Article
동계 광양만에서 식물플랑크톤 군집구조의 수평적 분포특성과 성장에 미치는 영양염 제한 특성
백승호 1* ·김동선 2 ·현봉길 1 ·최현우 3 ·김영옥 1
1한국해양연구원 남해연구소 (656-830) 경남 거제시 장목면 장목1길 41
2한국해양연구원 기후·연안재해연구부
3한국해양연구원 해양자료정보사업단 (425-600) 경기도 안산시 상록구 해안로 787
Characteristics of Horizontal Community Distribution and Nutrient Limitation on Growth Rate of Phytoplankton during a Winter in
Gwangyang Bay, Korea
Seung Ho Baek 1* , Dong Sun Kim 2 , Bong-Gil Hyun 1 , Hyun-Woo Choi 3 , and Young Ok Kim 1
1South Sea Institute, KORDI Geoje 656-830, Korea
2Climate Change & Coastal Disaster Research Department, KORDI
3Ocean Data Management Team and Information Division, KORDI Ansan P.O. Box 29, Seoul 425-600, Korea
Abstract : To estimate the effects of limitation nutrients for phytoplankton growth and its influences on short-term variations of a winter phytoplankton community structure, we investigated the abiotic and biotic factors of surface and bottom waters at 20 stations of inner and offshore areas from 6 to 7 February in Gwangyang Bay, Korea. Also, several algal bio-assay studies were conducted to identify any additional nutrient effects on phytoplankton assemblage using surface water for the assay. The dominant species in the bay was diatom Skeletonema costatum, which occupied more than 70% of total species in most stations (St.1-16) of the inner bay. According to a cluster and multidimensional scaling (MDS) analysis based on phytoplankton community data from each station, the bay was divided into three groups. The first group included stations from the south-western parts of Myodo lsland, which can be characterized as a semi- enclosed eutrophic area with high phytoplankton abundance. The second group included most stations from the north-eastern part of Myodo lsland, influenced indirectly by surface water currents from offshore of the bay. The standing phytoplankton crops were lower than those of the first group. The other cluster was restricted to samples collected from offshore of the bay. In the bay, silicon (Si) and phosphorus (P) were not a major limiting factor for phytoplankton production. However, since the DIN: DIP and DSi: DIN ratios clearly demonstrated that there were potential stoichiometric N limitations, nitrogen (N) was considered as a limiting factor. Based on the algal bio-assay, in vivo fluorescence values in N (+) added experiments were higher compared to control and P added experiments. Our results suggested that nitrogen may act as one of the most important factors in controlling primary production during winter in Gwangyang Bay.
Key words : winter phytoplankton community, nutrient limitation factor, cluster analysis, multidimensional scaling (MDS), Gwangyang Bay
*Corresponding author. E-mail : [email protected]
1. 서 론
광양만은 남해안에 위치한 폐쇄성 내만으로 남쪽에서는 쿠로시오의 지류인 대만난류의 영향을 받아 외해수가 유 입되고, 북쪽에서는 섬진강 기원의 담수가 년간 25억 8천 만톤이 유입되고 있다(김 1968). 광양만 주변에는 인구가 밀집되어 있고, 만 북부에는 광양제철소가 위치하며 남부 에는 여천국가산업단지가 조성되어 있다. 따라서 강우가 적은 동계에는 만 주변의 생활하수와 공업폐수가 영향을 미치고, 강우기에는 섬진강으로부터 많은 양의 담수가 유 입되어 만 전체에 저염분화와 더불어 높은 영양염 공급원 이 된다(이 등 2001).
식물플랑크톤의 영양염 제한 인자를 검토하는 방법으로 는 영양염 구성 성분비, Mesocosm 실험, Bio-assay 등이 있다(Brzezinski 1985; Fisher et al. 1992; Fujiki et al.
2004; Taylor et al. 1995). 특히 연안역에서 식물플랑크톤 성장에 대한 제한영양염을 평가하는 것은 하구역 및 인접 연안역의 부영양화 기작을 이해하는데 중요한 단서를 제 공할 수 있다. 국내에서는 영양염의 구성 성분비를 이용하 여 제한영양염을 판단한 연구는 담수호에서 많이 있으나 ( 김과 황 2004; 이 등 2008; 정 등 2008), 연안 해역에서는 극히 제한되어 있고, Algal bio-assay를 통해 식물플랑크 톤성장의 제한요인으로서 영양염류을 평가한 사례는 이 등 (2001)과 권 등 (2002)의 연구에 불가하다. 이 등 (2001) 의 보고에 의하면, 광양만에서는 일부 공단에서 유 입되는 인산염의 점오염에 의해 상대적으로 질소성분 영 양염이 제한요인이 되고, 강우기에도 섬진강 하구의 일부 지역에서는 질산염 제한 가능성을 시사하였다. 그러나 수 층혼합이 활발하게 일어나는 동계에 식물플랑크톤의 군집 구조와 영양염 제한을 평가한 사례는 미흡하다.
본 연구는 동계 광양만의 식물플랑크톤 수평적 군집구 조의 분포특성과 더불어 영양염 첨가 실험을 통하여 그들 의 성장에 영향을 미치는 필수 영양염류의 제한인자를 파 악하고자 한다.
2. 재료 및 방법
현장조사
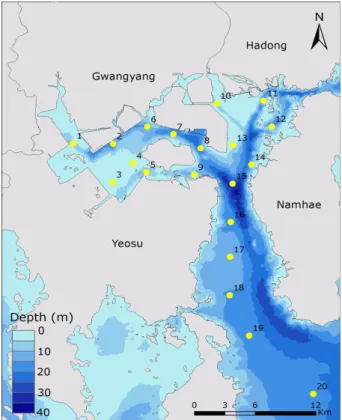
조사는 광양만에서 2010년 2월 6일-7일에 걸쳐 표층수 는 버켓으로 저층수는 니스킨 채수기를 이용하여 각각 채 수하였다(Fig. 1). 수온, 염분, pH는 수질측정기로 현장에 서 측정하였다. 투명도는 선상에서 Secchi disc를 이용하 여 측정하였다. Chlorophyll-a(Chl.a) 농도 측정을 위해서 표층과 저층수 1000 ml를 선상에서 GF/F 필터(47 mm Whatman glass fiber filters)로 여과하였고, 그 필터는 15 ml 튜브에 넣어 냉동보관하였다. 용존산소(DO)는 표층
과 저층수를 용존산소병에 채수하여 윙클러법으로 측정하 였다. 영양염 분석용 시료는 GF/F 필터를 통과한 여과 해 수 20 ml를 30 ml PC병에 넣고 냉동보관하였다. 식물플 랑크톤 종조성과 정량분석을 위해 500 ml용 채수병에 표 층수와 저층수를 각각 500 ml를 넣고 곧바로 Lugol로 최 종농도 1%로 고정했다.
Chl.a 농도는 냉동 보관한 여과지를 90% acetone에 넣 고 24시간 냉암소에서 엽록소를 추출한 후 형광측정기 (Turner Designs 10-AU Fluorometer) 로 분석하였다(Parsons et al. 1984).
영양염류는 냉동 보관한 시료를 분석전 해동하여 규산 염(SiO
2), 암모니아성 질소(NH
4-N), 아질산과 질산성 질 소(NO
2+ NO
3-N), 인산염(PO
4-P) 를 Parsons et al. (1984) 의 분석법에 따라서 Auto Analyzer(Bran Luebble)로 분석 하였다.
식물플랑크톤의 종조성과 현존량을 파악하기 위해서 500 ml의 샘플을 50 ml로 농축시킨 후 100-300 µl 분주 하여 ×20 또는 ×40배율로 동정 및 계수하였다. 특히 광 학현미경에서 동정이 극히 어려운 종은 속 수준에서 동 정하였다.
정점별 식물플랑크톤의 군집구조의 분포특성을 자세히 알아보기 위해서 각 정점에 출현한 식물플랑크톤을 바탕 으로 PRIMER version 5를 이용해 Bray-Curtis 유이도
Fig. 1. Location and water depth of study sites in
Gwangyang Bay, Korea
를 산출함과 동시에 군집(Cluster) 분석과 다차원척도 (multidimensional scaling: MDS) 분석을 수행하였다.
영양염 첨가 실험
영양염 첨가 실험은 주로 단일 배양된 식물플랑크톤의 영양염 제한을 검토하기 위해서 사용되고 있으나, 현장 식 물플랑크톤의 군집에 충분히 반영되지 않는 단점이 있다.
따라서 계절별로 출현하는 현장 식물플랑크톤을 이용하여 영양염 첨가 실험을 수행하면 그 지역의 특성을 잘 반영 할 수 있다.
식물플랑크톤을 이용한 영양염 첨가 실험은 광양만의 각 정점별 표층해수를 200 µm의 mesh로 여과하여 동물플 랑크톤의 제거한 후 1 l 멸균 채수병에 담아 암조건에 보 관하여 실험실로 운반하였다. 영양염 조절은 현장수에 영 양염을 첨가하지 않은 대조군(control), 질산염을 첨가한 질소첨가군(+N), 인산염을 첨가한 인산첨가군(+P)으로 설 정하였다. 질산첨가군은 KNO
3분말를 이용하여 첨가 후 최종농도가 20 µM로, 인산첨가군은 KH
2PO
4분말을 이용 하여 첨가 후 최종농도가 2 µM로 각각 조절하였다. 실험 은 각 정점별 현장수 50 ml을 두껑이 달린 70 ml의 시험 관(ø22 mm×200 mm; PYREX ®)에 넣고 duplicate로 수 행하였다. 배양조건은 광량 60 µmol m
−2s
−1, 광주기 12L:
12D로 조절하였고, 수온 7
oC( 조사시 현장 수온 범위)로 12 일 동안 정치 배양하였다. 영양염 제한이 언제 일어나는 가를 파악하기 위해서 2일 간격으로 Turner Designs 10- AU Fluorometer로 형광값(in vivo fluorescence)을 측정 하였고, 실험초기와 종료 시 식물플랑크톤의 군집조성을 각각 파악하였다. 또한 실험종료 시 영양염류(NO
2+ NO
3-N, PO
4-P) 농도를 상기에 언급한 방법으로 각각 측 정하였다.
3. 결 과
환경요인의 변화
동계 광양만 20개 정점에서 환경요인의 변화는 Fig. 2 와 3에 나타내었다. 투명도는 만 내측 정점에서 2 m 전후 로 극히 낮았고, 만 외측으로 갈수록 조금씩 증가했다 (Fig. 2a). 수온은 최저 4.68
oC(정점 3표층)에서 최고 7.88
oC( 정점 20저층)으로 3.2
oC 의 차이를 보였으나, 각 정 점별 표층과 저층의 수온차는 크지 않았다(Fig. 2b). 수온 의 수평적 변화는 만 내측(묘도 중심 서부해역; 정점 1-5) 에서 낮았고, 묘도의 동부해역(정점 8-13)으로 갈수록 상 승하는 경향을 보였다. 특히 정점 14을 기점으로 외측으 로 향하는 정점(15-18)에서 천천히 감소하였으나 최외측 정점(19,20)에서는 다시 증가하였다. 염분은 섬진강 하구 의 담수 영향을 직접적으로 받는 정점 10에서 최저 29.63
psu를 기록하였고, 나머지 다른 내측 정점은 32 psu 전후 로 유사한 경향을 보였으나, 정점 14를 기점으로 최외측 정점으로 향할수록 조금씩 증가하는 경향을 보였다. pH는 만 내측과 외측의 뚜렷한 차이는 관찰할 수 없었고, 점점 1 과 20에서는 낮게, 정점 10에서는 높게 나타났다(Fig. 2d).
용존산소는 정점 1의 표층에서 최저 7.37 mg l
−1를, 정점 5 표층에서 최고치 12.55 mg l
−1로 그 차이는 5.18 mg l
−1로 크게 나타났다(Fig. 2e).
각 정점에서의 영양염 농도는 규산염(DSi)이 가장 높게
관찰되었고, 다음으로 DIN(아질산염, 질산염, 암모니움),
DIP 순으로 나타났다(Fig. 3). 규산염은 정점 20의 저층에
서 14.43 µM로 최고치를, 정점 2저층에서 1.71 µM로 최
저치를 보였다(Fig. 3a). 규산염은 만 내측에서는 4-5 µM
전후로 비슷한 양상을 보였으나, 외측정점으로 향할수록
점차적으로 높게 나타났다. DIN 중 암모니움은 정점 7(표
Fig. 2. Horizontal distribution changes of transparency
(a), temperature (b), salinity (c), pH (d) and
dissolved oxygen (DO) (e) from February 6 to
February 7 2010. White and black circles indicate
surface and bottom layers, respectively
층 2.63 µM)과 9(표층 1.69 µM)를 제외하면 1 µM 이하 로 낮게 변동하였고, 아질산염 또한 모든 정점에서 0.2 µM 이하로 극히 낮게 나타났다. 따라서 DIN 농도는 질산 염 농도에 의존되었다고 판단되며, 그 농도는 담수의 영향 으로 염분이 낮은 정점 10에서 가장 높게 나타났다(13.3 µM). 이들 농도 또한 규산염 농도변동과 유사하게 외측정 점으로 향할수록 증가하였다(Fig. 3b). DIP 농도는 정점 5 저층에서 0.19 µM로 최저치, 정점 9 표층에서 1.14 µM로 최고치를 보였다. 그 중 정점 9는 비료화학공단 주변으로 비료제조과정에서 유출된 배수의 영향으로 DIP 농도가 다른 정점보다 특이적으로 높은 점오염원 현상이 나타났 으며, 이를 제외하면 표층과 저층의 차이는 크지 않았다 (Fig. 3c). 영양염 구성비중 N/P비는 정점 10에서 54로 높 게 나타났으나, 다른 정점에서는 Redfield 비 16에 못 미 치는 7-12수준으로 나타났다(Fig. 3c).
Chl.a 농도의 변동범위는 1.46-6.40 µg l
−1( 표층평균:
2.88±1.12 µg l
−1, 저층평균: 3.64±1.37 µg l
−1) 으로 저층 이 높게 관찰되었다. 특히 동천하구의 정점 1과 2에서 가 장 높게, 다음으로 섬진강하구 정점 10과 여수항부근 정 점 18에서 높게 나타났으며, 정점 11의 표층과 정점 20의 표층과 저층에서 낮게 관찰되었다.
식물플랑크톤의 군집구조
동계 광양만에서 식물플랑크톤 현존량은 정점 10에서 최고치 78×10
5cells l
−1, 정점 20에서 최저치 0.9×10
5cells l
−1를 기록하였고, 그들 군집조성의 70% 이상은 규조 류가 차지하였다(Figs. 4와 6a). 다음으로 은편모조류, 와 편모조류 순으로 나타났으며, 규질편모조류와 유글레나조 류는 출현빈도도 낮고 개체수 또한 극히 낮게 관찰되었 다. 특히 정점 1-16에서는 규조류 Skeletonema spp.가 70% 이상을 차지했다. 외해수의 영향을 받는 정점 17-19 에서는 Skeletonema spp.와 더불어 Thalassionema spp.과 Thalassiosira spp.가 각각 균등하게 우점 출현하였다(Figs.
5와 6b). 정점별 출현종수는 만내측에서 15종 내외로 관찰 되었고, 외측의 정점 17-19에서는 20종 전후로 약간 높게 관찰되었다(Fig. 7). Shannon-Wiener의 다양도지수와 균 등도 지수는 식물플랑크톤의 현존량이 가장 높게 관찰된 정점 10에서 극히 낮게 나타났다. 본 조사에서는 다양도
Fig. 3. Horizontal distribution changes of DSi (a), DIN (nitrate, nitrite, ammonia) (b), DIP, N:P ratios and Chl.a concentration from February 6 to February 7. White and black circles indicate surface and bottom layers, respectively
Fig. 4. Horizontal distribution changes of total phyto-
plankton abundance for surface layers in Gwangyang
Bay from 6 to 7 February, 2010
지수와 균등도 지수가 유사한 변동 경향을 나타났으며, 두 인자간의 상관성 또한 높게 나타났다(r=0.98, p<0.01).
조사기간 중 정점별 출현한 식물플랑크톤의 군집구조를 바탕으로 Cluster 분석과 MSD(multidimensional scaling) 분석한 결과를 Fig. 8에 나타내었다. 식물플랑크톤의 군집 구조와 그들의 출현양상에 따라서 형성되는 정점별 그룹 의 차이는 명확하게 나누어지지 않았지만, 크게 3개의 그
룹으로 구분할 수 있었다. 제1그룹은 묘도 남수로와 동천 하구의 정점이 작은 그룹으로 유사도 80%로 묶였고, 다음 으로 섬진강하구의 정점 10, 11이 차례로 묶였다(유사도 68%). 제2그룹은 묘도 북수로와 일부 정점, 중앙교차수로 ( 정점 15)와 더불어 동부해역의 대부분의 정점으로 구분되 었고(유사도 68%), 제3그룹은 해수의 교환이 원활한 외측 해역의 정점 17과 정점 18을 중심으로 구분되어졌다(유사 Fig. 5. Horizontal distribution changes of dominant phytoplankton species in field samples (A-G) and algal bio-assay
experiments (a-g)
Fig. 6. Relative contribution of phytoplankton classes (upper) and dominant diatom species (lower) during 6 to 7 February 2010 in Gwangyang Bay
Fig. 7. Horizontal distribution changes of number of observed species (a), Shannon-Wiener diversity indices (H’) (b) and evenness (J’) (c) in Gwangyang Bay from 6 to 7 February 2010
Fig. 8. Joint plots of sites association analyses obtained
by cluster (a) and non-metric multidimensional
scaling (MDS) (b) ordination analyses, using a
data set of the surface species abundances during
the study period in Gwangyang Bay
도 68%).
영양염 첨가 실험
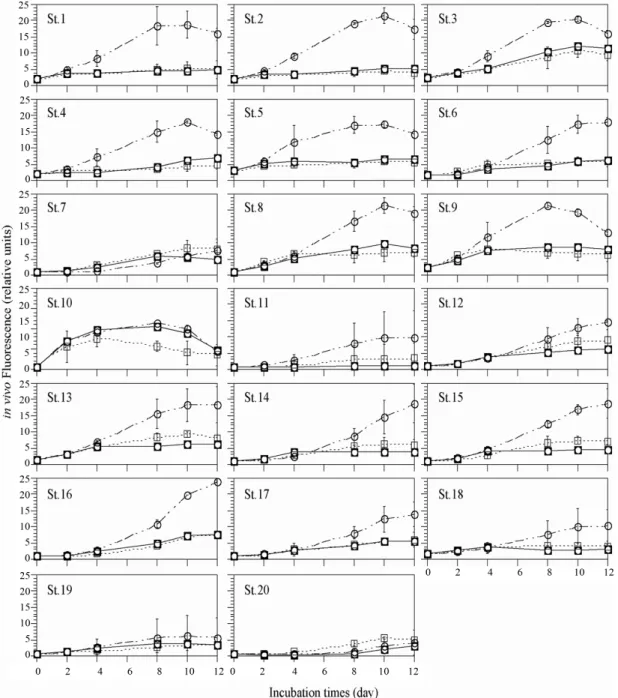
식물플랑크톤의 증식을 간접적으로 확인할 수 있는 형 광값을 이용하여 영양염 첨가에 따른 각 정점별 생물의 반응을 살펴본 결과, 대부분의 정점에서 인 첨가군 형광값 이 영양염을 첨가하지 않은 대조군의 값과 비슷하거나 그 이하로 나타났으나, 정점 11-15과 더불어 정점 20에서는 대조군보다 조금 높게 관찰되었다(Fig. 9). 반대로 질소 첨
가군에서는 배양시간의 경과와 더불어 형광값이 높게 나 타났다. 대조군에서도 시간의 경과와 더불어 형광값이 증 가하는 정점(8, 9, 10, 12, 13, 16)이 나타났고(Fig. 9), 이 들 대부분의 정점은 광양만 동부해역으로 담수의 영향을 직간접으로 받고 있는 정점이였다. 영양염 첨가의 효율을 알아보기 위해 대조군 대비 N 첨가군과 P 첨가군의 결과 를 Fig. 10에 나타내었다. 각각의 영양염첨가군의 효율은 각 정점별 12일간 배양한 형광값의 평균을 산출한 후 (Tf/
Cf-1)×100 식에 대입하여 구할 수 있었다. 여기서 Tf는
Fig. 9. Growth response of phytoplankton assemblages by algal bio-assay. These experiments are used surface water from each station in Gwangyang Bay. Control=no additions ( ); +N=nitrate addition (
○); +P=phosphate addition (
□)
■
첨가군의 형광값이며, Cf는 대조군의 형광값이다. 대조군 대비 N 첨가군에서는 정점 7과 10에서 제로 또는 부(−)의 효율을 보였고, 정점 11에서 최대 500%의 첨가효율을 나 타내었다. 나머지 대부분의 정점에서도 N 첨가에 따른 식 물플랑크톤의 증가효과를 확인하였다. 대조군 대비 P 첨 가군에서도 정점 11에서 150%로 높게 나타났으며, 다음 으로 정점 20에서 100% 이상의 효율을 보였다. 그러나 P 첨가군에서는 대조군대비 식물플랑크톤의 증가효과를 보 이지 않은 정점이 9개(2, 3, 4, 5, 8, 9, 10, 16, 19)로 나타 났으며 광양만 전체 45%의 비율을 차지했다.
4. 고 찰
광양만해역의 표층에서 환경인자간 상관성은 규산염과 투명도(Secchi depth) (r=0.87, p<0.01), 암모니움과 용존 무기질소(r=0.48, p<0.05)가 각각 유의한 관계를 보였다 (Fig. 11). 용존무기질소(DIN=아질산염+질산염+암모니 움) 농도의 대부분은 질산염 농도에 의해서 의존되었다.
연안해역에서는 영양염 각 인자간(DSi, DIN, DIP)의 상관 성이 높게 나타나는 경향이 강하지만(Justic et al. 1995;
Baek et al. 2009; 이 등 2001; 백 등 2008), 본 해역의 동
계 조사에서 DIN은 DSi와 DIP와의 상관성은 보이지 않 았다. 특히 조사해역에서 Chl.a 농도가 높게 나타나지 않 았음에도 불구하고 투명도가 2-3 m 전후로 극히 낮게 관 찰되었다. 이것은 수심이 얕은 광양만에서 동계 활발한 수 층혼합으로 해저의 미세입자가 재부유되어 입자성유기물 질의 형태로 수주내 대량 공급되어 투명도를 떨어뜨렸고, 이와 함께 공급된 영양염(퇴적층으로부터 재영양염화가 일어날 때 DIN보다 DSi, DIP 농도가 상대적으로 높음)이 각 정정별로 현저한 차이를 보여 영양염 각 인자간의 관 계도 낮았다고 추측된다.
DIN 농도는 Chl.a 농도와 상관성이 나타나지 않았고,
식물플랑크톤의 총개체수(특히 규조류의 개체수)와 상관
성도 나타나지 않았다. 섬진강 영향을 강하게 받는 정점
10 에서는 DIN 농도가 극히 높았고, S. costatum가 극우점
출현한 것이 특이적이었다(Fig. 11). S. costatum는 광온
광염성종으로 기수역과 연안해역에서 널리 분포하는 종으
로 계절에 관계없이 빈번히 출현한다. 그들은 여름철 고온
저염분 수괴에 자극을 받아 빠른 성장을 하는 것으로 보
고되고 있으나(Han et al. 1992; 백 등 2010), 본 조사에서
는 동계 낮은 수온조건(5-8
oC) 에서도 잘 적응하였으며, 특
히 정점 10에서 높은 질소기원의 영양염과 더불어 저염분
Fig. 10. Comparision of N and P nutrient additional efficiency vs. control based on Fig. 8
화에 자극받아 현저하게 증식하였을 것으로 판단된다.
Chl.a 농도와 식물플랑크톤 개체수와의 상관성은 상대적 으로 낮았고(r=0.45, p<0.05), 이는 식물플랑크톤의 각 종 마다 세포 크기가 다르고 세포내 함유하고 있는 Chl.a의 양적 차이 때문에 기인된 것으로 생각된다. 결과적으로 S.
costatum는 동계 광양만의 식물플랑크톤의 군집에 중요한 위치를 차지하는 종으로 구분되었고, 이들 종은 저수온 환 경하에서도 질소원의 영양염을 효율적으로 이용한 결과 다른 경쟁자보다 빠르게 성장할 수 있었다고 판단된다.
식물플랑크톤의 군집구조에 따른 Cluster 분석과 MDS 분석을 수행한 결과, 정점별 그룹의 차이는 명확하게 나누 어지지 않았지만, 크게 3개로 구분할 수 있었다. 제1그룹 은 산업시설이 밀집된 폐쇄성 내만해역으로 S. costatum 단일종의 현존량이 극히 높았고, 제2그룹은 묘도를 중심 으로 북동부해역으로 식물플랑크톤 현존량이 상대적으로 낮게 관찰된 곳으로 구분되었고, 제3그룹은 해수의 교환 이 원활한 외측해역을 중심으로 식물플랑크톤의 현존량도 낮으면서 종 다양도 지수와 균등도 지수가 높은 해역으로 나누어졌다. 이 등 (2007)은 광양만 유·무기물 분포 특성 과 해수의 흐름 등을 고려하여 주요 오염원의 영향권을 만내측에서 3개 권역으로 구분한 바가 있다. 제 1해역은 남북으로 하동 갈사에서 여수해만 입구(중앙교차수로)와 동서로는 광양제철소 끝부분에서 광양만 동부의 전해역으 로, 제2해역은 광양만 동천의 영향을 받고 묘도 안쪽의 해 역으로, 제3해역은 여수여천산업단지의 영향을 많이 받는 해역으로 구분하였다. 식물플랑크톤의 군집구조를 바탕으 로 한 본 연구결과에서도 선행연구와 유사하게 만 내측의 묘도를 중심으로 서부폐쇄성 해역과 동부해역으로 크게 2 개로 구분되었다. 이와 같이 구분된 식물플랑크톤의 분포 특성은 광양만 표층 해류의 흐름과 밀접한 관계가 있을 것으로 사료되었다. 해수의 흐름을 파악할 수 있는 수온의 분포특성에서도 서부해역보다 동부해역이 외양수의 유입 이 반영되었는 듯 평균 2-3
oC 로 높게 나타났다. 광양만
표층해류는 여수해만에서 광양항으로 유입되어 서쪽으로 는 묘도의 남북수로를 통해 내만으로 흐르고, 북쪽으로 는 대도군도 양측의 좁은 수로를 통과한 주류는 노량대 교를 거쳐 진주만으로 강하게 흘러 들어가고, 일부 지류 는 마도수로를 통과하면서 회귀하여 섬진강하구 주변에 영향을 미치며 바깥쪽 외해로 빠져나간다(National Oceanographic Research Institute 2002; 이 등 2007).
MDS 분석에서 제1그룹의 대부분의 정점은 표층 해류 이 동이 상대적으로 약화되는 곳으로서 해류 흐름이 빠른 동 부해역과 외측의 정점보다 식물플랑크톤의 분산 및 확산 이 낮았을 가능성이 크다. 생태학적으로 식물플랑크톤의 대발생기작은 환경요인에 대한 종 특이성에 따른 성장률 뿐만 아니라, 개체군 소멸 및 소실에도 영향을 미친다 (Smayda 1997). 그들 개체군 소멸은 상위영양단계의 포식 압과 더불어 난류(water turbulence)나 해류로 인한 수주 내 분산과 다른 지역으로 확산되어 일어난다(Margalef 1978; Smayda 2002). 결과적으로 수심이 깊고 해류의 이 동이 빠른 그룹 2와 3의 정점은 식물플랑크톤의 현존량이 상대적으로 낮게 나타났고, 그 원인은 동계 활발한 수층혼 합과 지형학적 특성에 따른 표층수 흐름의 차이로 발생된 것으로 생각되며, 이는 식물플랑크톤 군집구조에 따른 해 역의 구분에서도 잘 나타났다.
연안해역에서 영양염 구성성분 비의 변화는 부유생물의 성장과 종간 경쟁을 유도해 종천이를 유발하는 요인이 된 다(Goldman et al. 1979; Fisher et al. 1992). Dortch and Whitledge (1992) 의 보고에 의하면, 대부분의 식물플랑크 톤의 증식에 대한 영양염 제한은 DIN이 1.0 µM, DIP가 0.2 µM, DSi가 2.0 µM 이하로 관찰되는 해역에서는 영양 염 절대농도 하한값의 제한을 받고, 규조류의 성장은 Si:N 비가 1:1일 때 Si의 제한을 받으며 그들의 구성비가 1이하 의 환경이 지속되면 규조류가 성장할 수 없어 종 천이을 유발한다(Turner et al. 1998). 일반적으로 Redfield 비는 Si:N:P=16:16:1로 평가되고 있으나(Redifeld et al. 1963), Fig. 11. Relationships between abiotic and biotic factors. (a) Secchi depth and silicate. (b) DIN and nitrate, ammonium.
(c) DIN and Chl.a concentration. “ns” indicates statistically not siginificant
Justic et al. (1995) 이 제시한 상호복합적으로 연관되는 화 학량론적 제한(stoichiometric limitation)은 다음과 같다. P 제한은 DSi:DIP>22 또는 DIN:DIP>22이며, N 제한은 DIN:DIP<10 또는 DSi:DIN>1이며, Si 제한은 DSi:DIP<
10 또는 DSi:DIN<1으로 평가하고 있다. 이 기준치에 따 라 동계 광양만에 적용한 결과를 Fig. 12에 나타내었다.
본 조사시에는 식물플랑크톤의 성장에 필요한 영양염의 하한치 농도와 구성 성분비에 따른 제한은 Si와 P에 대해 서 각각 1회씩 적용되었을 뿐 다른 대부분의 정점에서는 제한을 받고 있지 않는 것으로 평가되었다. 하지만 N 영 양염에 관해서는 하한치농도의 제한은 관찰되지 않았으 나, 영양염 구성성분비에 따른 제한은 조사정점의 60%
전후로 높게 나타났다. 이와 같은 결과는 광양만 내만에서 담수유입이 적은 시기에 질소기원의 영양염 제한이 현저 하게 일어난다는 이 등 (2001)의 보고와 유사하다. 그들의 결과에 의하면, 강우로 인한 질소기원의 영양염이 대량 유 입되는 시기조차도 인산염의 제한은 명확하게 나타나지 않았다. 본 연구의 영양염 첨가 실험에서도 대조군 대비 P 첨가군에서는 정점 11과 20을 제외한 나머지 정정에서 50% 이하을 보였거나, 부(−) 첨가 효율을 보였다. 반면 N 첨가군에서는 배양시간의 경과와 더불어 식물플랑크톤의 증식이 현저하게 나타났다. 또한, DIN 농도가 극히 낮게 관찰된 정점 2, 3, 4, 5는 P 첨가의 효과를 관찰할 수 없었 다. 즉 DIN 농도가 낮은 환경하에서는 추가적으로 P 영양 염을 첨가하여도 대조군대비의 효율이 나타나지 않았다.
하지만, 정점 11의 경우는 예외로 N, P 첨가군의 효율이 극히 높게 관찰되었다. 영양염 첨가 실험에서 정점 11은 다른 정점과 대조적으로 실험기간동안 대조군의 성장은 거의 이루어 지지 않았고(Fig. 9), 상대적으로 N과 P의 첨가군에서는 식물플랑크톤의 성장이 관찰되어 대조군대 비의 효율이 각각 500과 150%로 높게 관찰되었다.
Goldman et al. (1979)에 의하면, 대부분의 식물플랑크톤 은 수주내에 높은 농도의 인이 존재하면 그것을 과량섭취 한 후 인이 부족한 환경이 되었을 때 과량섭취한 인을 이 용하여 일정기간 동안 성장한다고 보고했다. 현장에서 낮 은 농도 인이 검출된 정점에서도 영양염 첨가 실험의 N 첨가군에서 높은 형광값을 관찰할 수 있었던 것은 과량섭 취한 인을 이용하여 일정기간 동안 성장했을 가능성이 크 다. 결과적으로 광양만에서 P 영양염의 첨가효과가 낮게, N 영양염의 첨가효과가 높게 나타난 것은 N 영양염에 의 해 그들 식물플랑크톤 성장에 중요한 영향을 미친 것으로 판단되며, N 영양염의 제한은 동계 광양만의 일차생산력 을 저하시키는 요인으로 생각된다.
영양염 첨가 실험 후 남은 영양염 농도를 파악하면 식 물플랑크톤 성장의 제한 영양염인자를 구체적으로 파악할 수 있고, 각 정점의 소비 후 남은 영양염 농도를 Table 1 에 나타내었다. 영양염을 첨가하지 않은 대조군에서는 정 점 11, 20을 제외하면 모든 정점에서 N과 P의 영양염 농 도가 극히 낮게 관찰된 것으로 보아 N과 P 영양염의 제한 이 식물플랑크톤 성장의 저해요인으로 파악된다. 특히 동 Fig. 12. Scatter diagrams of nutrient ratios at all station
in Gwangyang Bay. Stoichiometric limitation are
indicated as -N, -P, and -Si. Snow symbol and
open circles are surface and bottom layers,
respectively (n=40)
계 광양만 광역해역에서 대부분 정점의 NP비는 약 10전 후로 관찰되었으나, 실험후 대조군의 남은 영양염 농도의 NP비는 0.5-4 수준에 머물러 있었으며, 이것은 현장에서 추가적인 영양염 공급이 이루어지지 않는다고 가정하면 N 영양염류의 소비률이 P보다 높게 나타나 결과적으로 N 영양염제한이 일어날 것으로 추측할 수 있다. N 첨가군에 서는 대부분의 정점에서 대조군 대비 식물플랑크톤의 성 장이 현저하게 나타났고, 추가한 N 영양염은 일정량 소비 된 후 P 영양염의 제한으로 더 이상 소비되지 않는 경향 을 관찰할 수 있었다. 하지만, N 첨가군에서는 정점별로 사용후 남은 N 영양염의 현저한 차이를 보였고, 이는 P 첨가군에서 나타난 N의 영양염 고갈 현상과 남은 P 영양 염농도의 변화에서도 확실하게 구분되었다. 앞서 언급했 드시 동계 광양만에서는 화학량론적 제한에서도 P보다는 N의 영양염이 현저하게 제한된다는 것을 알 수 있었고, 영양염 첨가 실험에서도 N 영양염의 소비된 농도가 높은 것으로 보아 P보다 N 영양염이 제한 요인이 될 수 있다는 것을 가늠할 수 있었다. 결과적으로 본 실험과 현장조사를 통하여 동계 출현한 식물플랑크톤의 군집에서는 N-uptake
가 압도적으로 높게 나타났고, 이것은 동계 광양만에서 N 영양염의 제한이 두드러지게 나타날 수 있다는 것을 시사 한다.
N-첨가군에서 질산염과 용존무기인의 소모량은 각각 평균 15.70 µM와 0.31 µM로 나타났고, 대조군에서는 각 각 3.59 µM, 0.31 µM로 나타났다. N-첨가군과 대조군 사 이 용존무기인의 농도는 유사한 값을 보인 반면, 대조군대 비 N-첨가군의 엽록소 증가 효율은 약 125%로 극히 높게 나타났다. 여기서 용존무기인의 소모률의 차이 없어 엽록 소 증가 효율이 높게 나타난 것에 관하여 의문을 가질 수 있고, 그 의문에 두 가지의 가능성을 제시할 수 있다. 먼저 대조군에서 용존무기인의 고갈 원인을 밝히면 중요한 단 서를 제공할 수 있다. 영양염을 첨가하지 않은 현장영양염 농도의 대조군에서는 일정 기간(4-6일) 성장후 N 영양염 이 제한되었을 것으로 사료되는 시점부터 식물플랑크톤 성장의 평형을 이루었다(Fig. 9). 이 과정에서 식물플랑크 톤의 증식은 이루어지지 않았지만, 진핵식물플랑크톤의 P 의 저장고에 용존무기인을 저장하여 P가 일정량 소비되어 실험 최종일 N 첨가군과 대조군에서 P 영양염의 차이가 없게 나타났을 것이다. 진핵식물플랑크톤은 인산염 에스 테르 화합물을 세포내 축적하는 생물저장고를 가지고 있 고, 일정량 P 영양염을 비축할 수 있는 역할을 담당한다 (Yamaguchi and Adachi 2010). 다음으로 N 첨가군에서 영양염 소비경향을 파악하면 된다. N첨가군은 일정기간 배양후 식물플랑크톤이 기하급수적으로 증식하여 첨가된 N 영양염 보다 첨가하지 않은 용존무기인이 먼저 고갈되 었을 가능성이 있다. 각 정점마다 차이는 보였지만, 배양 8-10 일을 전후해서 성장곡선이 평형을 이루거나 하강하는 경향을 관찰할 수 있었는데, 이것은 계(system)내의 용존 무기인의 고갈 후 용존상태의 유기인이나 생물세포내 저 장고에 비축한 P을 사용하여 지속적으로 성장을 유지하였 을 가능성이 높다. 추후 이와 같은 가능성을 보다 명확하 게 증명하기 위해서는 식물플랑크톤 세포내 N과 P 함유 량을 파악하는 연구가 필요하다.
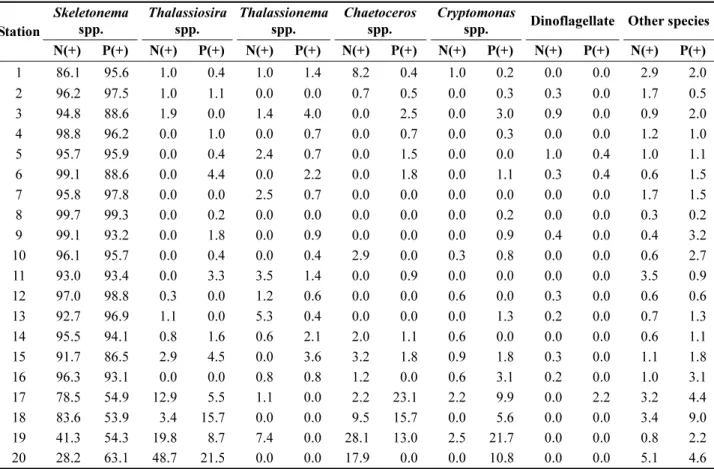
현장에 우점 출현한 종이 영양염 첨가 실험에서도 높은 점유률을 보였고, 각각의 종의 개체수의 증가률은 초기농 도의 10-100배 정도 높게 나타나는 경향을 관찰하였다 (Fig. 5와 Table 2). 특히 규조류 Skeletonema spp.(주로 S. costatum)가 우점 출현한 곳(정점 1-16)에서 그들의 점 유률은 85% 이상으로 높게 차지했다. 현장관측에서 S.
costatum과 더불어 Thalassionema nitzschioides, Thalassiosira spp. 가 높게 출현한 해역(정점 17-20)에서 동일종이 높은 점유률(80% 이상)을 보인 것을 영양염 첨가실험으로 확 인할 수 있었고, 다음으로 Chaetoceros spp., 은편모조류 가 일정량 차지하였다(Table 1). 이와 같이 현장에서 우점 출현한 종이 영양염 첨가 실험에서도 높은 점유률을 차지 Table 1. Remained nutrient concentrations after algal
bio-assay of N and P additional experiments.
The sample was collected in final days of experiment
Contorl N(+) Exp. P(+) Exp.
NO
2+NO
3PO
4NO
2+NO
3PO
4NO
2+NO
3PO
4µM µM µM µM µM µM
1 0.16 0.04 6.63 0.02 0.20 1.08
2 0.07 0.04 4.06 0.03 0.04 1.51
3 0.05 0.02 3.60 0.04 0.06 1.03
4 0.08 0.03 4.94 0.06 0.01 1.26
5 0.07 0.03 5.23 0.09 0.04 1.42
6 0.08 0.03 8.92 0.06 0.05 1.28
7 0.05 0.04 19.25 0.30 0.04 1.56
8 0.04 0.03 7.07 0.06 0.05 1.49
9 0.06 0.06 0.61 0.09 0.19 1.52
10 0.06 0.09 12.83 0.09 0.12 0.70
11 0.12 0.13 9.68 0.04 0.14 1.60
12 0.08 0.05 11.22 0.04 0.14 1.48
13 0.05 0.02 9.14 0.04 0.14 1.39
14 0.05 0.02 11.31 0.06 0.04 1.14
15 0.02 0.04 4.16 0.07 0.08 1.53
16 0.06 0.03 5.85 0.11 0.34 2.02
17 0.03 0.08 4.05 0.07 0.06 1.23
18 0.01 0.02 13.49 0.11 0.08 1.58
19 0.04 0.09 7.27 0.03 0.09 2.03
20 0.57 0.15 13.45 0.04 0.08 1.58
할 수 있었던 것은 계(System)내 영양염 경쟁에 따른 종 천이 현상이 두드러지게 나타나지 않았고, 각 정점에서 출 현한 우점종의 초기 접종 밀도에 의한 영향이 컸다. 하지 만, N 첨가군의 Chaetoceros 속의 개체수 밀도가 정점 1 과 정점 19에서 현저히 증가하였고, 그 원인은 파악할 수 없었다. 본 연구에서 광양만은 규조류가 차지하는 비중이 극히 높았고, 그들의 분해로 인하여 Si 영양염의 재순환과 담수로부터 공급된 높은 영양염은 규조류가 우점하기 좋 은 환경이라 사료된다. 특히 광양만과 같이 표층수 흐름이 강하고 수층혼합이 심한 해역은 와편모조류보다 규조류가 우점하기에 유리한 조건이라 할 수 있다.
5. 요약 및 결론
동계 광양만에서 식물플랑크톤 군집구조와 그들의 성장 에 영향을 미치는 영양염제한 특성을 파악하기 위해서 만 내외측의 20개 정점에서 생물학적 요인과 무생물학적 요 인을 조사했다. 또한 식물플랑크톤 군집에 대한 영양염 제한 효과를 알아보기 위해서 실험실에서 현장 20개 정 점의 표층수를 이용하여 영양염 첨가 실험을 수행하였
다. 만내측의 대부분의 정점(1-16)에서 규조류 Skeletonema costatum이 극우점하였고, 그들은 전체 식물플랑크톤 개체 수의 70% 이상을 찾지하였다. 각 정점에서 출현한 식물플 랑크톤의 군집조성을 바탕으로 한 Cluster와 MDS 분석에 의하면, 광양만은 3개의 그룹으로 구분되었다. 제1그룹은 묘도를 중심으로 서부해역에 위치한 정점으로 구성되었 고, 이들 정점의 특성은 반폐쇄성 내만으로서 식물플랑크 톤의 개체수 밀도가 높았다. 제2그룹은 묘도 북부와 동부 해역에 위치한 대부분의 정점으로 구분되었고, 이들 정점 은 광양만 외해로부터 유입되는 표층해류의 영향을 강하 게 받아 식물플랑크톤 개체수가 수주내 분산과 다른 지역 으로 확산되어 그들의 현존량이 만내측의 그룹1보다 낮게 나타났다. 나머지 다른 그룹은 만외측에 위치한 정점으로 구분되었다. 본 조사에서 Si와 P는 식물플랑크톤의 증식 에 제한 요인이 되지 않았으나 N은 DIN: DIP 비와 DSi:
DIN 비에서 잠재적 제한요인으로 고려되었다. 영양염 첨 가 실험에서는 N 첨가군은 대조군과 P 첨가군보다 높은 형광값을 나타내었다. 결과적으로 N 기원의 영양염은 동 계 광양만의 기초생산력을 제어하는 중요한 요인 중 하나 로 판단되었다.
Table 2. Relative contribution (%) of main phytoplankton species by algal bio-assay of N and P additional experiments. The sample was collected in final days of experiment
Station
Skeletonema spp.
Thalassiosira spp.
Thalassionema spp.
Chaetoceros spp.
Cryptomonas
spp. Dinoflagellate Other species
N(+) P(+) N(+) P(+) N(+) P(+) N(+) P(+) N(+) P(+) N(+) P(+) N(+) P(+)
1 86.1 95.6 1.0 0.4 1.0 1.4 8.2 0.4 1.0 0.2 0.0 0.0 2.9 2.0
2 96.2 97.5 1.0 1.1 0.0 0.0 0.7 0.5 0.0 0.3 0.3 0.0 1.7 0.5
3 94.8 88.6 1.9 0.0 1.4 4.0 0.0 2.5 0.0 3.0 0.9 0.0 0.9 2.0
4 98.8 96.2 0.0 1.0 0.0 0.7 0.0 0.7 0.0 0.3 0.0 0.0 1.2 1.0
5 95.7 95.9 0.0 0.4 2.4 0.7 0.0 1.5 0.0 0.0 1.0 0.4 1.0 1.1
6 99.1 88.6 0.0 4.4 0.0 2.2 0.0 1.8 0.0 1.1 0.3 0.4 0.6 1.5
7 95.8 97.8 0.0 0.0 2.5 0.7 0.0 0.0 0.0 0.0 0.0 0.0 1.7 1.5
8 99.7 99.3 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.3 0.2
9 99.1 93.2 0.0 1.8 0.0 0.9 0.0 0.0 0.0 0.9 0.4 0.0 0.4 3.2
10 96.1 95.7 0.0 0.4 0.0 0.4 2.9 0.0 0.3 0.8 0.0 0.0 0.6 2.7
11 93.0 93.4 0.0 3.3 3.5 1.4 0.0 0.9 0.0 0.0 0.0 0.0 3.5 0.9
12 97.0 98.8 0.3 0.0 1.2 0.6 0.0 0.0 0.6 0.0 0.3 0.0 0.6 0.6
13 92.7 96.9 1.1 0.0 5.3 0.4 0.0 0.0 0.0 1.3 0.2 0.0 0.7 1.3
14 95.5 94.1 0.8 1.6 0.6 2.1 2.0 1.1 0.6 0.0 0.0 0.0 0.6 1.1
15 91.7 86.5 2.9 4.5 0.0 3.6 3.2 1.8 0.9 1.8 0.3 0.0 1.1 1.8
16 96.3 93.1 0.0 0.0 0.8 0.8 1.2 0.0 0.6 3.1 0.2 0.0 1.0 3.1
17 78.5 54.9 12.9 5.5 1.1 0.0 2.2 23.1 2.2 9.9 0.0 2.2 3.2 4.4
18 83.6 53.9 3.4 15.7 0.0 0.0 9.5 15.7 0.0 5.6 0.0 0.0 3.4 9.0
19 41.3 54.3 19.8 8.7 7.4 0.0 28.1 13.0 2.5 21.7 0.0 0.0 0.8 2.2
20 28.2 63.1 48.7 21.5 0.0 0.0 17.9 0.0 0.0 10.8 0.0 0.0 5.1 4.6
사 사
본 연구는 한국해양연구원 연구과제 “남해특별관리해 역의 관리를 위한 해양 생태계 건강지수 개발(PE98582)”
와 한국해양과학기술진흥원의 “기후변화가 남해권역 해양 생태계에 미치는 영향 및 기능평가 기술개발(PM55980)”
지원으로 수행되었습니다.
참고문헌