠재 생육지를 예측하였다. 반응변수로서 난 온대 상록활엽수의 실제 분포에서 추출한 유/무자료와 4가지 기후변수(온량지수, 최한월최저기온, 동경강수량, 하계강 수량)를 예측변수로 사용하였다. 현재기후에서 잠재 생육지(PH)는 28,230km2로 예측되었으며, 3종류 미래기후 시나리 오(CCCMA-A2, CSIRO-A2, HADCM3-A2)에서는 77,140~89,285km2로 예측되었다. 현재기후에서 토지 이용을 고려 한 잠재 생육지(PHLU)는 8,274km2로 예측되었으며, 잠재 생육지의 29.3%를 차지하였다. 미래기후에서 토지 이용을 고려한 잠재 생육지는 35,177~45,170km2로 예측되었으며, 26.9~36.9% 증가하였다. 기후변화에 따른 난온대 상록활엽 수의 분포 확대는 토지 이용에 제한되어 생육지 파편 형태로 진행되고 있다. 난온대 상록활엽수의 생육지 증가는 난온대 낙엽활엽수림과의 경쟁이 예상되며, 난온대 상록활엽수림대의 확대 및 북상을 시사하고 있다.
주요어: CT모델, 토지 이용, 생육지 파편
1 접수 2011년 4월 5일, 수정(1차: 2011년 6월 21일, 2차: 2011년 7월 6일), 게재확정 2011년 7월 7일 Received 5 April 2011; Revised(1st: 21 June 2011, 2nd: 6 July 2011); Accepted 7 July 2011
2 국립생물자원관 식물자원과 Plant Research Division, National Institute of Biological Resource, Incheon(404-708), Korea 3 일본 산림총합연구소 식물 생태학 연구실 Department of Plant Ecology, Forestry and Forest Products Research Institute,
Tsukuba, Ibaraki(305-8687), Japan
a 이 논문은 2011년도 환경부 기후변화 적응 생물 다양성 관리 연구 조성비에 의하여 연구되었음.
* 교신저자 Corresponding author([email protected])
ABSTRACT
The research was carried out for prediction of the potential habitats of warm-temperate evergreen broad-leaved trees under the current climate(1961~1990) and three climate change scenario(2081~2100) (CCCMA-A2, CSIRO-A2 and HADCM3-A2) using classification tree(CT) model. Presence/absence records of warm-temperate evergreen broad-leaved trees were extracted from actual distribution data as response variables, and four climatic variables (warmth index, WI; minimum temperature of the coldest month, TMC;
summer precipitation, PRS; and winter precipitation, PRW) were used as predictor variables. Potential habitats(PH) was predicted 28,230㎢ under the current climate and 77,140~89,285km2 under the three climate change scenarios. The PH masked by land use(PHLU) was predicted 8,274km2 and the proportion of PHLU within PH was 29.3% under the current climate. The PH masked by land use(PHLU) was predicted 35,177~45,170km2 and increased 26.9~36.9% under the three climate change scenarios. The expansion of warm-temperate evergreen broad-leaved trees by climate change progressed habitat fragmentation by
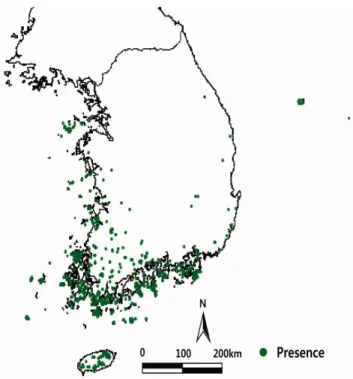
Figure 1. Actual distribution of warm-temperate evergreen broad-leaved trees(1913-2009)
restriction of land use. The habitats increase of warm-temperate evergreen broad-leaved trees had been expected competitive with warm-temperate deciduous broadleaf forest and suggested the expand and northward shift of warm-temperate evergreen broad-leaved forest zone.
KEY WORDS: CLASSIFICATION TREE MODEL, LAND USE, HABITAT FRAGMENTATION,
서 론
향후 100년간 지구의 온도는 1.8℃〜4.0℃ 상승할 것으 로 예측되고 있으며, 이러한 온도 상승은 생태계 및 생물 생육지 등에 상당한 영향을 미칠 것으로 예상되고 있다 (IPCC, 2007). 지구 온난화에 의한 식물 분포에 관한 예측 연구는 1990년대부터 유럽과 미국을 중심으로 활발히 진행 되어 왔다(Huntley et al., 1995; Iverson et al., 1998;
Leathwick et al, 1996; Sykes et al., 1996). 식물종이 생태 적으로 생육에 적합한 환경 조건 지역에 분산되어 분포하고 있을 경우, 식물종의 분포를 환경 조건에 따른 예측 모델을 개발함으로써 현재의 생육에 적합한 생육 적지(Suitable habitat) 및 생육이 가능한 잠재 생육지(PH: Potential habitat)를 예측할 수 있다. 이렇게 구축된 식물종의 분포 예측 모델에 미래기후 조건을 적용하여 미래기후에 따른 생육 적지 및 잠재 생육지를 예측할 수 있다. 생육지 예측 연구는 기후변화에 취약한 식물종과 취약 지역, 기후변화에 도 불구하고 계속해서 생육이 가능한 지역(도피지, Refugia)을 특정 할 수 있으며, 지구 온난화가 진행되는 21 세기에 식물군락과 야생식물의 보전 관리계획에 크게 활용 될 수 있다. 미국에서는 동부지역에 분포하는 주요수종 80 종(Iverson et al. 1999), 영국에서는 33종의 식물(Harrison et al., 2001; Berry et al., 2003)에 대해서 현재 및 미래기후 에서의 잠재 생육지를 예측하였다. 유럽에서 식물종 1000 종 이상의 잠재 생육지를 예측하여 기후변화에 따른 식물종 의 다양성에 대한 평가에 대하여 보고하였다(Bakkenes et al., 2002; Thuiller et al., 2005). 일본에서는 기후요인에 의한 자연 식생대, 5개 산림형, Fagus crenata林(일명: 부나 림), 상록침엽수 등의 잠재 생육지 예측과 온난화 영향 평가 연구보고(Uchijima et al., 1992; Tsunekawa et al., 1993;
Matsui et al., 2004a; 2004b, Tanaka et al., 2009)가 있으나, 한국에서는 기후변화에 따른 한반도의 식물종에 대한 분포 예측 연구는 활발히 진행되고 있지 않다.
본 연구는 종분포 모델을 이용하여 구축된 한반도 난온대 상록활엽수 분포 예측 모델(EG-model)(Yun et al., 2011)의 생육지에 대한 기후조건을 토대로 기후변화에 따른 잠재 생육지를 예측하고자 하였다.
재료 및 방법
1. 식물분포 자료
식물분포 자료는 한반도 난온대 상록활엽수 57종(Table 1)을 대상으로 국립생물자원관(KB), 국립수목원(KH), 고 려대학교(KUS), 경북대학교(KNU), 대전대학교(TUT), 서 울대학교(SNU), 순천대학교(SNUH), 영남대학교(YNUH), 인하대학교(IUI), 전남대학교(CNU), 전북대학교(JNU), 한 남대학교(HNHM)에 소장되고 있는 1913~2009년을 수집 표본 기간으로 하는 표본 4,670점의 위치정보 자료(Yun et al., 2011)를 이용하였다(Figure 1). 식물종 57종의 학명과 국명은 Flora of Korea Editorial Committee(2007)의 The genera of Vascular Plant of Korea에 준하여 작성하였다 (Table 1).
Family name Species name Period of collection Scientific name / Korean name
Lauraceae 녹나무과
1. Actinodaphne lancifolia (Siebold & Zucc.) Meisn. 육박나무 1953-2007
2. Cinnamomum camphora (L.) Siebold 녹나무 1924-2008
3. Cinnamomum japonicum Siebold 생달나무 1955-2009
4. Litsea japonica (Thunb.) Juss. 까마귀쪽나무 1953-2008
5. Machilus japonica Siebold & Zucc. 센달나무 1953-2008
6. Machilus thunbergii Siebold & Zucc. 후박나무 1936-2009
7. Neolitsea aciculata (Blume) Koidz. 새덕이 1984-2009
8. Neolitsea sericea (Blume) Koidz. 참식나무 1952-2009
Chloranthaceae 홀아비꽃대과
9. Chloranthus glaber (Thunb.) Nakai 죽절초 1991-2005
Piperaceae 후추과
10. Piper kadzura (Choiy) Ohwi 후추등 1978-2007
Illiciaceae 붓순나무과
11. Illicium anisatum L. 붓순나무 1955-2008
Schisandraceae 오미자과
12. Kadsura japonica (L.) Dunal 남오미자 1921-2008
Lardizabalaceae 으름덩굴과
13. Stauntonia hexaphylla (Thunb.) Decne. 멀꿀 1921-2009
Hamamelidaceae 조록나무과
14. Distylium racemosum Siebold & Zucc. 조록나무 1991-2008 Daphniphyllaceae 굴거리나무과
15. Daphniphyllum macropodum Miq. 굴거리나무 1935-2008
16. Daphniphyllum teijsmannii Kurz ex Teijsm. & Binn. 좀굴거리나무 1972-2009 Moraceae 뽕나무과
17. Ficus oxyphylla Miq. ex Zoll. 모람 1919-2008
18. Ficus thunbergii Maxim. 왕모람 1954-1972
Myricaceae 소귀나무과
19. Myrica rubra Siebold & Zucc. 소귀나무 1991
Fagaceae 참나무과
20. Castanopsis cuspidata (Thunb.) Schottky 모밀잣밤나무 1971-2009
21. Quercus acuta Thunb. 붉가시나무 1915-2009
22. Quercus gilva Blume 개가시나무 1991-2008
23. Quercus glauca Thunb. 종가시나무 1978-2009
24. Quercus myrsinaefolia Blume 가시나무 1956-2007
25. Quercus salicina Blume 참가시나무 1913-2007
Theaceae 차나무과
26. Cleyara japonica Thunb. 비쭈기나무 1969-2003
27. Eurya emarginata (Thunb.) Makino 우묵사스레피나무 1962-2009
28. Eurya japonica Thunb. 사스레피나무 1935-2009
29. Ternstroemia gymnanthera (Wight & Arn.) Sprague 후피향나무 1935-2009 Elaeocarpaceae 담팔수과
30. Elaeocarpus sylvestris var. elliptcus (Thunb.) H. Hara 담팔수 1991-2009 Flacourtiaceae 산유자나무과
31. Xylosma congestum (Lour.) Merr. 산유자나무 1955-2009
Ericaceae 진달래과
32. Vaccinium bracteatum Thunb. 모새나무 1918-2008
Symplocaceae 노린재나무과
33. Symplocos prunifolia Siebold & Zucc. 검은재나무 2008
Table 1. Warm-temperature evergreen broad-leaved trees
Family name Species name Period of collection Scientific name / Korean name
Myrsinaceae 자금우과
34. Ardisia crenata Sims 백량금 1958-2008
35. Ardisia japonica (Thunb.) Blume 자금우 1935-2009
36. Ardisia pusilla A. DC. 산호수 1971-2009
Pittosporaceae 돈나무과
37. Pittosporum tobira (Thunb.) W. T. Aiton 돈나무 1950-2008
Rosaceae 장미과
38. Rhaphiolepis indica var. umbellata (Thunb.) Ohashi, J. 다정큼나무 1951-2008
39. Rubus buergeri Miq. 겨울딸기 1984-2009
Elaeagnaceae 보리수나무과
40. Elaeagnus glabra Thunb. 보리장나무 1919-2009
41. Elaeagnus macrophllya Thunb. 보리밥나무 1928-2009
42. Elaeagnus maritima Koidz. 녹보리똥나무 1967-2008
43. Elaeagnus submacrophylla Servett. 큰보리장나무 1949-2008
Thymelaeaceae 팥꽃나무과
44. Daphne kiusiana Miq. 백서향나무 2001-2005
Aucubaceae 식나무과
45. Aucuba japonica Thunb. 식나무 1928-2009
Aqulfoliaceae 감탕나무과
46. Ilex cornuta Lindl. ex Paxton 호랑가시나무 1955-2006
47. Ilex integra Thunb. 감탕나무 1916-2009
Rutaceae 윤향과
48. Zanthoxylum planispinum Siebold & Zucc. 개산초 1971-2009 Araliaceae 두릅나무과
49. Dendropanax trifidus (Thunb.) Makino ex H. Hara 황칠나무 1941-2009 50. Fatsia japonica (Thunb.) Decne. & Planch. 팔손이 1999-2006
51. Hedera rhombea (Miq.) Bean 송악 1953-2009
Loganiaceae 마전과
52. Gardneria insularis Nakai 영주치자 1922-2003
Asclepiadaceae 박주가리과
53. Marsdenia tomentosa C. Morren & Decne. 나도은조롱 1969-2008 Oleaceae 물푸레나무과
54. Ligustrum japonicum Thunb. 광나무 1978-2008
55. Ligustrum lucidum W. T. Aiton 당광나무 1991-2006
Rubiaceae 꼭두선이과
56. Damnacanthus indicus C. F. Gaertn. 호자나무 1979-2009
Viburnaceae 산분꽃나무과
57. Viburnum odoratissimum var. awabuki (K. Koch) Zabel ex Rümpler 아왜나무 1935-2009 Table 1. (Continue)
2. 기후자료
기후 자료는 Worldclim에서 제공하는 Global Climate Data(www.worldclim.org)에서 30 arc-seconds(〜1km) 격 자에 대하여 한반도 지역의 기후값을 추출하여 식물 생육에 주요한 4개의 기후변수를 산출하여 적용하였다. 4개의 기후 변수는 식물 생육기의 열량 지표인 온량지수(WI: Warmth Index)(Kira, 1977), 동계의 저온 극값 지표인 최한월 최저
기온(TMC: Mean minimum temperature of the coldest month), 생육기의 수분공급 지표로서 5〜9월의 하계강수량 (PRS: Summer precipitation), 동계의 건조정도 및 적설량의 지표로서 12〜3월의 동계강수량(PRW: Winter precipitation) 을 이용하였다.
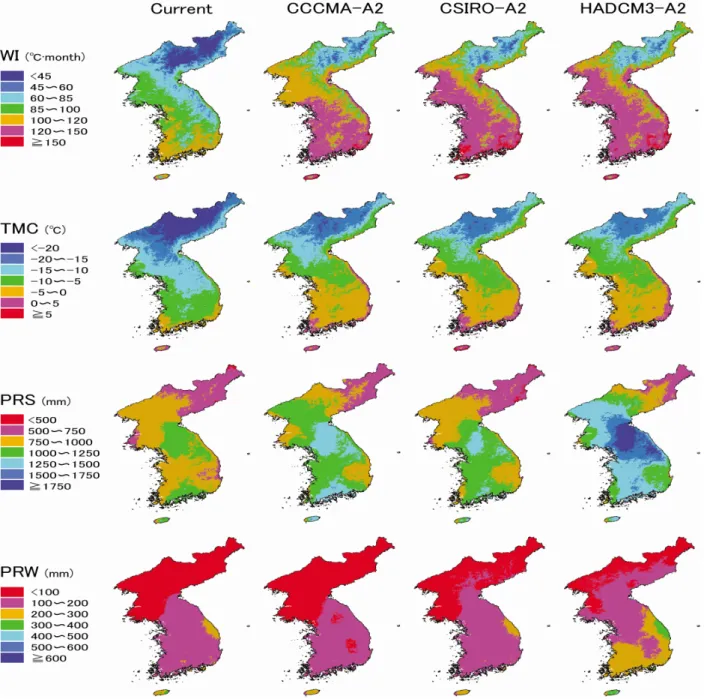
한반도 지역에 대한 현재기후 30년(1961∼1990년) 평균 기후값은 WI 95.0, TMC -.7.7℃, PRS 955.6mm PRW 157.mm 산출되었다. 미래기후는 2000년 IPCC(Intergovernmental
Figure 2. Maps of four climatic variables under the current climate(1961-1990) and future climate scenarios, i.e., the CCCMA-A2, CSIRO-A2 scenario and HADCM3-A2 scenario of 2081~2100 (WI: Warmth index, TMC: Mean minimum temperature of the coldest month, PRS: Summer(May-September) Panel on Climate Change)의 배출 시나리오에 관한 특별보
고서(SRES: Special Report on Emissions Scenario)에서 제 시한 A2(경제성장 및 지역주의 중심) 시나리오에 대하여 3가지 전지구 기후모델 CCCMA(Canadian Centre for Climate Modelling and Analysis), CSIRO(Commonwealth Scientifiec and Industrial Reserch Organisation), HADCM3 (Hadley Centre Coupled Model, version3)을 이용하여 예 측된 전지구 기후자료에서 한반도의 미래 기후값을 추출하
였다. 미래기후 시나리오에 의한 한반도 지역 평균 기후값 (2081~2100년)은 CCCMA-A2에서 WI 126.8, TMC -2.
6℃, PRS 1163.3mm, PRW 134.8mm, CSIRO-A2에서 WI 131.7, TMC -2.5℃, PRS 1093.0mm, PRW 148.8mm HADCM3-A2에서 WI 133.4, TMC -2.3℃, PRS 1477.8mm, PRW 220.1mm로 산출되었다. 연구 대상지역에 대한 4개의 기후변수를 약 1km 격자로 지도화하여 나타냈다(Figure 2).
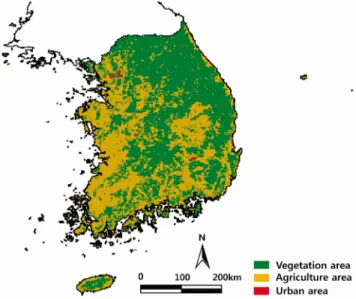
Figure 3. Spatial distribution of land use. Three types of land use: Vegetation, agriculture and urban area were extracted from the digital vegetation map generated by the Global
Land Cover 2000 Project(GLC 2000). Figure 4. Methods to find the optimal threshold point 3. 토지이용 자료
기후조건은 식물종의 생육을 규정하는 주요 요인이며, 식 물종의 생육지는 인간 활동에 의한 토지 이용에 영향을 받 는다. 토지 이용을 고려한 현실적인 식물의 잠재 생육지 (Potential habitats)를 분석하기 위해 전세계 토지 피복 및 모니터링을 위해 기획된 Global Land Cover 2000 Project (GLC 2000)에서 제공하는 DIVA-GIS(www.diva-gis.org) 로부터 30″격자의 약 1km2단위로 생성된 한국의 토지 피 복 자료를 이용하였다(Figure 3).
4. 통계해석 및 평가
한반도 난온대 상록활엽수 57종의 분포에 대한 위치 정 보 자료와 기후변수(WI, TMC, PRS, PRS)와의 관계를 최 대한 균등하게 반복하여 자료(Data)가 무의미 할 때 까지 2분할되도록 구축되는 CT-Model(Classification tree model) (Clark and Pregibon, 1992)을 이용하였다. 본 연구에서는 현재기후(1961~1990) 자료를 적용하여 구축된 한반도 난 온대 상록활엽수 분포 예측 모델(EG-model)(Yun et al., 2011)에 미래기후(2081~2100) 시나리오 CCCMA-A2, CSIRO- A2, HADCM3-A2의 기후자료에서 4가지 기후변수를 적용 하여 잠재 생육지를 분석하였다.
한반도 난온대 상록활엽수의 분포 예측 모델(EG-model) 에서 생육 적지(Suitable habitat)는 식물종의 생육에 적합한
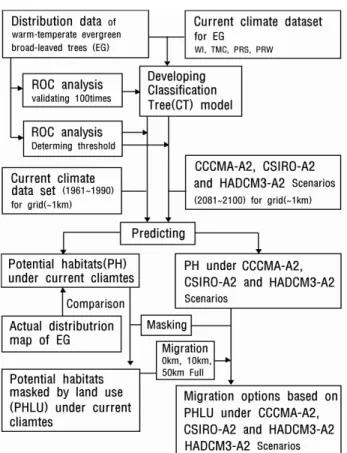
환경을 갖는 지역으로 모델의 적합도가 최고점일 때 분포확 률의 역치 이상인 지역으로 판단하였다(Tsuyama et al., 2008a; 2008b). 생육 적지의 역치는 모델의 예측정도 평가 를 위한 ROC(Receiver operating characteristic)의 해석 (Zweig and Campbelll, 1993)에서 감도(Sensitivity)와 특이 도(Specificity)가 서로 1.0에 가까운 이상점(理想点)과 1.0 과의 거리이며, 계산식은 MinROC dis.= √[(1-Se)² + (1-Sp)²] 이다(Figure 4). 잠재 생육지(PH: Potential habitat) 는 95%의 감도(Sensitivity)일 경우 정의되어지는 출현확률 을 역치로 적용하였다(Tsuyama et al., 2011). 또한, 생육 적지의 역치(閾値)보다는 낮고 잠재 생육지의 분포확률 보 다는 높은 지역으로 식물종의 생육에 그다지 적합하지 않은 환경을 갖는 지역을 분포 변록지(MH: Marginal habitat)로 판단하여 분석하였다. 현실적 분포에 가까운 분포지를 예측 하기 위하여 한반도 토지 이용 자료를 적용하여 현재기후에 서 잠재 생육지(PH: Potential habitats)로 예측된 지역에서 식물종 생육이 불가능한 경작지, 시가지 등의 지역을 제거 하여 토지 이용을 고려한 잠재 생육지(PHLU: Potential habitats masked by land use)를 예측하였다. 또한, 향후 100 년간 현재기후에서의 생육지가 0km, 10km, 50km, Fullkm 확대한다고 가정하여 미래기후에서 토지이용의 영향 및 이 동거리를 고려한 PHLU를 예측하였다. Figure 5는 본 연구 에서 적용한 기후변화에 따른 한반도 난온대 상록활엽수의 잠재 생육지에 대한 분석 흐름도이다. 모델 해석 프로그램 으로는 R2.11.1(R Development Core Team, 2011)을 사용 하였다.
Figure 5. Flowchart of analysis.
(EG: Warm-temperature evergreen broad-leaved trees, PH: Potential habitats, PHLU: Potential habitats masked by land use)
Figure 6. Predicted distribution of warm-temperate evergreen broad-leaved trees under the current climate conditions based on CT- model (EG-model). (A) Potential habitats.
(B) Potential habitats masked by land use. The optimal threshold probability of occurrence was 0.405 from the ROC analysis, which was used to divide the potential habitats into suitable (red area) and marginal habitats (green area).
결 과
1. 현재기후에 대한 잠재적 생육지 예측
Yun et al.(2011)은 한반도 난온대 상록활엽수의 57종에 대하여 현재기후를 적용하여 난온대 상록활엽수 분포 예측 모델(EG-model)을 구축하였다. EG-model은 ROC(Receiver operating characteristic)해석에 의한 모델 예측정도(豫測精 度)를 나타내는 AUC값(Area under the curve)(Hanley and McNeil, 1982; Zweig and Cambell, 1993)이 0.983으로 AUC의 평가기준(Swets, 1988; Thuiller, 2003)에 따라 모 델 구축이 우수(Excellent; 0.9〜1)한 것으로 판단하였으며, TMC(최한월최저기온)가 분포를 규정하는 주요 요인으로 작용하고 있음을 밝힌 바 있다.
난온대 상록활엽수 분포 예측 모델 (EG-model)에 대한 현재기후에서의 4가지(온량지수, 최한월최저기온, 하계강 수량, 동계강수량) 기후변수를 적용한 생육 적지는 모델의 적합도가 최고점일 경우 분포확률을 역치로 한 값 이상의 지역이다. EG-model의 예측정도를 평가하기 위한 ROC분
석에서 ROC 곡선은 X축의 1-특이도(1-Specificity), Y축의 감도(Sensitivity)로 작성된다. 곡선하부 면적이 최대일 때, 즉, 감도(Sensitivity)와 특이도(Specificity)가 서로 1.0에 가 까운 이상점(理想点)과 1.0과의 거리(MinROC dis.)가 역 치이며 0.485로 계산되었다(Figure 4). 잠재 생육지(PH:
Potential habitats)는 분포 재지점(在地点)의 95%를 포함하 는 최대 출현확률로서 95%의 감도(Sensitivity)일 경우 정 의되어지는 출현확률(Tsuyama et al., 2011) 0.3을 역치로 판단하였으며, 역치 이상의 지역을 잠재 생육지로 판단하였 다. 또한, 생육 적지의 역치(閾値)보다는 낮고 잠재 생육지 의 역치보다는 높은 지역으로 역치 0.3~0.485의 지역을 분 포 변록지(MH: Marginal habitats)로 판단하여 현재기후에 서 EG-model에 의한 한반도 상록활엽수의 생육지를 예측 하였다(Figure 6).
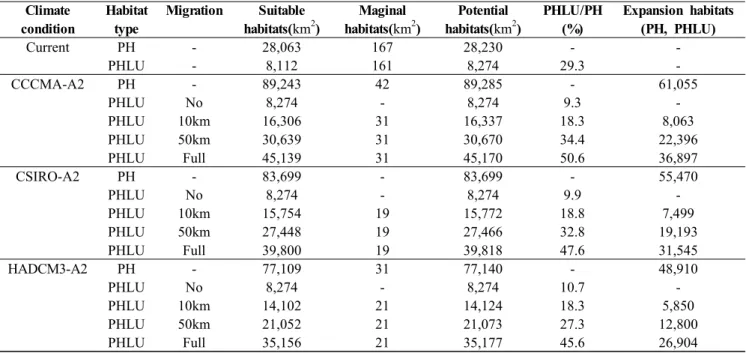
한반도(남한) 전역을 지도화 할 경우 30″단위의 총 격자 수 145,012개이며, 약 99,898km2의 면적이다. 난온대 상록 활엽수 분포 예측 모델(EG-model)의 결과, 현재기후에서 생육 적지와 분포 변록지의 면적은 각각 28,063km2, 167km2이며, 잠재 생육지(PH)는 28,230km2로 예측되었다 (Figure 6, Table 2). 잠재 생육지의 분포 지역으로는 서해안
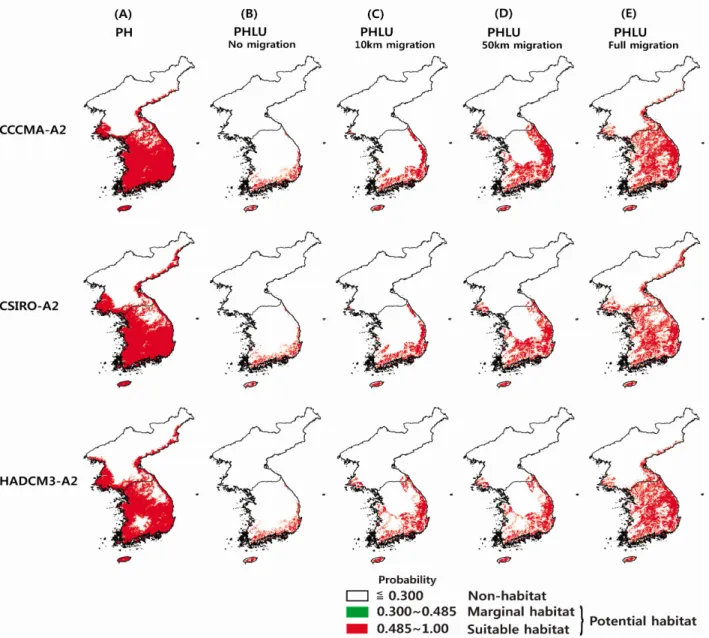
Figure 7. Predicted distribution of warm-temperate evergreen broad-leaved trees under the climate change scenarios (CCCMA-A2, CSIRO-A2 and HADCM3-A2). (A) Potentioal habitat. (B) No migration options based on PHLU. (C) 5km migration options based on PHLU. (D) 50km migration options based on PHLU. (E) Full migration options based on PHLU. A spatial filter was built on GIS, which allows range expansions of the species only within the neighboring grid.
백령도, 충청남도 태안, 전라북도 전주, 전라남도 담양, 경상 남도 하동, 경상북도 대구, 경상북도 울진, 강원도 속초를 잇는 하한을 생육지로 하고 있으며, 경상남도 가지산 일대는 제외되었다. 또한, 현재기후에서 예측된 잠재 생육지에서 토 지 이용에 의하여 식물종의 생육이 불가능한 경작지, 시가지 등을 제거하여 예측된 토지 이용을 고려한 잠재 생육지 (PHLU)는 8,274km2이며, 잠재 생육지(PH)의 29.3%를 차 지하며, 남한 면적의 약 8.3%를 차지하는 면적으로 예측되 었다.
2. 미래기후에 대한 잠재적 생육지 예측
EG-mode을 토대로 2081~2100년의 미래기후에 대한 난 대성 상록활엽수의 분포를 예측한 결과, 잠재 생육지(PH) 는 CCCMA-A2에서 89,285km2 분포역이 예측되었으며, CSIRO-A와 HADCM3-A2에서 각각 83,699km2, 77,140km2 분포역을 차지할 것으로 예측 되었다(Figure7-A). 3가지 미 래기후 시나리오에 대하여 난대성 상록활엽수는 남한 면적 의 77~89% 해당한다. 또한, CCCMA-A2 시나리오에서의
Climate Habitat Migration Suitable Maginal Potential PHLU/PH Expansion habitats condition type habitats(km2) habitats(km2) habitats(km2) (%) (PH, PHLU)
Current PH - 28,063 167 28,230 - -
PHLU - 8,112 161 8,274 29.3 -
CCCMA-A2 PH - 89,243 42 89,285 - 61,055
PHLU No 8,274 - 8,274 9.3 -
PHLU 10km 16,306 31 16,337 18.3 8,063
PHLU 50km 30,639 31 30,670 34.4 22,396
PHLU Full 45,139 31 45,170 50.6 36,897
CSIRO-A2 PH - 83,699 - 83,699 - 55,470
PHLU No 8,274 - 8,274 9.9 -
PHLU 10km 15,754 19 15,772 18.8 7,499
PHLU 50km 27,448 19 27,466 32.8 19,193
PHLU Full 39,800 19 39,818 47.6 31,545
HADCM3-A2 PH - 77,109 31 77,140 - 48,910
PHLU No 8,274 - 8,274 10.7 -
PHLU 10km 14,102 21 14,124 18.3 5,850
PHLU 50km 21,052 21 21,073 27.3 12,800
PHLU Full 35,156 21 35,177 45.6 26,904
Table 2. Comparison between the area of PH (PHLU) under the current climate and that under the climate change scenarios
잠재 생육지(PH)는 현재기후에서 보다 61,055km2 증가하며, CSIRO-A2와 HADCM3에서 각각 55,470km2, 48,910km2 분포역이 증가하는 것으로 나타났다(Table 2).
현재기후에서 식물종의 생육지는 토지 피복에 의해 축소 및 산재되어 생육하고 있으며, 기후변화에 따른 미래기후에 서 난온대 상록활엽수 생육지는 현재 생육지를 중심으로 식물종의 이동거리에 따라 분포역이 확대 된다(Nakao et al., 2011). 따라서, 본 연구에서 난온대 상록활엽수의 향후 100년간 이동거리를 0km(이동하지 않음), 10km, 50km, 완 전(Full)이동 할 것으로 가정하여 미래기후에서 토지이용을 고려한 잠재 생육지(PHLU)를 예측하였다. 미래기후에서 전 혀 이동하지 않을 것(0km, No migration)이라고 가정 할 경 우, 토지이용을 고려한 잠재 생육지(PHLU)는 CCCMA-A2, CSIRO-A2, HADCM3-A2에서 8,274km2의 분포역이 예측 되었으며, 잠재 생육지(PH)의 약 9~10%에 해당한다 (Figure 7-B, Table 2). 10km 이동 할 경우, CCCMA-A2에 서 PHLU는 16,337km2, CSIRO-A2와 HADCM3-A2에서 각각 15,772km2, 14,124km2의 분포역이 예측되었으며, PH 의 약 18.5%해당한다(Figure 7-C, Table 2). 50km 이동 할 경우, CCCMA-A2에서 PHLU는 30,670km2, PH의 34.4%
에 해당하며, CSIRO-A2와 HADCM3-A2에서 PHLU는 각 각 27,466km2, 21,073km2의 분포역이 예측되며, PH의 32.8%, 27.3%에 해당한다(Figure 7-D, Table 2). 완전 이동 (Full migration)의 경우, 미래기후에서 예측된 잠재생육지 (PH)에서 식물종 생육의 불가능한 경작지, 시가지 등의 지
역을 제거한 생육지와 동일하며, CCCMA-A2에서 PHLU 는 45,170km2, PH의 50.6%에 해당한다. CSIRO-A2와 HADCM3-A2에서 PHLU는 각각 39,818km2, 35,177km2 의 분포역이 예측되며, PH의 47.6%, 45.6%에 해당한다 (Figure 7-E, Table 2).
고 찰
Yun et al.(2001)은 EG-model을 통해 예측된 난온대 상 록활엽수의 분포역을 결정하는 기후요인으로 최한월최저 기온(TMC) -.5.95℃가 제 1요인으로 작용하고 있으며, 최 한월평균기온 -1.7℃에 대응하고 있는 것으로 분석하였다.
또한, 온도요인 이외의 강수요인으로 동계 강수량보다 하계 강수량이 난온대 상록활엽수의 분포에 영향 요인으로 작용 하고 있음을 분석하였다(Yun et al., 2011).
현재기후에서 잠재 생육지(PH)는 서해안 도서지역을 비 롯하여 남부 도서지역과 남부 내륙지역, 동해안 인접지역의 낮은 구릉지를 대상으로 분포역이 예측되었으며, 분포면적 28,230km2로 남한면적의 28%를 차치하였다. 생육적지는 28,063km2로 한반도에서 예측되었으며, 분포 변록지는 167km2로 제주도와 울릉도 지역에서 예측되었다.
기후변화에 의한 미래기후에서의 잠재 생육지(PH)는 3종 류 기후 시나리오 CCCMA-A2, CSIRO-A2, HADCM3-A2 에서 면적 77,140~89,285km2로 예측되어 한반도 면적의 77.2~89.4%를 차지하였다. CCCMA-A2, CSIRO-A2는 강
원도의 일부 지역을 제외한 지역에서 잠재 생육지가 예측되 었으며, HADCM-A2에서는 설악산, 오대산, 태백산의 서부 내륙지역의 강원도 지역, 덕유산과 지리산일대의 전라북도 와 경상남도 일부 지역을 제외한 지역에서 잠재 생육지가 예측되었다. 북한지역에서는 동해의 인접지역과 서해의 인 접지역 및 황해도, 평안남도의 내륙지역에 잠재 생육지가 예측되었다.
기후변화에 따른 온난화 영향 예측은 현재 및 미래의 식 물종의 생육 가능 지역을 평가할 때 중요한 자료로 활용할 수 있다. 기후변화에 따른 생육지의 변화는 기후조건과 함 께 토지이용에 따른 식물 생육지의 분단화(分斷化), 고립화 (孤立化) 및 식물종의 이동 속도 등의 복합적 요인이 작용 함으로 기후조건 만에 의한 생육지 변화와 비교하면 크게 제한되어 분포한다. 따라서, 토지이용 및 분포 이동을 고려 한 예측은 기후적으로 적합하지만 토지이용이 진행되는 지 역을 현재의 생육지에서 제거함으로써 기후조건만으로 예 측된 과대평가된 지역을 보정하여 현실에 근접한 예측을 할 수 있다.
현재기후에서 토지이용을 고려한 잠재 생육지(PHLU)의 면적은 8,274km2로 남한면적의 8.3%를 차지하며, 잠재 생 육지(PH) 면적의 29.3%를 차지하였다. 미래기후에서 토지 이용을 고려한 잠재 생육지는 35,177~45,170km2로 예측되 었으며, 약 26.9~36.9%로 증가하였다. 또한, 토지 이용으로 난온대 상록활엽수의 잠재생육지는 분단화 및 고립화로 인 해 산재적으로 분포한다. 이러한 임분(林分) 면적의 축소, 분단화, 고립화는 상록활엽수림의 종다양성을 저하시키는 주요요인으로 작용 될 수 있다(Hattori et al., 2002). 또한, 난온대 상록활엽수의 이동속도 및 거리는 분포확대에 중요 한 요소이다. 기후변화에 따른 난온대 상록활엽수림의 분포 확대는 졸참나무림, 신갈나무림 등의 낙엽활엽수림과의 경 쟁이 예상되며, 상록활엽수와 낙엽활엽수가 국소적으로 분 포하는 이행대(移行帶)의 지역에서는 상록활엽수의 그늘로 인한 낙엽활엽수의 실생(實生)과 치수(稚樹)의 생육에 악 영향을 줄 것으로 판단된다(Tagawa, 1973). 또한, 상록활엽 수의 생육 증가는 낙엽활엽수의 생육에 제한요인으로 빛의 수광량(受光量)의 변화로 인한 낙엽활엽수림의 쇠퇴에 영 향을 줄 수 있다. 본 연구의 결과에 더불어 정확한 생육지 예측을 위해서는 식물종의 종자산포 방식과 정착지의 종간 상호작용, 인접지역과의 관계 등의 영향, 미래 인구 증가에 따른 인간 간섭 등을 고려한 예측이 필요할 것으로 판단된 다. 또한, 일본과 중국에 분포하는 난온대 상록활엽수의 분 포자료를 포함하여 분석함으로써 한반도 지역에서 과소평 가된 잠재 생육지 예측을 보정할 수 있다고 판단된다.
본 연구에서 기후변화에 따른 한반도 난온대 상록활엽수 의 온난화 영향 예측을 기후조건, 토지이용, 현재의 생육지
에서 인접거리를 고려한 잠재 생육지를 분석하였다. 향후 난온대 상록활엽수 중에서 교목층을 구성하는 상록활엽수 종 및 군락의 분포적 기후조건과 잠재 분포지에 대한 예측 연구는 기후변화에 따른 난온대 상록활엽수림의 식생대 변 화를 구체적으로 분석할 수 있을 것이며, 동시에 지속적인 분포지의 모니터링을 바탕으로 기후변화에 따른 식물의 보 전 전략수립에 중요한 자료로 활용될 것으로 생각된다.
인용문헌
Clark, L.A. and D. Pregibon(1992) Tree-based models. In: J. M.
Chambers and T. J. Hastie, eds., Statistical Models in S, California, Wadsworth & Brooks/Cole Advanced Books &
Software, Pacific Grove, pp. 377-419.
De’Ath, G., and K. E. Fabricius(2000) Classification and regression trees: A powerful yet simple technique for ecological data analysis. Ecology 81(11): 3178-3192.
Bakkenes, M., J.R.M. Alkenmade, F. Ihle, R. Leemans and J.B.
Latour(2002) Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Global Change Biology 8: 390-407.
Berry, P.M., T.P. Dawson, P.A. Harrison, R. Pearson and N.
Butt(2003) The sensitivity and vulnerability of terrestrial hab- itats and species in Britain and Ireland to climate change.
Journal of Nature Conservation 11: 15-23.
Flora of Korea Editorial Committee(2007) The genera of Vascular Plants of Korea. Academy Publishing co., Seoul, 1,482pp.
Hanley, J. and B. McNeil(1982) The meaning and use of the area un- der a receiver operating characteristic (ROC) curve. Radiology 143: 29-36.
Harrison, P.A., P.M. Berry and T.P. Dawson, eds.(2001) Climate Change and Nature Conservation in Britain and Ireland:
Modelling Natural Resource Responses to Climate Change(the MONARCH project), UKCIP Technical Report, Oxford.
Hattori T., S.H. Ishida, S. Kodane, and N. Minamiyama(2002) The characteristics of lucidophyllous forest flora and conditions en- dangering them in Japan. J JILA 65(5): 609-614. (in Japanese with English summary)
Huntley, B., P.M. Berry, W. Cramer and A.P. McDonald(1995) Modelling present and potential future ranges of some European higher plants using climate response surfaces.
Journal of Biogeography 22: 967-1001.
IPCC(2007) Climate Change 2007: the Physical Science Basis, Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change.
Cambridge University Press.
Iverson, L.R. and A.M. Prasad(1998) Predicting abundance of 80 tree species following climate change in the eastern United
States. Ecological Monographs 68: 465-485.
Iverson, L.R., A.M. Prasad, B.J. Hale and E.K. Sutherland(1999) An Atlas of Current and Potential Future Distributions of Common Trees of the Eastern United States, Gen. Tech. Rep.
NE-265, US. Department of Agriculture, Forest Service, Northeastern Research Station.
Kira, T.(1977) A Climatological interpretation of Japanese vegeta- tion zones. In Miyawaki, A. and Tuxen, R. (eds.) Vegetation science and environmental protection. Maruzen, Toktyo, pp.
21-30.
Leathwick, J.R., D. Whitehead and M. McLeod(1996) Predicting changes in the composition of New Zealand's indigenous for- ests in response to global warming: a modeling approach.
Environmental Software 11: 81-90.
Matsui, T., T. Yagihashi, T. Nakaya, H. Taoda and N. Tanaka (2004a) Climatic controls on distribution of Fagus crenata for- ests in Japan. Journal of Vegetation Science 15: 57-66.
Matsui, T., T. Yagihashi, T. Nakaya, H. Taoda S. Yoshinaga, H.
Daimaru and N. Tanaka(2004b) Probability distributions, vul- nerability and sensitivity in Fagus crenata forests following predicted climate changes in Japan. Journal of Vegetation Science 15: 605-614.
Nakao, K., T. Matsui, M. Horikawa, I. Tsuyama and N. Nobuyuki (2011) Assessing the impact of land use and climate change on the evergreen broad-leaved species of Quercus acuta in Japan.
Plant Ecol. 221: 229~243.
R Development Core Team(2011) R: A language and environment for statistical computing. R. Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL://www.R-project.org.
Swets, K.A.(1988) Measuring the accuracy of diagnostic systems.
Science 240: 1285-1293.
Sykes, M.T., I.C. Prentice and W. Cramer(1996) A bioclimatic model for the potential distributions of European tree species under present and future climates. J. Biogeography 23:
203-233.
Tagawa, H.(1973) Seitai seni. Kyoritu Shuppan. Tokyo, pp. 87-92.
Tanaka, N., E. Nakazono, I. Tsuyama and T. Matsui(2009) Assessing impact of climate warming on potential habitats of ten conifer species in Japan. Global Environmental Research 14: 153-164.
Thuiller, W.(2003) BIOMOD-optimizing predictions of species distributions and projecting potential shifts under global change. Global Change Biology 9: 1353-1362.
Thuiller, W., S. Lavorel, M.B. Araújo, M.T. Sykes and I.C.
Prentice(2005) Climate Change threats to plant diversity in Europe. Proceeding of the National Academy of Sciences of the United States of America 102: 8245-8250.
Tsunekawa, A., H, Ikeguchi and K. Omasa(1993) Effects of cli- matic change on the vegetation distribution in response to cli- matic change. In: Omsa, K., K. Kai, H. Taoda, Z. Uchijima and M. Yoshino eds., Climate change and Plant in East Asia, Springer, 57-65.
Tsuyama, I., T. Matsui, M. Ogawa, Y. Kominami and N.
Tanaka(2008) Habitat prediction and impact assessment of cli- mate change on Sasa kurilensis in eastern Honshu, Japan.
Theory and Applications of GIS 16(1): 11-25.
Tsuyama, I., K. Nakao, T. Matsui, M. Higa, M. Horikawa, Y.
Kominami and N. Tanaka(2011) Climatic controls of a key- stone understory species, Sasamorpha borealis, and an impact assessment of climate change in Japan. Annals of Forest Science: dio: 10.1007/s 13595-011-0086-y.
Uchijima, Z., H. Seino and M. Nogami(1992) Probable shifts of nat- ural vegetation in Japan due to CO2-climate warming. In: M.
Shiomi, E. Yano, H. Koizumi, D. A. Andow and N. Hokyo, eds., Ecological Process in Agro-ecosystems, Yokendo: 189-201.
Yun, J.H., J.H. Kim, K.H, Oh and B.Y. Lee(2011) Distributional change and climate conditions of warm-temperate evergreen broad-leaved trees in Korea. Kor. J. Env. Eco. 25: 47-56. (in Korean with English summary)
Zweig, M.H. and G. Campbell(1993) Receiver-operating charac- teristic (ROC) Plots: a fundamental evaluation tool in clinical medicine. Climical Chemistry 39: 561-577.