Alterations of Amino Acid Level in Depressed Rat Brain
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치
관련 문서
Hwa Jung Kim, Bon-Cheol Ku, Jae Kwan Lee “Use of Pyrolyzed Amino Acids as Versatile Dopants for Synthesis of Heteroatoms-doped
따라서 본 연구에서는 미역과 다시마 열수추출물을 이용하여 항암 , 혈당강하 , 항산화 및 간 기능보호와 숙취해소 등이 용이한 기능성 식품 제조 기술을 개발하기 위해
Dombret H, Sutton L, Duarte M, Daniel MT, Leblond V, Castaigne S, Degos L: Combined therapy with all-trans retinoic acid and high-dose chemotherapy in
The stereoscopic microscope image of gold particles in the leaching residue from Geumsan sample as a function of nitric acid concentration on the filter papers by lead
Manipulation of protein’s amino acid sequence to change its function or properties.
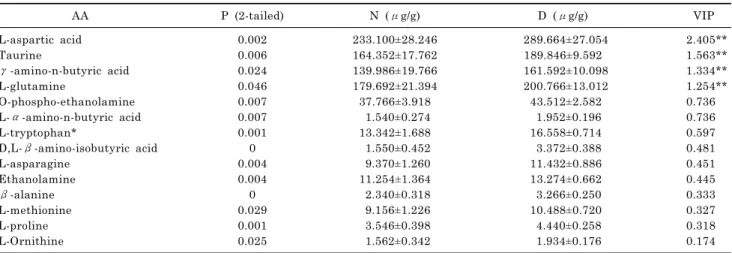
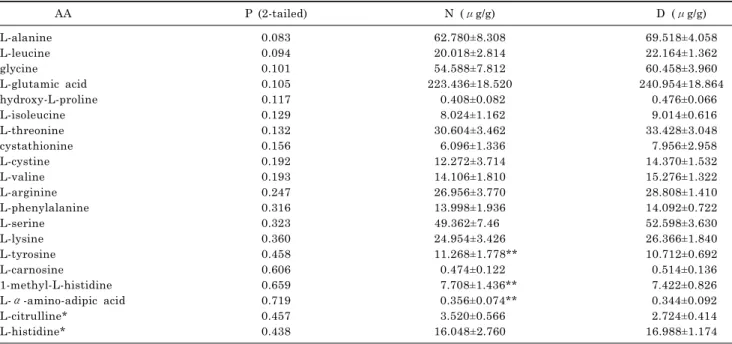
phenylalanine, carnosine, ornitine contents were significantly higher in carrot than in parsnip, while α-aminoadipic acid, leucine, tyrosine, γ-amino-n-butyric
이 스케일억 제제들로는 Acrylic acid-hydroxypropyl acrylate copolymer, Styrene sulfonic acid-maleic acid copolymer, Acrylic acid-sulfonic acid copolymer
학순도 측정을 위해서 키랄 컬럼을 이용한 액체 크로마토그래피 방법이 가장 정확하고 편리한 분석법으로 알려져 있다 [2]. 그 중에서 α-amino acid ester 의 아미노
