기후변화에 따른 한반도 난대성 상록활엽수 잠재서식지 분포 변화
박선욱*·구경아**·공우석***
Potential Impact of Climate Change on Distribution of Warm Temperate Evergreen Broad-leaved Trees in the Korean Peninsula
Seon Uk Park* · Kyung Ah Koo** · Woo-Seok Kong***
요약 :본 연구는 난대성 상록활엽수에 대한 기후변화의 영향을 평가하고자 하였다. 이를 위해 분포 북방한계 지가 중부해안인 난대성 상록활엽수 9종을 선정하고, 각 종의 분포 자료와 최한월최저기온, 겨울철강수량에 Generalized Additive Model(GAM) 알고리즘을 적용하여 종분포모형을 구축하였다. 종분포모형에 현재와 미 래기후자료, 토지이용도를 적용하여 난대성 상록활엽수의 현재와 미래 잠재서식지를 예측하였다. 기후요소 분 석 결과에서 최한월최저기온은 모든 종의 분포에 유의한 영향을 주지만, 겨울철강수량은 종에 따라 영향이 다 르게 나타났다. 9종은 잠재서식지의 분포 패턴에 따라 3개의 유형(중부해안형, 남서해안형, 중부경상내륙형)으 로 분류되며 기후변화와 토지이용의 영향이 다르게 나타났다. 토지이용을 고려했을 때 9종에서 60% 이상의 잠 재서식지가 감소하며, 특히 중부경상내륙형에 속하는 종들은 다른 유형에 비해 높은 서식지 감소를 보였다. 9 종 모두 기후변화에 따라 2050년과 2070년에 분포지가 북쪽으로 확대될 것으로 예측되었지만, 분포 유형에 따 라 각기 다르게 나타났다. 이러한 분포 유형별 기후변화 영향이 다른 것은 겨울철강수량의 시공간적 변화와 종 별로 미치는 영향의 정도가 다르기 때문으로 판단된다.
주요어 : 난대성 상록활엽수, 종분포모형, GAM, 기후변화, 토지이용, 잠재서식지
Abstract : We accessed the climate change effects on the distributions of warm-evergreen broad-leaved trees (shorten to warm-evergreens below) in the Korean Peninsula (KP). For this, we first selected nine warm- evergreens with the northern distribution limits at mid-coastal areas of KP and climate variables, coldest month mean temperature and coldest quarter precipitation, known to be important for warm-evergreens growth and survival. Next, species distribution models (SDMs) were constructed with generalized additive model (GAM) algorithm for each warm-evergreen. SDMs projected the potential geographical distributions of warm evergreens under current and future climate conditions in associations with land uses. The nine species were categorized into three groups (mid-coastal, southwest-coastal, and southeast-inland) based on their current spatial patterns. The effects of climate change and land uses on the distributions depend on the current spatial patterns. As considering land uses, the potential current habitats of all warm-evergreens decrease over 60%, showing the highest reduction rate for the Kyungsang-inland group. SDMs forecasted the expansion of potential habitats for all warm-evergreens under climate changes projected for 2050 and
* 국립생태원 생태연구본부 융합연구실 전문위원 (Research Fellow, Division of Ecosystem Services & Research Planning, Bureau of Ecological Research, National Institute of Ecology), [email protected]
** 국립생태원 생태연구본부 융합연구실 전문위원 (Research Fellow, Division of Ecosystem Services & Research Planning, Bu- reau of Ecological Research, National Institute of Ecology), [email protected]
*** 경희대학교 이과대학 지리학과 교수(Professor, Department of Geography, College of Sciences, Kyung Hee University), ws- [email protected]
박선욱·구경아·공우석
1. 서론
20세기 산업화와 도시화로 지난 100년간 전지 구 평균온도는 0.78℃ 상승하였으며, 향후 100년간 1.0~3.7℃ 증가할 것으로 예측된다(Stocker et al., 2013). 특히, 우리나라가 위치한 동아시아는 복잡 한 지역적 특성으로 인해 기후변동성이 크며, 21세 기 말에 기온이 1.8~4.9℃까지 증가할 것으로 추정 되었다(국립기상연구소 2012). 기온과 더불어 해수면 과 강수량에서도 변화가 관측되었으며, 앞으로 이러 한 변화는 가속화될 것으로 예측되었다. 해수면은 과 거 112년(1901~2011년)간 0.17~0.21m 높아졌고, 최 근 들어 해수면 상승 추세는 빨라지고 있다(Stocker et al., 2013). 기후변화에 따른 우기와 건기 및 건조지역 과 습윤지역의 강수량 격차가 현재보다 증가하고, 강 수량의 시·공간적 변동성이 더욱 커질 것으로 예측 되었다(Stocker et al., 2013).
미래 기후변화에 대한 전망이 보고되면서, 다양한 생물들의 서식지 변화에 대한 기후변화 영향을 평가 하고, 그에 따른 대응 대책을 마련하고자 하는 연구 들이 진행되었다(공우석, 1999; 2005; 구경아, 2000;
서민환 등, 2006; 임종환·신준환, 2005; Colwell et al., 2008; Feeley et al., 2011; Feeley, 2012). 많은 연 구들이 기후변화에 따라 생물계절의 변화, 난대성 식 물들의 북상 및 서식지 확대, 한대성 식물들의 서식 지 유실 및 멸종가능성 등을 예측하였다(Díaz-Varela et al., 2010; Dullinger et al., 2012; Koo et al., 2015).
기후변화에 따라 고산지역과 극지역에 서식하는 식 물들의 봄철 식물계절현상들이 빨라졌으며, 유럽의 산지들에 분포하는 난대성 식물들의 서식지가 고도 가 높은 지역으로 확대되었다(Zimmermann et al.,
2013). 이와는 대조적으로, 한랭한 기후에 적응한 유 럽의 고산식물들은 분포지가 쇠퇴하고 있는 것으로 분석되었다(Randin et al., 2009).
종분포모형(Species Distribution model, SDM)은 종의 출현/비출현과 환경변수와의 관계를 통해 종의 출현 가능성을 분석하는 모형으로, 생물종들의 서식 지 환경에 대한 기후변화를 포함한 미래 환경변화의 영향을 평가하는데 광범위하게 사용되어 왔다(Aus- tin, 2002). 종분포모형은 통계기법들을 이용한 상관 모형(Correlative Model)(Guisan and Zimmermann, 2000)부터 생태적인 과정을 기반으로 한 과정기반모 형(Process-based model)(Kleidon and Mooney, 2000;
Morin et al., 2007), 그리고 종분포와 환경변수들 사 이의 역학적 작용들을 바탕으로 종의 분포를 예측하 는 역학모형(Mechanistic Model)(Kearney and Porter, 2009)으로 나누어진다. 가장 광범위하게 사용되는 종분포모형은 통계기법들을 기반으로 한 모형으로 (Guisan and Zimmermann, 2000), 종의 출현/비출현 과 환경변수들의 상관관계를 바탕으로 종의 출현율 을 예측한다. 이 종분포모형에는 Generalized linear model(GLM), Generalized additive model(GAM), Multivariate adaptive regression splines(MARS), Classification tree analysis(CTA) 등 다양한 통계기법 이 활용되고 있다(서창완 등, 2008; 권혁수 등, 2012;
2014; 김지연 등, 2012; 박현철 등, 2015; 신만석 등, 2015; 이용호 등, 2015; Thuiller, 2003).
우리나라 난대성 상록활엽수는 한반도 남부 및 해 안도서에서 나타나는 식물로써 겨울철기온에 따라 북 방한계선이 결정되는 것으로 알려져 있으며(Uyeki, 1941), 기온상승에 따라 분포지가 확대될 것으로 예 측되었다(박종철 등, 2010). 식물표본자료을 바탕으 로 복원된 일제시대 난대성 상록활엽수 분포를 현재 2070. However, the expansion patterns were different among three groups. The spatial patterns of projected coldest quarter precipitation in 2050 and 2070 could account for such differences.
Key Words : warm temperate evergreen broad-leaved tree, species distribution model(SDM), generalized additive model (GAM), Climate change, land use, potential habitat
의 분포와 비교한 결과 분포한계선이 북상한 것으로 나타났다(윤종학 등, 2011). 또한, 붉가시나무, 구실 잣밤나무, 후박나무, 참식나무 등 4개 종을 대상으로 종분포모형을 구축하고 미래 분포지를 예측한 결과, 모든 종의 서식지가 북쪽으로 확대되는 것으로 나타 났다(Yun et al., 2014). 특히, 후박나무의 잠재서식지 는 남한 전역으로 확대될 것으로 예측되었다(Yun et al., 2014). 한국과 동일한 상록활엽수종들을 공유하 는 일본과 대만에서도 기후변화에 따라 난대성 상록 활엽수의 분포가 북쪽으로 확대될 것으로 보았다. 일 본의 붉가시나무를 대상으로 종분포모형을 구축하고 미래 분포를 예측한 결과, 분포지가 북상하는 것으로 예측되었다(Nakao et al., 2011). 일본과 대만에 서식 하는 난대성 상록활엽수 10종을 대상으로 미래 잠재 서식지의 변화를 예측한 결과 분포가 확대되는 것으 로 예측되었다(Nakao et al., 2011; 2014). 미국 플로 리다에 자생하는 100여개의 고유종의 분포자료와 8 개의 기후변수(최난월 평균기온, 최한월평균기온, 월 평균 연교차, 최한월 평균최저기온, 절대최저기온, 연평균강수량, 건조한 달의 연 평균 강수량, 연수분 지수)를 통해 Climatic-envelope model을 구축하여 기후변화의 변화에 따른 분포범위 변화를 예측한 결 과, 아열대 수종의 분포범위가 기존보다 북상하는 것 으로 분석되었다(Box et al., 1993; Crumpacker et al., 2001).
본 연구에서는 남부와 해안도서에 주로 분포하 며 분포 북한계지가 충남 태안반도보다 북쪽에 위 치한 난대성 상록활엽수 9종(보리밥나무(Elaeagnus macrophylla Thunb.), 보리장나무(Elaeagnus glabra Thunb.), 마삭줄(Trachelospermum asiaticum(Siebold
& Zucc.) Nakai), 줄사철나무(Euonymus fortunei var.
radicans (Miq.) Rehder), 개산초(Zanthoxylum planis- pinum Siebold & Zucc.), 동백나무(Camellia japonica L.), 후박나무(Machilus thunbergii Siebold & Zucc.), 센달나무(Machilus japonica Siebold & Zucc.), 붉가시 나무(Quercus acuta Thunb.))을 대상으로 종분포모형 을 구축하고, 이를 토대로 현재의 종별 잠재서식지와 미래 기후변화에 따른 잠재서식지의 변화추세를 예 측하였다. 본 연구는 특히 잠재서식지를 고려함에 있
어 토지이용을 고려하여 현재 기후조건 하에서 잠재 서식지의 예측에 현실성을 높였으며, 인간의 간섭이 난대성 상록활엽수의 분포에 미치는 영향을 설명하 였다.
2. 연구방법 및 자료
1) 연구종 및 연구지역
연구종은 남부와 해안도서에 주로 분포하며 분포 북한계지가 충청남도 태안반도 이북에 분포하고 있 는 9종의 난대성 상록활엽수인 보리밥나무, 보리장 나무, 마삭줄, 줄사철나무, 개산초, 동백나무, 후박나 무, 센달나무, 붉가시나무이다. 이 난대성 상록활엽 수들은 식물의 외관형에 따라 교목 4종(동백나무, 후 박나무, 센달나무, 붉가시나무), 관목 1종(개산초), 덩굴식물 4종(보리밥나무, 보리장나무, 마삭줄, 줄사 철나무)으로 분류되었다. 이 수종들 중에 후박나무와 동백나무는 기후변화가 한반도 생물종에 미치는 영 향을 감시 및 예측하기 위한 기후변화 생물지표 100 종으로 선정되었다(환경부, 2000).
연구지역은 북위 33°~38° 동경126°~133°에 위치 한 한반도와 그 부속도서로 선정하였다. 한반도는 남 북으로 길게 나타난 반도로써 전체 면적 220,847㎢의 70%가 산지로 이루어졌으며 부속도서는 3,400여개 이다. 서해와 남해안은 리아스식 해안으로 해안선이 복잡하며 조수간만의 차가 크지만 동해안은 단순하 며 조수간만의 차가 크지 않다(국토교통부 국토지리 정보원, 2014). 또한 동고서저 지형으로 서해안은 해 안과 도서지역에 난대성 상록활엽수가 서식할 수 있 는 공간을 제공하지만 이와 반대로 동해안 지역은 상 대적으로 높은 온도(기상청, 2014)에 비해 협소한 공 간으로 동일 위도인 서해안과 비교해서 난대성 상록 활엽수가 나타나지 않는다.
박선욱·구경아·공우석
2) 환경변수들
난대성 상록활엽수의 분포에 영향을 주는 기후요 소들은 최한월최저기온, 겨울철강수량, 여름철평균 기온, 여름철강수량 등이다(Nakaoa et al., 2014). 최 한월최저기온은 난대성 상록활엽수의 겨울철 생존에 영향(Sakai, 1975)을 주며, 여름철평균기온은 여름철 성장과 생존에 영향을 미친다. 두 기온변수들 모두 난 대성 상록활엽수의 분포와 생장을 설명하는데 생리학 적으로 중요한 의미를 가진다. 그러나 두 변수간의 상 관관계가 매우 높아(r>0.75), 본 연구에서는 난대성 상 록활엽수의 분포한계 및 생장의 기온에 대한 내성을 설명하는 최한월최저기온을 독립변수로 사용하였다.
겨울철강수량은 겨울철 수분공급에 매우 중요한 지표이며, 일부 지역에서는 적설량으로 측정할 수 있 다. 특히, 적설량은 겨울철 식물의 하단부를 덮어서 보온효과를 주며(Nakao et al., 2014), 겨울철에 쌓 인 눈이 건조한 봄철에 녹으면서 식물의 수분공급을 원활하게 도와준다. 여름철강수량은 난대성 상록활 엽수들이 성장하는 시기에 수분을 공급하여 중요한 의미를 가지나, 최한월최저기온과 상관관계가 높아 (r>0.75), 겨울철강수량만을 종분포모형 개발을 위한 독립변수로 사용하였다.
독립변수로 사용되는 기후자료는 worldclim(http:
//www.worldclim.org)에서 30-arc second(ca. 1㎢) 의 해상도로 제공하는 50년 평균기후(1950~2000년) 의 최한월최저기온과 겨울철강수량을 한반도 범위에 서 추출하여 사용하였다. 미래기후는 미래 온실가스 절감 정책이 많은 부분 실현되는 RCP 4.5와 현재 배 출 추세가 계속되는 것으로 가정하는 RCP 8.5시나리 오에 따라 영국헤들리연구소에서 제공하는 전세계기 후모델(GCM, Global Climate Model)인 HadGEM2- AO를 활용하여 추정된 2050년(2040~2060년)과 2070년(2060~2080년)의 기후자료를 사용하였다 (Hijmans et al., 2005).
종분포모형은 환경변수를 통해 종 출현가능성을 예측하는 모형으로 비환경변수를 추가함으로써 모형 의 현실성을 높일 수 있다(Guisan and Zimmermann 2000). 토지이용도는 자연환경에 대한 인간의 간섭
을 확인할 수 있는 비기후적 요소로써 잠재서식지에 서 토지이용현황을 고려함으로 인간의 간섭이 포함 된 실재서식지를 파악하는 데 유용하다. 본 연구에서 는 Global Land Cover 2000 Project(GLC2000)에서 위성사진을 통해서 분류한 토지피복도를 DIVA-GIS (http://diva-gis.org)에서 다운받아 한반도 범위에서 산림, 농경지, 시가지로 재분류하여 산림지역과 잠 재서식지가 중복되는 영역을 토지이용을 고려한 잠 재서식지로 분류하여 면적을 산출하였다. 산출된 토 지이용을 고려한 잠재서식지 면적과 잠재서식지와의 비율을 통해 인간에 의한 영향을 종별로 분석하였다.
3) 모형개발과 정확도 평가
종분포모형의 개발에 필요한 연구종들에 대한 출 현/비출현자료는 구경아(2000), 이우철과 임양재 (2002), 국립수목원의 관속식물분포도(2004; 2005;
2006; 2007; 2008; 2009; 2010a; 2010b; 2011)에 수 록된 식물 분포자료를 사용하였다. 식물조사 지점이 동일하거나 인접되어 나타나는 지점은 통합하여 종 별 200여개의 식물분포자료를 사용하였다. 식물조사 가 이루어졌지만 연구종들이 나타나지 않은 지점에 대해서는 비출현자료로 활용하였다.
종분포모형은 독립변수와 종속변수간의 비선 형관계를 표현하기 위해 GLM(Generalized Linear Model)에 평활함수(smooth function)가 추가된 GAM(Generalized Additive Model)을 사용하여 개 발하였다(Wood, 2006). GAM 알고리즘에서 각 독 립변수의 평활함수 차수는 교차검증에 의해 자동적 으로 선정되며, 4차 함수 이내로 한정된다(Thuiller, 2003). 가장 적절한 수의 독립변수들을 가지며, 동시 에 모델 정확도가 높은 모형을 선정하기 위하여 독립 변수의 수를 단계적으로 늘려가며 검증하는 방법을 사용한다(Thuiller, 2003). 모델의 정확도는 GLM과 같이 Akaike Information Criterion(AIC) 값을 적용하 여 평가된다.
잠재서식지는 환경적 요소가 종의 출현을 제한하 는 생태적 지위(Ecological niche)에 기반한다. 연구종 의 분포자료와 환경변수간의 상관관계를 분석을 통
해 연구지역의 종 출현 가능성을 추정하며 종이 분포 할 수 있는 지역을 잠재서식지로 분류한다. 연구종에 대한 출현 가능성은 확률로 나타내기 때문에 잠재서 식지를 설정하기 위하여 분포 여부를 구분하는 임계 점을 설정해야 한다. 이를 위해 sensitivity(출현율 예 측 정확도)와 specificity(비출현율 예측 정확도)가 동 일해지는 지점을 임계점으로 설정하였다(Freeman and Moisen, 2008). Sensitivity(omission error)는 모 델에 의해 예측된 종의 출현이 실제의 관찰과 얼마 나 일치하는지를 보여주는 값이며, Specificity는 모 델에 의해 예측된 종의 absence(비출현) 값이 실제 의 관찰된 값과 얼마나 일치하는지는 보여주는 값 이다(Jiménez-Valverde et al., 2007). 모델 예측의 정 확도는 ROC(Receiver operating characteristic)분석 의 AUC(Area under curve) 방법을 사용하여 평가되 었다. 식물분포자료가 많지 않기 때문에 부트스트랩 (bootstrap) 기법을 사용하여 종의 출현/비출현 자료 와 기후자료를 1000번 복원추출하고, 이 1000개의 복원추출 자료에 대하여 ROC분석의 AUC 값을 계산 하여 모델의 정확도를 측정하였다(Zweig and Camp- bell, 1993). 모델의 정확도를 나타내는 AUC 값은 0.5~0.6(매우 낮음), 0.6~0.7(낮음), 0.7~0.8(보통), 0.8~0.9(높음), 0.9~1(매우 높음)으로 평가할 수 있 다(Swets, 1988; Thuiller, 2003).
3. 연구 결과 및 고찰
1) 난대성 상록활엽수의 현재 잠재서식지 예측 및 모델의 정확도
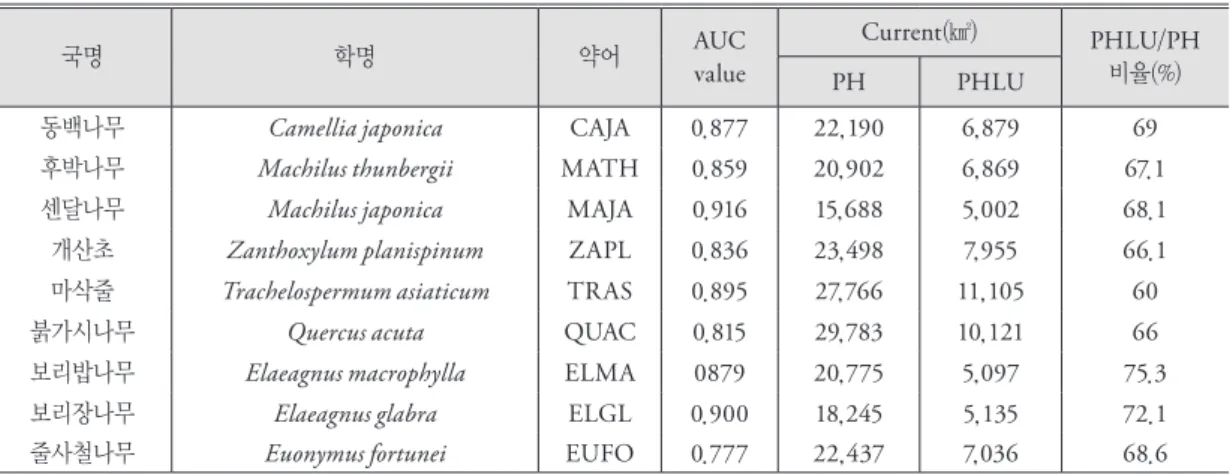
기후요소들과 각 종의 출현/비출현 자료에 GAM 알고리즘을 적용하여 9개의 난대성 상록활엽수종들 에 대한 종분포모형들을 개발하였다. 이 모형들을 적 용하여 현재 기후에서 각각의 난대성 상록활엽수종 들의 잠재서식지를 예측하였다(그림 1). 모형 예측 결 과, 센달나무와 보리장나무를 제외한 7종의 난대성 상록활엽수의 잠재서식지 면적은 20,000㎢ 이상으로 나타났다(표 1). 마삭줄은 27,766㎢로 가장 넓은 잠재 서식지 면적을 보였고, 센달나무는 15,688㎢로 가장 적은 면적을 갖는다. 모든 난대성 상록활엽수들의 분 포는 최한월최저기온과 유의한 상관관계(P<0.05)를 보였으며, 3종(보리장나무, 동백나무, 후박나무)을 제외한 나머지 종들은 겨울철강수량과도 유의한 상 관관계(P<0.05)를 나타냈다. 각 모형들의 AUC 값은 센달나무와 보리장나무가 0.9~1로 모형 예측력이 매 우 좋았으며, 줄사철나무가 0.77로 보통이었고, 나머 지 6개 종은 0.8~0.89로 좋은 정도였다(표 1).
잠재서식지 추정 결과(그림 1), 동백나무의 잠재서 식지는 주로 해안지대를 따라 좁게 분포하며 중부해
표 1. 난대성 상록활엽수의 국명, 학명 AUC value, 잠재서식지(PH), 토지이용을 고려한 잠재서식지(PHLU) 면적, 잠재서식지와 토지이용을 고려한 잠재서식지 비율
국명 학명 약어 AUC
value
Current(㎢) PHLU/PH 비율(%)
PH PHLU
동백나무 Camellia japonica CAJA 0.877 22,190 6,879 69
후박나무 Machilus thunbergii MATH 0.859 20,902 6,869 67.1
센달나무 Machilus japonica MAJA 0.916 15,688 5,002 68.1
개산초 Zanthoxylum planispinum ZAPL 0.836 23,498 7,955 66.1
마삭줄 Trachelospermum asiaticum TRAS 0.895 27,766 11,105 60
붉가시나무 Quercus acuta QUAC 0.815 29,783 10,121 66
보리밥나무 Elaeagnus macrophylla ELMA 0879 20,775 5,097 75.3
보리장나무 Elaeagnus glabra ELGL 0.900 18,245 5,135 72.1
줄사철나무 Euonymus fortunei EUFO 0.777 22,437 7,036 68.6
박선욱 ·구경아·공우석
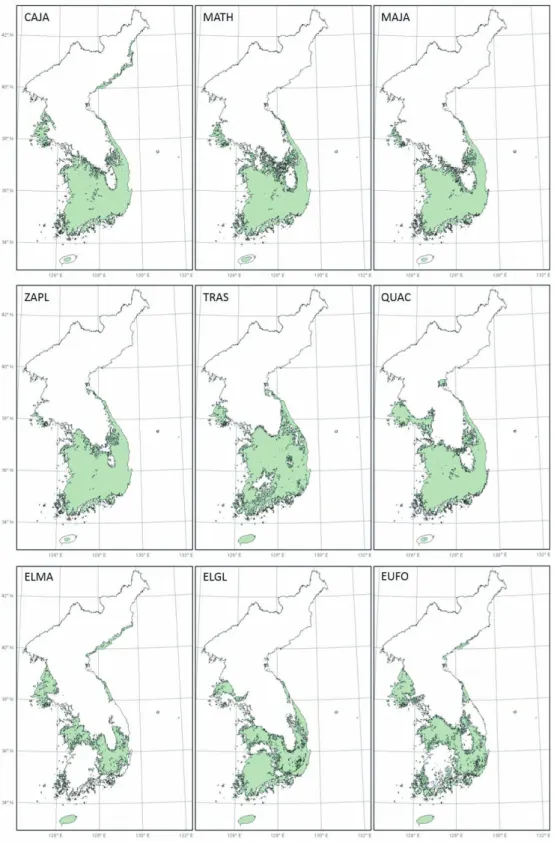
그림 1. 난대성 상록활엽수의 잠재서식지
안지역까지 나타났으며, 후박나무, 센달나무, 개산 초, 마삭줄의 대부분의 잠재서식지는 남서해안에 나 타나는 것으로 예측된다. 붉가시나무, 보리밥나무, 보리장나무, 줄사철나무의 잠재서식지는 중부해안 및 경상내륙으로 추정되었다. 이러한 잠재서식지의 분포에 따라 동백나무는 중부해안형, 후박나무, 센달 나무, 개산초와 마삭줄는 남서해안형, 붉가시나무, 보리밥나무, 보리장나무와 줄사철나무는 중부경상내 륙형 등 3개의 유형으로 나뉘었다.
중부해안형인 동백나무의 잠재서식지는 중부해안 에서 연평도와 대청도를 포함한 황해남도 해안과 덕 적도 및 안면도, 남서해안지역에서는 전라남도 내륙 부터 경상남도 해안지역까지로 예측되었다. 남서해 안형인 후박나무, 센달나무, 개산초 및 마삭줄은 전 라남도 내륙부터 경상남도 해안지역에서 잠재서식지 가 예측되었다. 중부경상내륙형인 보리밥나무, 보리 장나무, 붉가시나무와 줄사철나무는 서해 5도, 영종 도, 덕적도, 승봉도, 태안반도 등 중부해안지역 및 경 상도 내륙지역이 잠재서식지로 추정되었다.
줄사철나무를 제외한 모형예측력은 좋은 것으로 (AUC>0,8) 나타났으나 모형에 의해 추정된 잠재서식 지는 현재의 분포를 설명하는데 한계를 보였다. 예를 들어, 영동지방, 남동부해안, 경상내륙지역은 기후요 소가 적합한 동백나무의 잠재서식지로 식별되었지만 실제 분포지가 나타나지 않는다. 줄사철나무는 붉가 시나무, 보리밥나무, 보리장나무 등과 잠재서식지가 유사하게 나타났지만, 이들과 달리 충청내륙지역에 실제 분포하는 것으로 나타났다.
이러한 예측과 실제 분포의 불일치는 이 연구에서 고려되지 않은 고도, 사면, 토양 등을 포함하는 물리 적 환경요인들과 다른 연관 종들과의 상호작용들을 포함하는 생물학적 환경요인들 때문으로 판단된다.
특히, 고도, 사면 등의 지형적인 인자들은 미기후환 경에 영향을 주어 식물의 생육에 직접적인 영향을 주 는 것으로 알려져 있다(이우철·임양재, 2002). 또한, 외래종 및 교란종의 침입 및 병충해의 영향은 한 식물 의 실재 분포를 결정하는데 중요한 요인이 된다. 한라 산 구상나무의 경우 제주조릿대의 확산으로 서식지 의 축소가 우려되고 있으며(송국만 등, 2014), 소나무
의 경우 소나무 재선충의 확산에 의해 그 서식지가 심 각하게 훼손되고 있다(김종원, 2005).
2) 현재기후에서의 토지이용을 고려한 잠재서식 지 예측
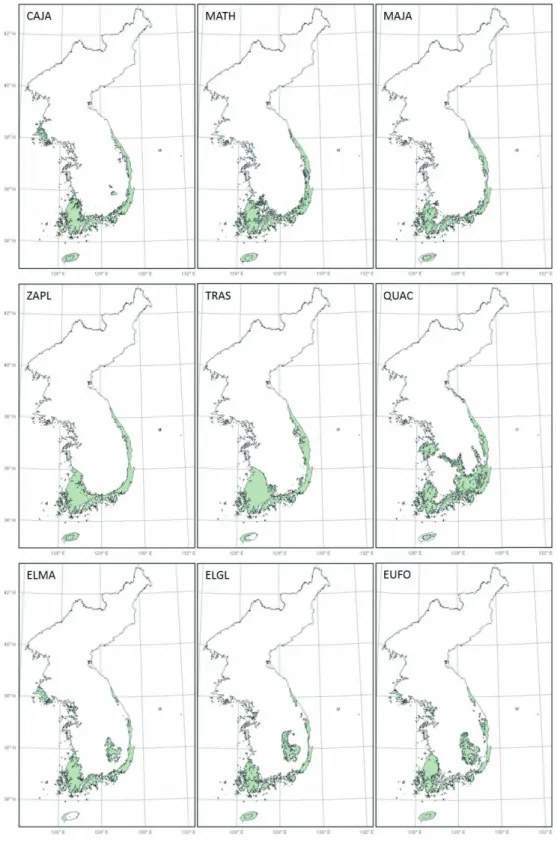
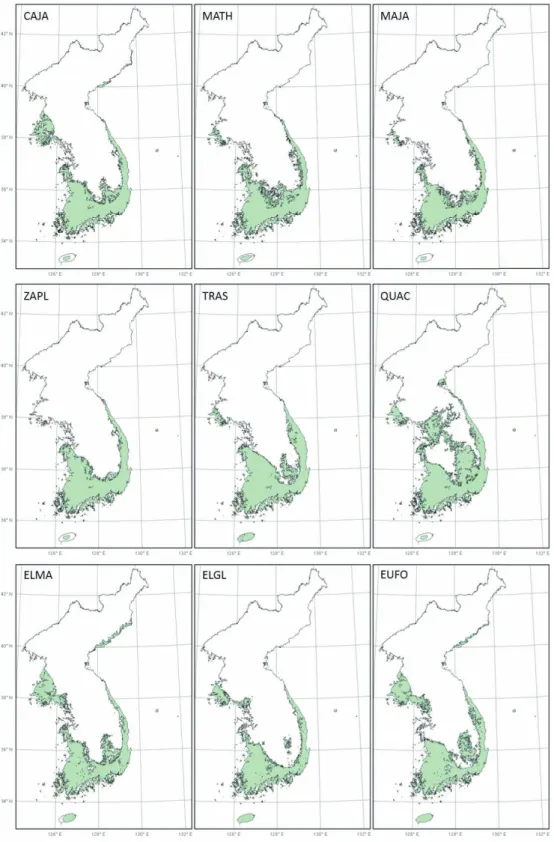
난대성 상록활엽수들의 잠재서식지 면적은 토지이 용을 고려했을 때 모두 60% 이상 감소하는 것으로 나 타났다(표 1, 그림 2). 특히, 중부경상내륙형에 속하 는 종들의 잠재서식지 감소율이 69% 이상으로 다른 분포 유형에 비해 높은 것으로 나타났다(표 1).
동백나무, 후박나무, 그리고 센달나무의 잠재서식 지는 비산림지역인 해안지역에 주로 위치하며, 토지 이용을 고려했을 때 잠재서식지가 각각 69%, 67%, 68% 감소하여 6,879㎢, 20,902㎢, 15,688㎢로 추정 되었다. 개산초와 마삭줄은 잠재서식지가 내륙지역 까지 올라오며, 토지이용을 고려한 잠재서식지 면적 은 각각 66%, 60% 감소된 23,498㎢, 27,766㎢로 다 른 종들에 비해 다소 낮은 감소율을 보였다. 붉가시나 무는 산림이 주로 나타나는 경상남도 내륙지역이 잠 재서식지로 토지이용을 고려하였을 때 잠재서식지 면적이 66% 감소한 10,121㎢로 추정되었다. 보리밥 나무, 보리장나무, 줄사철나무의 잠재서식지는 비산 림지역이 주로 나타나는 해안지역에 위치하며, 토지 이용을 고려한 잠재서식지의 면적은 각각 72%, 75%, 69% 감소한 5,097㎢, 5,135㎢, 7,036㎢로 다른 수종 들에 비해 높은 감소율을 보였다.
GAM에 의해 예측된 잠재서식지가 해안지역에 나 타나는 동백나무, 후박나무, 센달나무, 보리밥나무, 보리장나무 그리고 줄사철나무의 경우, 토지이용을 고려하였을 때 개산초와 마삭줄에 비해 잠재서식지 의 감소율이 매우 높았다. 이는 난대성 상록활엽수 가 주로 분포하는 남부해안 및 도서지역은 농경활동 등 인위적인 영향으로 산림이 파괴되고 파편화되었 기 때문으로 판단된다. 토지이용을 고려할 때 나타나 는 60% 이상의 잠재서식지 감소율은 인간활동에 의 한 직접적인 서식지 파괴가 다른 환경적인 인자들보 다 난대성 상록활엽수의 분포를 결정하는데 더 큰 영 향을 미칠 수 있음을 보여준다.
박선욱 ·구경아·공우석
그림 2. 난대성 상록활엽수의 토지이용을 고려한 잠재서식지
3) 기후변화에 따른 잠재서식지 변화
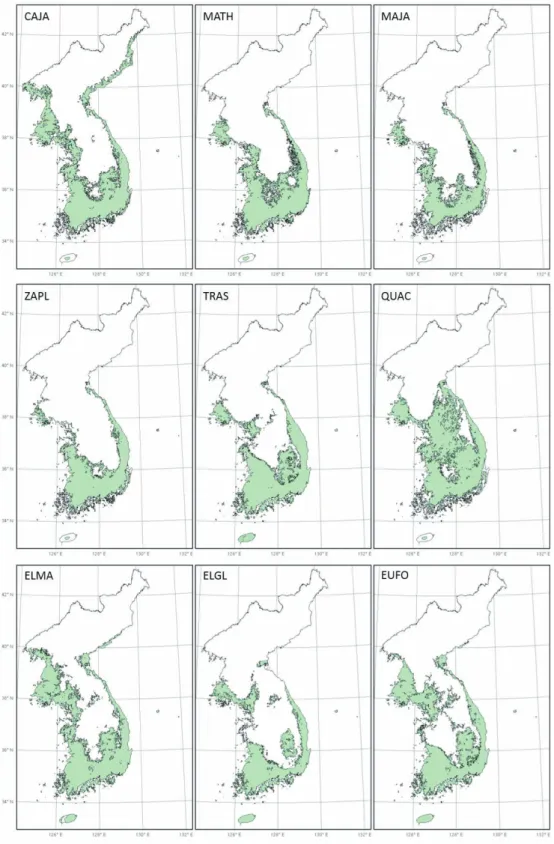
RCP 4.5와 8.5 시나리오에 따라 HadGEM2-AO 기후변화 모델을 사용하여 예측한 2050년과 2070년 의 최한월최저기온과 겨울철강수량을 적용하여, 난 대성 상록활엽수 9종의 미래 잠재서식지 변화추세를 예측하였다(그림 3~5). 예측 결과, 9개의 난대성 상 록활엽수 모두 기후변화에 따라 분포지가 북쪽으로 확대되는 것으로 나타났다. 그러나 기후변화 시나리 오나 분포 유형에 따라 분포 면적의 확대 정도가 다르 게 나타났다(그림 3).
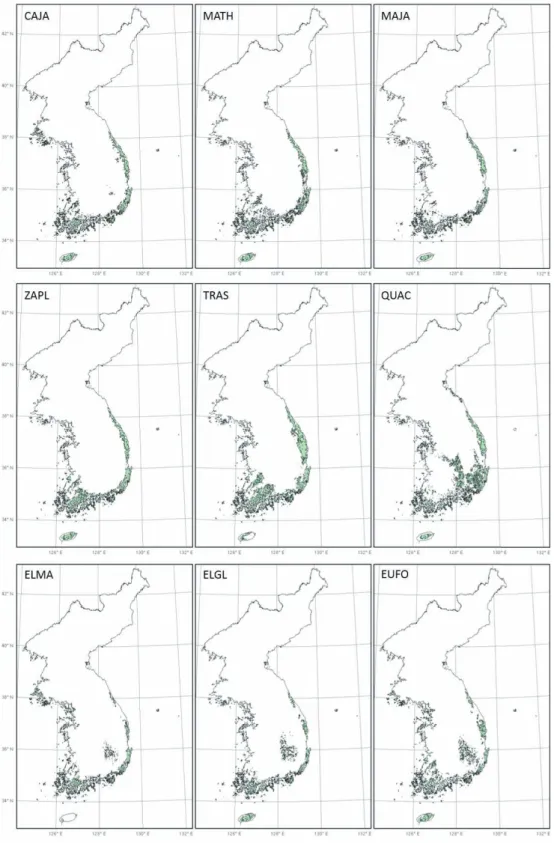
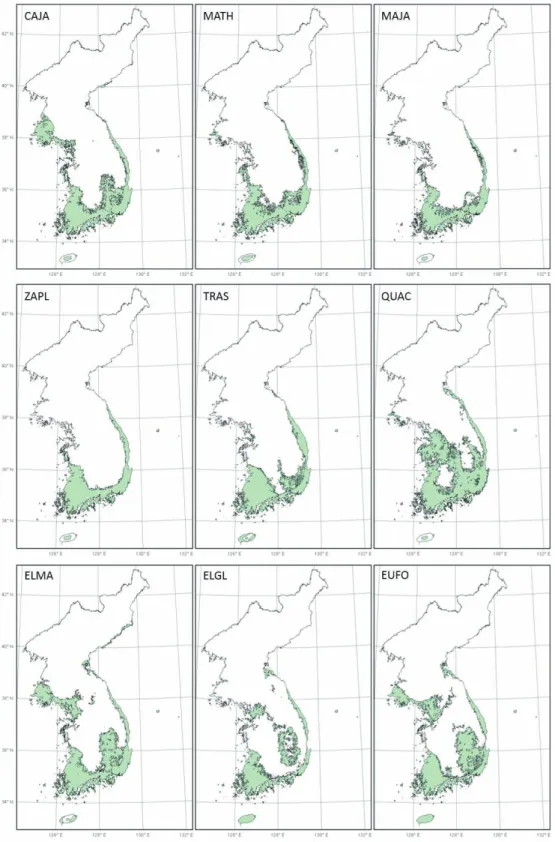
RCP 8.5는 모든 분포 유형들에 대하여 2050년과 2070년에 잠재서식지의 점진적인 확대를 보여주었다 (그림 5). RCP 4.5는 2050년과 2070년 모두에 중부 해안형과 남서해안형의 잠재서식지가 점진적으로 확 대될 것으로 예측하였으나, 중부경상내륙형은 2050 년에는 증가하나, 2070년에는 보리밥나무와 줄사철 나무의 잠재서식지가 남동부지역에서 사라질 것으로 예측하였다(그림 4). 또한, 전반적인 잠재서식지의 확장 정도는 2050년에는 RCP 4.5보다는 RCP 8.5에 서 더 크게 나타났으며, 2070년에는 RCP 4.5에서 더 크게 나타났다(그림 3).
RCP 4.5와 8.5에 따른 분포변화 예측에서, 동백나 무는 2050년에 현재와 비교해서 황해남도 해안, 전라 내륙, 경상남도 내륙지역까지 잠재서식지가 확대되
며, 2070년에는 경기북구, 강원도 내륙과 경상북도 내륙지역을 제외한 남한의 모든 지역이 잠재서식지 로 예측되었다. 특히 RCP 8.5에 따른 예측에서 함경 도, 평안도, 황해도지역의 해안지역까지 잠재서식지 가 북상할 것으로 나타났다.
남서해안형인 후박나무, 센달나무, 개산초, 마삭줄 의 경우, RCP 4.5와 8.5에 따른 예측치의 차이가 거 의 나타나지 않았다. 이들의 2050년 잠재서식지는 현 재와 비교해서 남부해안과 내륙으로 확대되며, 2070 년에는 전라도와 경상남도 내륙지역이 잠재서식지로 예측되었다. 하지만 4개종 모두 잠재서식지가 중부내 륙지역까지 확대되지는 못할 것으로 예측되었다.
RCP 4.5에 따른 예측에서 중부경상내륙형인 붉 가시나무, 보리밥나무, 보리장나무, 줄사철나무의 2050년 잠재서식지는 기존 경상남도 내륙지역과 전 라해안에서 더 내륙으로 확대되는 것으로 추정되었 다. 하지만 2070년에는 경상남도 내륙지역의 잠재서 식지는 감소하고 전라내륙지역의 잠재서식지는 확대 되는 것으로 예측되었다. 반면 RCP 8.5에 따른 예측 에서 붉가시나무는 2050년에 현재와 비교해서 남부 지역, 황해도와 경기도 해안과 내륙지역까지 잠재서 식지가 확대되는 것으로 추정되었고, 2070년에는 남 한의 대부분 지역과 황해도와 북한의 강원도지역까 지 확대되는 것으로 나타났다. 보리밥나무, 보리장나 무, 줄사철나무는 2050년에 현재와 비교해서 전라도
0 10,000 20,000 30,000 40,000 50,000 60,000 70,000 80,000 90,000
CAJA MATH MAJA ZAPL TRAS QUAC ELMA ELGL EUFO
잠재서식지면적(㎢)
Current RCP 4.5 2050 RCP 4.5 2070 RCP 8.5 2050 RCP 8.5 2070 그림 3. 현재와 미래의 난대성 상록활엽수 잠재서식지 면적 변화
박선욱 ·구경아·공우석
그림 4-1. RCP 4.5의 2050년 난대성 상록활엽수의 잠재서식지 변화
그림 4-2. RCP 4.5의 2070년 난대성 상록활엽수의 잠재서식지 변화
박선욱 ·구경아·공우석
그림 5-1. RCP 8.5의 2050년 난대성 상록활엽수의 잠재서식지 변화
그림 5-2. RCP 8.5의 2070년 난대성 상록활엽수의 잠재서식지 변화
박선욱·구경아·공우석
내륙, 경상도와 황해도 해안지역까지 잠재서식지가 확대되고, 2070년에는 해안지역의 잠재서식지가 북 상하지만 내륙으로 확대 추세는 상대적으로 약한 것 으로 나타났다.
난대성 상록활엽수 9종 모두 기후변화에 따라 분포 지가 북쪽으로 확대된 것은 분포를 결정하는 가장 중 요한 요인인 기온이 지속적이고 점진적으로 상승하 였기 때문으로 판단된다. 그러나 기후변화 시나리오 나 분포 유형에 따라 분포 면적의 확대 정도가 다르 게 나타났는데(그림 3), 이것은 겨울철강수량의 시공 간적 변화 때문으로 예측된다. RCP 4.5에 따른 Had- Gem2-AO 기후변화모델의 예측에서 우리나라의 남 동부지역의 2050년 겨울철강수량 감소가 예측되었 으며, 이것은 강수량과 출현율에 유의한 상관관계를 보인 보리밥나무과 줄사철나무의 잠재서식지 감소를 설명한다. 그러나, 잠재서식지와 토지이용을 고려한 잠재서식지를 통해 인간의 토지이용을 고려할 때, 인 간활동에 의한 직접적인 서식지 파괴는 기후변화에 의해 나타나는 난대성 상록활엽수의 서식지 확대 추 세를 감소시킬 것으로 판단된다. 또한 HadGem2-AO 단일기후변화모델을 통한 예측은 불확실성을 가지고 있기 때문에 향후 연구에서 다양한 기후변화모델을 결합한 앙상블 모형을 이용한 예측이 필요할 것으로 사료된다.
4. 결론
본 연구에서는 한반도 남부에 주로 나타나는 난대 성 상록활엽수의 분포에 대한 기후변화의 영향을 예 측하였다. 이를 위해 분포지가 중부해안까지 나타나 는 난대성 상록활엽수 9종의 분포 자료를 수집하였 고 지점별 최한월최저기온과 겨울철강수량을 통해 GAM알고리즘을 사용하여 종분포모형을 구축하고 잠재서식지를 추정하였다. 또한 토지이용을 고려하 여 모형예측에 현실성을 높이고 인간의 영향을 평가 하였다. 9종의 난내성 상록활엽수는 잠재서식지의 분 포 패턴에 따라 3개의 유형(중부해안형, 남서해안형,
중부경상내륙형)으로 나뉘어졌으며, 토지이용을 고 려했을 때 9종 모두 60% 이상의 잠재서식지가 감소 하는 것으로 나타났다. 9개의 난대성 상록활엽수 모 두 기후변화(RCP 4,5와 8.5)에 따라 2050년과 2070 년에 분포지가 북쪽으로 확대되는 것으로 예측되었 다. 그러나 분포 유형별로 토지이용과 기후변화에 대 한 영향이 다른 것으로 추정되었다. 이것은 겨울철강 수량의 시공간적 변화와 겨울철강수량의 종별로 미치 는 영향의 정도가 다르기 때문으로 판단된다. 또한 인 간활동에 의한 서식지 파괴는 기후변화에 의해 나타 나는 난대성 상록활엽수의 서식지 확대를 감소시킬 것으로 판단된다. 본 연구결과는 기후변화에 따른 상 록활엽수의 분포 변화를 예측함으로써 기후변화에 따 른 미래의 한반도 식생구조와 기능들의 변화를 이해 하는 데 기초자료가 될 것이다. 앞으로, 기후변화에 따른 난대성 상록활엽수의 미래 서식지 예측에 정확 도를 높이기 위하여 다양한 물리적, 생물적 환경요인 들과 함께 인간의 영향들을 고려하는 것이 필요하다.
사사
본 연구는 환경부와 한국환경산업기술원 “2014001 310009(기후변화대응 환경기술개발사업)”의 지원으 로 수행되었습니다.
참고문헌
공우석, 1999, “한라산의 수직적 기온 분포와 고산식물의 온도적 범위,” 대한지리학회지, 34(4), 385-393.
공우석, 2005, “지구온난화에 취약한 지표식물 선정,” 한 국기상학회지, 41(2-1), 263-273.
구경아, 2000, “한반도 상록활엽수의 지리적 분포와 기 후요소와의 관계 -내장산 굴거리나무를 중심으 로-,” 경희대학교 대학원 석사학위논문.
국립기상연구소, 2012, IPCC 5차 평가보고서 대응을 위 한 전지구 기후변화 보고서 2012 RCP 2.6/4.5/
6.0/8.5 따른 기후변화 전망.
국립수목원, 2004, 한반도 관속식물 분포도-남해안아구.
국립수목원, 2005, 한반도 관속식물 분포도-남부아구(전 라도 및 지리산).
국립수목원, 2006, 한반도 관속식물 분포도-중·남부아 구(충청도).
국립수목원, 2007. 한반도 관속식물 분포도-중·남아구 (경상북도).
국립수목원, 2008, 한반도 관속식물 분포도-중부아구(경 기도).
국립수목원, 2009, 한반도 관속식물 분포도-중부아구(강 원도).
국립수목원, 2010a, 한반도 관속식물 분포도-남부아구 (경상남도) 및 울릉도아구.
국립수목원, 2010b, 한반도 관속식물 분포도-제주도아구.
국립수목원, 2011, 한반도 관속식물 분포도-서남해안도 서지역.
국토교통부 국토지리정보원, 2014, 대한민국 국가지도집.
권혁수, 2014, “종분포모형의 불확실성 확인을 위한 앙상 블모형 적용,” 한국지형공간정보학회지, 22(4), 47-52.
권혁수·류지은·서창완·김지연·임동옥·서민환, 2012,
“종분포모형을 이용한 히어리 서식지의 분포 특 성 연구,” 환경영향평가, 21(5), 735-743.
기상청, 2014, 기상연보2014.
김종원, 2005, “소나무재선충과 동해안 산불을 통해서 본 우리나라의 소나무, 무엇이 문제인가,” 한국생태 학회지, 28(2), 113-120.
김지연·서창완·권혁수·류지은·김명진, 2012, “전국자 연환경조사 자료를 이용한 종분포모형 연구,” 환 경영향평가, 21(4), 593-607.
박종철·양금철·장동호, 2010, “기후변화에 따른 난온대 상록활엽수림대의 이동에 관한 연구,” 건국대학 교 기후연구, 5(1), 29-41.
박현철·이정환·이관규·엄기증, 2015, “구상나무와 분비 나무 분포지의 환경 특성 및 기후변화 민감성 평 가,” 환경영향평가, 24(3), 260-277.
서민환·신영규·김정현·최태봉·노환춘·김태규·김기 대, 2006, 한반도 기후변화 진단지표 생물종 조 사, 국립환경과학원,
서창완·박유리·최윤수, 2008, “위치자료의 종류에 따른 생물종 분포모형 비교 연구,” 한국지형공간정보 학회지, 16(4), 59-64.
송국만·강영제·현화자, 2014, “한라산 구상나무림의 사 면별 식생구조와 치수발생 특성,” 한국환경과학 회지, 23(1), 39-46.
신만석·장래익·서창완·이명우, 2015, “종풍부도와 세분 화된 관리지역 비교 연구-보령시를 대상으로,”
환경영향평가, 24(1), 35-50.
윤종학·박찬호·이병윤·오경희, 2011, “기후변화에 따른 한반도 난온대 상록활엽수의 잠재 생육지 변화 예측,” 한국환경생태학회지, 25(4), 590-600.
이용호·오영주·홍선희·나채선·나영은·김창석·손수 인, 2015, “기후변화 시나리오에 의한 외래식물 실망초(conyza bonariensis)의 서식지 분포 예측,”
한국기후변화학회지, 6(3), 243-248.
이우철·임양재, 2002, 식물지리. 강원대학교 출판부.
임종환·신준환, 2005, “지구온난화에 따른 산림식생대 이동과 식물계절 변화,” 자연보존, 130, 8-17.
환경부, 2000, 국가 기후변화 생물지표 100종.
Austin, M., 2002, Spatial prediction of species distribu- tion: An interface between ecological theory and statistical modelling, Ecological Modelling, 157(2), 101-118.
BOX, E.O., CRUMPACKER, D.W. and HARDIN, E.D., 1993, A climatic model for location of plant spe- cies in Florida, USA, Journal of Biogeography, 20, 629-644.
CRUMPACKER, D.W., BOX, E.O. and HARDIN, E.D., 2001, Implications of climatic warming for con- servation of native trees and shrubs in Florida, Conservation Biology, 15(4), 1008-1020.
Colwell, R. K., Brehm, G., Cardelus, C. L., Gilman, A.
C., & Longino, J. T., 2008, Global warming, ele- vational range shifts, and lowland biotic attrition in the wet tropics, Science (New York, N.Y.), 322 (5899), 258-261. doi:10.1126/science.1162547 [doi].
Díaz-Varela, R. A., Colombo, R., Meroni, M., Calvo-Igle- sias, M. S., Buffoni, A., & Tagliaferri, A., 2010, Spatio-temporal analysis of alpine ecotones: A spa- tial explicit model targeting altitudinal vegetation shifts, Ecological Modelling, 221(4), 621-633.
Dullinger, S., Gattringer, A., Thuiller, W., Moser, D., Zim- mermann, N. E., Guisan, A., Mang, T., 2012,
박선욱·구경아·공우석
Extinction debt of high-mountain plants under twenty-first-century climate change, Nature Cli- mate Change, 2(8), 619-622.
Feeley, K. J., 2012, Distributional migrations, expansions, and contractions of tropical plant species as re- vealed in dated herbarium records, Global Change Biology, 18(4), 1335-1341.
Feeley, K. J., Silman, M. R., Bush, M. B., Farfan, W., Ca- brera, K. G., Malhi, Y., Saatchi, S., 2011, Upslope migration of andean trees, Journal of Biogeography, 38(4), 783-791.
Freeman, E. A., & Moisen, G., 2008, PresenceAbsence:
An R package for presence absence analy-sis, Jour- nal of Statistical Software, 23(11), 1-31.
Guisan, A., & Zimmermann, N. E., 2000, Predictive habitat distribution models in ecology, Ecological Modelling, 135(2), 147-186.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., &
Jarvis, A., 2005, Very high resolution interpolated climate surfaces for global land areas, Interna- tional Journal of Climatology, 25(15), 1965-1978.
Jiménez-Valverde, A., & Lobo, J. M., 2007, Threshold criteria for conversion of probability of species presence to either–or presence–absence, Acta Oecologica, 31(3), 361-369.
Kearney, M., & Porter, W., 2009, Mechanistic niche mod- elling: Combining physiological and spatial data to predict species’ ranges, Ecology Letters, 12(4), 334-350.
Kleidon, A., & Mooney, H. A., 2000, A global distribution of biodiversity inferred from climatic constraints:
Results from a process-based modelling study, Global Change Biology, 6(5), 507-523.
Koo, K. A., Kong, W., Nibbelink, N. P., Hopkinson, C.
S., & Lee, J. H., 2015, Potential effects of cli- mate change on the distribution of cold-tolerant evergreen broadleaved woody plants in the korean peninsula, PloS One, 10(8), e0134043.
Morin, X., Augspurger, C., & Chuine, I., 2007, Process- based modeling of specie’s distributions: What limits temperature tree species range boundaries, Ecology, 88(9), 2280-2291.
Nakao, K., Higa, M., Tsuyama, I., Lin, C., Sun, S., Lin, J., Tanaka, N., 2014, Changes in the potential habitats of 10 dominant evergreen broad-leaved tree species in the taiwan-japan archipelago, Plant Ecology, 215(6), 639-650.
Nakao, K., Matsui, T., Horikawa, M., Tsuyama, I., &
Tanaka, N., 2011, Assessing the impact of land use and climate change on the evergreen broad-leaved species of quercus acuta in japan, Plant Ecology, 212(2), 229-243.
Randin, C. F., Engler, R., Normand, S., Zappa, M., Zim- mermann, N. E., Pearman, P. B., Guisan, A., 2009, Climate change and plant distribution:
Local models predict high-elevation persistence, Global Change Biology, 15(6), 1557-1569.
Sakai, A., 1975, Freezing resistance of evergreen and deciduous broad-leaf trees in japan with special reference to their distributions. Japanese Journal of Ecology (Japan).
Stocker, T., Qin, D., Plattner, G., Tignor, M., Allen, S., Boschung, J. Midgley, B., 2013, IPCC, 2013: Cli- mate change 2013: The physical science basis, Contri- bution of working group I to the fifth assessment report of the intergovernmental panel on climate change.
Swets, J. A., 1988, Measuring the accuracy of diagnostic systems. Science (New York, N.Y.), 240(4857), 1285-1293.
Thuiller, W., 2003, BIOMOD–optimizing predictions of species distributions and projecting potential future shifts under global change. Global Change Biology, 9(10), 1353-1362.
Uyeki, H., 1941, Northern distribution limit of korean evergreen broadleaved trees. Acta Phytotax.Geobot, 10, 89-93.
Wood, S., 2006, Generalized additive models: An introduc- tion with R, CRC press.
Yun, J., Nakao, K., Tsuyama, I., Higa, M., Matsui, T., Park, C., Tanaka, N., 2014, Does future cli- mate change facilitate expansion of evergreen broad-leaved tree species in the human-disturbed landscape of the korean peninsula? Implication
for monitoring design of the im-pact assessment, Journal of Forest Research, 19(1), 174-183.
Zimmermann, N. E., Jandl, R., Hanewinkel, M., Kun- stler, G., Kölling, C., Gasparini, P., Ulmer, U., 2013, Potential future ranges of tree species in the Alps, Management Strategies to Adapt Alpine Space Forests to Climate Change Risks, InTech, 37-48.
Zweig, M. H., & Campbell, G., 1993, Receiver-operating characteristic (ROC) plots: A fundamental evalu- ation tool in clinical medicine. Clinical Chemistry, 39(4), 561-577.
교신: 박선욱, 33657, 충청남도 서천군 마서면 금강로 1210 국립생태원 생태연구본부 융합연구실(이메일: psu [email protected])
Correspondence: Seon Uk Park, bureau of Ecological Research, Division of Ecosustem Services & Research Planning, National Institute of Ecology, 1210, Geumgang- ro, Maseo-Myeon, Seocheun-gun, Choongnam 325-813, Korea (e-mail: [email protected])
최초투고일 2016. 1. 13 수정일 2016. 4. 4 최종접수일 2016. 4. 12