Vol. 11, No. 2, June, 2004
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
<접수일:2004년 3월 10일, 심사통과일:2004년 6월 1일>
※통신저자:김 완 욱
경기도 수원시 팔달구 지동 93
가톨릭대학교 의과대학 내과학교실 류마티스 내과
Tel:031) 249-8204, Fax:031) 249-7060, E-mail:[email protected] 본 연구는 보건복지부 연구비 지원에 의하여 이루어진 것임(NO 03-PJ10-20500-0021).
배양된 류마티스 활액 단핵세포 및 활막 섬유아세포에서 Vascular Endothelial Growth Factor의 자극에 의한 Tumor
Necrosis Factor-α와 Interleukin-6 생성의 증가
가톨릭대학교 의과대학 내과학교실 류마티스 내과
고혁재․유승아․우성용․김해림․조철수․김완욱
= Abstract =
Vascular Endothelial Growth Factor Stimulates Productions of Tumor Necrosis Factor-α and Interleukin-6 from Mononuclear Cells and
Synoviocytes of Rheumatoid Arthritis Patients
Hyeok-Jae Ko, M.D., Seung-Ah Yoo, M.S., Seong-Yong Woo, M.D., Hae-Rim Kim, M.D., Chul-Soo Cho, M.D., Wan-Uk Kim, M.D.
Department of Internal Medicine, Division of Rheumatology, Colloge of Medicine, The Catholic University of Korea, Seoul, Korea
Objective: Vascular endothelial growth factor (VEGF), an angiogenic factor, has been suggested to play a critical role in the pathogenesis of rheumatoid arthritis (RA). In this study, we investigated whether VEGF would directly regulate the activation of mononuclear cells of RA patients.
Methods: Mononuclear cells and/or synoviocytes of RA patients were cultured in the presence of VEGF, and the levels of TNF-α and IL-6 were determined in the culture supernatants by ELISA. The TNF-α-or IL-6-producing cells were also assessed by flow cytometry analysis. Blocking experiments were performed by adding anti-VEGF receptor (anti-Flt-1) mAb to the cells, stimulated with VEGF.
서 론
류마티스 관절염(rheumatoid arthritis, 이하 RA)은 활막세포, 대식세포, 침윤성 백혈구, 신생혈관 등으 로 이루어진 활막의 염증과 과도한 증식을 특징으로 한다1). 특히, 신생혈관형성(angiogenesis)은 RA 활막 에서 비정상적으로 증가되어 있는데, 이는 질병 초 기에 더욱 뚜렷하다2). 새로이 생성된 혈관은 활막염 부위로 염증세포를 운반하여 만성 염증상태를 유지 할 수 있게 하고 또한 pannus에 영양분과 산소를 공 급한다3,4). 신생 혈관형성은 다양한 유발물질과 억제 물질들에 의해 엄격히 조절된다. 현재까지 RA에서 다양한 신생혈관 자극인자들이 알려져 왔는데, aci- dic and basic fibroblast growth factor, transforming growth factor (TGF)-β, angiogenin, placenta growth factor (PIGF), VEGF 등이 해당되며, 이 중 VEGF가 가장 핵심적인 역할을 한다3,4).
VEGF는 dimeric glycoprotein으로 혈관 내피세포의 증식과 이동을 유도하여, 새로운 혈관을 형성하고 혈관 투과성을 증가시킨다. 흥미롭게도 최근의 연구 결과에서 VEGF는 직접적으로 nuclear factor-κB (NF- κB)를 활성화시키고, 혈관 내피세포(endothelial cell) 로부터 Interleukin-8 (IL-8)과 monocyte chemoat- tractant protein-1 (MCP-1)의 합성을 유도한다고 알려 졌다5,6). 이 보고는 VEGF가 신생 혈관 생성부위를 향해 백혈구의 충원이 이루어지도록 염증세포를 직 접 자극하거나 활성화시킬 수 있음을 시사한다. 실 제 VEGF는 RA 환자의 혈청, 활액(synovial fluid, 이
하 SF), 혹은 염증이 생긴 활막 등에서 그 표현이 증가되어 있고 이러한 점이 치료적 응용을 가능하게 한다7-9). 예를 들면, anti-VEGF antibody (Ab)를 이용 한 치료는 마우스에서 collagen-induced arthritis (CIA) 를 감소시키는 효과를 보인다10). 또한 soluble VEGF 수용체에 의해 특이적으로 VEGF의 작용을 차단할 경우 설치류에서 CIA의 위중도가 감소된다11). VEGF는 내피세포에 존재하는 Flt-1, Flk-1과 같은 수용체에 결합함으로써 활성을 나타낸다. Flk-1은 강 력한 intrinsic tyrosine kinase 활성을 가지고 있으며 VEGF에 반응하여 내피세포 증식을 유발하는 데 중 요한 연결 인자이다. 그러나 Flt-1은 Flk-1과는 다르 게 대식세포(macrophage)/단구(monocyte) 등의 염증세 포 표면에서도 표현되어 있다12,13). 최근 연구결과, VEGF와 구조적으로 아주 유사한 삼차원 구조를 가 진 PIGF로 처리한 말초 혈액 단핵구(peripheral blood mononuclear cell, 이하 PBMC)에서 MCP-1, IL-8, TNF- α, IL-1β를 포함하는 chemokine들의 생성이 증가된 다고 보고되었다14,15). 이러한 결과는 VEGF-Flt-1 복 합체가 전 염증성(pro-inflammatory) 사이토카인을 유 도하여 염증세포를 활성화시킬 수 있음을 시사하나, 아직 이에 대한 연구는 없다.
본 연구에서, 저자들은 VEGF가 직접 류마티스 염 증반응을 증폭시키는 매개체로 작용할 수 있는지를 조사하였다. 이를 위해 RA 환자로부터 단핵구를 분 리하여 VEGF로 자극한 후 배양액 상층액에서 대표 적인 염증 매개 물질인 TNF-α와 IL-6를 측정하였 다. 그 결과 VEGF는 RA 환자 혹은 정상인의 단구 (monocytes) 표면의 수용체인 Flt-1에 결합하여, TNF- Results: VEGF directly increased the productions of TNF-α and IL-6 from peripheral blood mononuclear cells (PBMC) from healthy controls. Treatment of PBMC with anti-VEGF receptor (anti-Flt-1) mAb blocked the VEGF-induced productions of TNF-α and IL-6, sug- gesting that VEGF activates the PBMC via a receptor (Flt-1) coupling event. Synovial fluid mononuclear cells (SFMC) of RA patients showed a greater response to VEGF stimulation than the PBMC of healthy controls. The major cell types responding to VEGF were mono- cytes and synoviocytes. In addition, dexamethasone completely abrogated VEGF- stimulated productions of TNF-α and IL-6 from adherent cells, isolated from SFMC.
Conclusion: Our data suggest that VEGF may directly activate RA monocytes and synoviocytes to produce TNF-α and IL-6.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: VEGF, TNF-α, IL-6, Monocytes, Rheumatoid arthritis
α와 IL-6생성을 농도 의존적으로 증가시킬 수 있음 을 확인하였다. 이러한 결과는 VEGF가 ‘신생혈관 형성’ 이라는, 이미 알려진 고유의 기능 이외에, 단 구를 자극-활성화시킴으로써 RA ‘염증반응을 유지/
증폭’시키는 데 직접적으로 관여함을 시사한다.
대상 및 방법 1. 시약(Reagents)
인형 재조합(human recombinant) VEGF165는 R &
D (Minneapolis, MN)에서 구입하였다. Anti-Flt-1 monoclonal antibody (mAb)와 isotype control mAb도 역시 R & D에서 구입하였다. Flow cytometry anal- ysis를 위한 phycoerythrin-conjugated (PE) anti-CD14 mAb, PE-conjugated anti-CD3 mAb, fluorescein isothio- cyanate (FITC)-conjugated anti-TNF-α mAb, FITC-con- jugated anti-IL-6 mAb는 PharMingen (San Diego, CA) 에서 구입하였다. ELISA를 위한 recombinant human TNF-α, IL-6, VEGF 및 이 cytokine들에 대한 차단항 체(mAb)는 R & D에서 구입하였다. Phytohaemagglutin (PHA), phorbol 12-myristate 13-acetate (PMA), ionom- ycin, lipopolysacchride (LPS), dexamethasone (Dexa)는 Sigma (St. Louis, MO)에서 구입하였다. 배양을 위해 사용한 모든 시약은 Limulus amebocyte cell lysate assay를 이용하여 endotoxin이 200 pg/mL 이하로 포 함되어 있음을 확인하였다.
2. 단핵구의 분리와 배양
15명의 건강한 정상인으로부터 동의서(informed consent)를 얻은 후, 헤파린(heparin) 처리된 말초혈액 (100 mL)을 무균적으로 채취하였고, RPMI-1640으로 1:1 비율로 희석하였다. 또한 미국 류마티스학회의 진단 기준16)에 부합하는 8명의 RA 환자로부터 관절 천자를 통하여 SF를 얻었으며, 채취된 SF는 즉시 PBS를 이용하여 1:5로 희석하고 sterile gauze를 통 과시켰다17). RA 환자는 7 명이 여자였고 1명이 남자 였으며, 평균연령은 45.6세, 평균 유병기간은 28.7개 월이었다. 8명의 환자 중 7명이 류마티스 인자가 양 성이었다. 말초혈액과 SF의 단핵구는 Ficoll-Hypaque (SG 1077)상의 비중 차에 의해 분리되었다. 분리된 단핵구를 complete medium (RPMI-1640 supplemented
with 10% fetal calf serum, 100 U/mL penicillin, 100 mg/mL streptomycin and 2 mM L-glutamine)에 넣고, 96 well plate에 1×106/well의 농도로 재부유하였다.
다양한 농도의 VEGF165 (0.1 to 10 ng/mL, R & D) 를 배양된 단핵구에 첨가하였으며, 양의 대조항원으 로 PHA 1μg/mL, PMA 50 ng/mL plus ionomycin 5μg /mL과 LPS 5μg/mL 등을 사용하였다. 단핵구에 대 한 VEGF 작용의 특이성을 조사하기 위해, VEGF를 첨가하기 1시간 전에, VEGF 수용체를 차단하는 항 체인 anti-Flt-1 mAb를 단핵구 배양액에 첨가하였다.
일부 실험에서는, 이전에 기술된 방법에 따라 SF내 단 핵구로부터 부착세포(adherent cell)만을 분리하였고18), 분리된 부착세포 혹은 비부착세포(non-adherent cell) 만을 따로 배양하여 VEGF로 자극하였다. Flow cytometry를 이용한 세포 아형 분석에서 부착세포의 70% 이상이 단구임이 확인되었다 (결과 생략). 세포 배양은 24시간 동안 시행하였으며, 배양액 상층액 내에서 TNF-α와 IL-6 등과 같은 세포활성물질의 농 도를 ELISA방법으로 측정하였다. 배양액 상층액은 측 정할 때까지 -20oC에 보관하였다.
3. 활막세포의 분리와 배양
관절 치환 수술을 받은 RA 환자로부터 활막조직 을 채취하였으며, 활막 조직 채취 전 모든 환자로부 터 동의서를 받았다. 이전에 시행한 방법과 같이19), 얻어진 조직을 잘게 자르고 collagenase로 처리한 후 원심 분리하여 얻어진 활막세포를 계대 배양하여 섬 유아 세포 양 활막세포(fibroblast-like synoviocyte, 이 하 FLS)를 분리하였다. 약술하면, 조직을 2∼3 mm 조각으로 잘게 썬 후 37oC (5% CO2 atmosphere) Dulbecco’s modified Eagle’s medium (DMEM) 안에서 type I collagenase (Worthington Biochemical, Freehold, NJ)로 4시간 동안 처리하였다. 분리된 세포를 DMEM (supplemented with 10% FCS, 2 mM glutamine, peni- cillin and streptomycin)에서 재부유시킨 후, 75 cm2 flask에 plate시켰다. 밤새 배양한 후 비부착세포를 제거하고 부착세포만을 DMEM plus 10% FCS에 배 양하였다. 배양온도는 37oC (5% CO2 atmosphere)를 유지하였고 medium은 3일마다 갈아주었다. 이 세포 를 fresh medium으로 1:3으로 희석한 후 사용될 때 까지 재배양하였다. 이후 계대 배양을 거듭하여 FLS
만 남게 되는 passage 4에서 8 사이의 세포를 각각 의 실험에 사용하였다. 이 세포들은 형태학적으로 균일하였으며, 역상 현미경 (inverse microscopy)하에 서 FLS에 특징적인 양극성(bipolar) 모양을 보였다 (결과 생략). 예비 연구에서 세포들의 균질성을 flow cytometry analysis를 이용해 확인하였다 (>95%
CD90, <2 %CD14, <1% CD3, and <1% CD19 positive, 결과 생략). FLS cell line은 다양한 농도의 VEGF와 함께 24-well plates (Nunc)에 2×104 cell/well 농도로 DMEM (supplemented with 5% FCS, 100 U/mL penicillin, 100 mg/mL streptomycin and 2 mM L-glutamine) 용액에서 배양되었다. 모든 배양은 trip- licate로 시행하였고, 24시간 배양 후 cell-free media 를 채취하였다.
4. TNF-α, IL-6, VEGF165 농도의 측정
배양액 상층액에 유리된 TNF-α와 IL-6의 양은 이 전에 기술한 바와 같이 enzyme-linked immunosorbent assay (ELISA)를 통해 측정하였다20). RA 환자의 SF (n=40) 내 TNF-α, IL-6, VEGF165의 농도 역시 ELISA 방법으로 측정하였다. 이때 재조합 인형(recombinant human) TNF-α, IL-6, VEGF165 (R & D)는 배양액으 로 희석하였고, calibration standard (5∼2000 pg/mL)로 사용하였다.
5. Flow cytometry analysis
단핵구에서 TNF-α와 IL-6의 세포표면항원 (e.g CD3, CD14)의 표현과 동시에 세포 내 사이토카인 합성정도를 flow cytometry 방법으로 측정하였다. 요 약하면, 서로 다른 농도의 VEGF (0.1∼10 ng/mL) 또 는 mitogen으로 12시간 동안 단핵구를 자극한 뒤에, 1μM monensin (GolgiStop; PharMingen)을 배양의 마 지막 2시간 동안 첨가하였다. 살아있는 세포를 채취 하여 blocking buffer (PBS with 10% normal rat serum and 0.02% 1M sodium azide)로 20분 동안 배양하였 고, 30분 동안 (4oC in the dark) fluorescein isothio- cyanate (FITC)-labeled anti-TNF-α mAb (PharMingen) 또는 FITC-labeled anti-IL-6 mAb (PharMingen)로 염 색하였다. 마지막으로 이 세포들을 2차례 세척 한 후 staining buffer에 재부유하였고, FACScan cytome- ter (Becton Dickinson, Mountain View, CA)로 염색된
정도를 분석하였다.
6. 통계분석
결과는 평균±표준편차로 표시하였다. 집단 간의 평균값 비교는 paired 또는 unpaired Mann-Whitney U-test를 이용하였다. 두 변수 사이의 상관관계는 Spearman’s rank correlation coefficient를 이용하여 분 석하였다. 결과는 p<0.05일 때 통계학적으로 유의 하다고 판정하였다.
결 과
1. 정상인 말초혈액 단핵구에서 VEGF에 의한 TNF-α와 IL-6의 생성의 증가
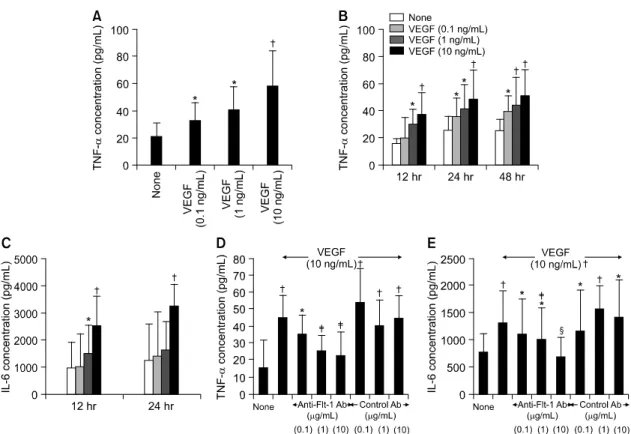
저자들은 먼저 정상인 PBMC에서 VEGF165가 염증 반응을 직접 유도할 수 있는지 조사하였다. 이를 위 해, 건강한 정상인(n=7)의 말초혈액에서 PBMC를 분리 하여 VEGF165와 함께 배양한 후, RA 병인에서 핵심 적인 염증 매개물질들인 TNF-α와 IL-6의 농도를 배 양 상층액에서 측정하였다. 그림 1A와 1B에서 보는 바와 같이, VEGF165 (0.1∼10 ng/mL)로 자극 시, 자 극한 VEGF의 농도 및 배양 시간에 비례하여 단핵 구로부터 생성되는 TNF-α의 농도가 점차 증가하였 다. PBMC로부터 생성되는 IL-6의 합성 역시, VEGF165 자극에 의해 농도 및 시간 의존적으로 증가 되었다(그림 1C). TNF-α와 IL-6의 최대 증가치는 VEGF165 10 ng/mL로 자극할 경우 자극이 없는 경우 (즉, 배양액만으로 자극할 경우)에 비교하여 각각 2.7배, 2.6배였다. VEGF에 의한 TNF-α와 IL-6생성 증가의 특이도는 anti-VEGF receptor Ab를 사용한 억 제 실험을 통해 확인하였다. PBMC에 0.1∼10 mg/
mL의 anti-Flt-1 mAb를 처리한 결과, VEGF에 의한 TNF-α와 IL-6의 생성증가가 농도 의존적으로 감소 하여 거의 기저 수준까지 저하되었다(그림 1D와 1E).
그러나 isotype control mAb는 VEGF에 의한 TNF-α 와 IL-6의 생성증가에 전혀 영향을 주지 않았다.
2. RA 환자의 활액 내 단핵구에서 VEGF에 의한 TNF-α와 IL-6의 생성의 증가
RA 환자의 활액 단핵구(synovial fluid mononuclear cell, 이하 SFMC)는 만성 염증 상태를 유지, 증폭시
킬 수 있는 다양한 전 염증성 사이토카인(e.g. TNF-α, IL-6) 환경에 노출되어 있다. 저자들은 RA 환자의 SFMC를 VEGF로 자극할 경우 정상인의 PBMC처럼 TNF-α와 IL-6의 생성이 증가될 수 있는지 조사하였 다. 그림 2A와 2B에서 보는 바와 같이, RA 환자 (n=5)의 SFMC를 VEGF로 자극 시, TNF-α와 IL-6의 생성이 농도 의존적으로 증가함을 확인하였다. RA 와 정상인 간에 TNF-α와 IL-6의 생성 정도를 비교 하였을 때, 자극이 주어지지 않을 경우(배양액만으 로 자극할 경우) RA SFMC로부터 생성되는 TNF-α 와 IL-6양은 정상인 PBMC에 비해 2배, 1.8배 높았다
(p<0.05). 단핵구를 VEGF (0.1∼10 ng/mL)로 자극할 경우 역시, RA SFMC는 정상인 PBMC와 비해 높은 농도의 TNF-α와 IL-6를 생산하였다(p<0.01). 이 경 우, SFMC에 의한 TNF-α와 IL-6의 생성 정도는 정 상인 PBMC의 최대 2.9배, 1.9배였다(그림 2A, 2B).
40명의 RA 환자에게서 분리한 SF에서 측정된 VEGF 농도는 TNF-α와 IL-6농도와 강한 순 상관 관계를 보였다(각각 r=0.513 and p<0.001, r=0.654 and p<
0.001)(그림 2C).
Fig. 1. VEGF specifically induces the productions of TNF-α and IL-6 by peripheral blood mononuclear cells (PBMC). PBMC were isolated from healthy volunteers and cultured in triplicate with various con- centrations of VEGF, ranging from 0.1 to 10 ng/mL. The levels of TNF-α and IL-6 were measured in the culture supernatants by ELISA. (A, B, C) Dose- and time-dependent increase in the production of TNF-α and IL-6 by PBMC. Data are presented as mean±SD of seven independent experiments. (D, E) The specificity of VEGF-stimulated production of TNF-α and IL-6. PBMC were cultured with VEGF 10 ng/mL in the presence or the absence of various concentrations (0.1 to 10μg/mL) of blocking antibodies (Abs) to Flt-1. The equivalent concentration of isotype Abs were used for a negative control. Each value represents the mean±SD of five separate experiments. (*: p<0.05, †: p<0.01 versus medium alone, ‡: p<0.05, §: p<0.01 versus cells treated with 10 ng/mL of VEGF.
3. VEGF 자극에 반응하는 세포의 아형분석
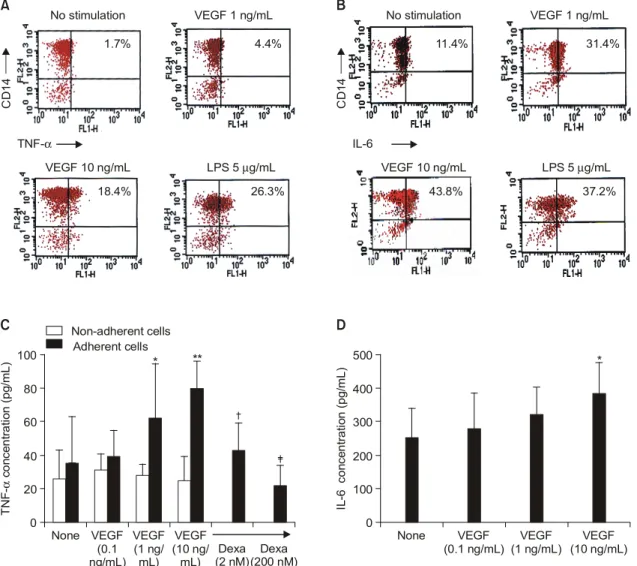
다음으로, VEGF 자극에 대해 반응하여 TNF-α와 IL-6의 생성을 증가시키는 주된 세포아형을 분석하 였다. 그림 3A에서 보듯이 건강인의 PBMC를 다양 한 농도의 VEGF로 자극했을 때, CD14 (+) 세포에 서 자극한 VEGF에 비례하여 TNF-α를 합성하는 세 포의 비율이 농도-의존적으로 증가되었다(1.7% for medium alone, 4.4% for 1 ng/mL of VEGF, 18.4% for 10 ng/mL of VEGF). CD14와 IL-6가 모두 양성인 세 포의 비율도 VEGF 자극에 의해 농도 의존적으로 증가하였다(11.3% for medium alone, 31.3% for 1
ng/mL of VEGF and 43.8% for 10 ng/mL of VEGF) (그림 3B). 그러나 CD3 (+) 세포는 VEGF로 자극 시 TNF-α와 IL-6 합성에 전혀 변화를 보이지 않았 다(결과 생략).
대식세포/단구와 섬유아세포(synovial fibroblast)는 염증이 생긴 RA 활막을 구성하는 주된 세포로서, VEGF, TNF-α, IL-6의 합성의 대부분이 이 세포들 에서 이루어진다7,8). 저자들은 마지막으로, 활액 내 단구 혹은 활막세포 역시 VEGF에 반응하여 TNF-α 와 IL-6생성을 증가시킬 수 있는지 조사하였다. 그림 3C에서 보듯이 SFMC에서 분리된 부착세포(주로 단 구)는 VEGF로 자극 시, TNF-α의 생성을 농도 의존 Fig. 2. Effect of VEGF on the productions of TNF-α and IL-6 by synovial fluid mononuclear cells (SFMC).
SFMC were isolated from 5 RA patients and then incubated with VEGF for 24 hours. The productions of TNF-α and IL-6 from SFMC were determined by ELISA in the culture supernatants, and compared with those from PBMC of healthy controls. (A, B) Dose-dependent increase in the productions of TNF-α and IL-6 by SFMC. Data are expressed as the mean±SD of five independent experiments. *: p<0.05, †: p<
0.01 versus PBMC of seven healthy controls. (C) Correlation of VEGF levels with TNF-α and IL-6 concentrations measured in synovial fluid from 40 RA patients.
Fig. 3. Monocytes and synovial fibroblasts are major cell types responding to VEGF stimulation. (A, B) Flow cytometry analysis for TNF-α and IL-6 production by PBMC of healthy controls. PBMC were stimulated with different concentrations of VEGF for 12 hours, and then stained with PE-conjugated anti-CD14 monoclonal antibody (mAb), PE-conjugated anti-CD3 mAb, FITC-conjugated anti-TNF-α mAb and FITC- conjugated anti-IL-6 mAb. LPS (5μg/mL) was used as a positive control antigen to stimulate monocytes.
A representative result from three independent experiments using different cells is shown. (C) VEGF-triggered TNF-α production by the adherent versus non-adherent cells. The adherent and non- adherent cells were separated from SFMC, and then each type of cells was cultured apart with VEGF (0.1 to 10 ng/mL) for 24 hours. Additionally, adherent cells were incubated with various concentrations of dexamethasone (Dexa) in the presence of VEGF 10 ng/mL. The TNF-α concentration was measured by ELISA in the culture supernatants. Data represent the mean±SD of three independent experiments. *: p<
0.05, **: p<0.01 versus medium alone, †: p<0.05, ‡: p<0.01 versus adherent cells stimulated with VEGF 10 ng/mL. (D) IL-6 production by fibroblast-like synoviocytes (FLS) of RA patients. FLS were stimulated with various concentrations of VEGF for 24 hours. The IL-6 concentration was measured by ELISA in the culture supernatants. Data are presented as mean±SD of three independent experiments. *:
p<0.05 versus medium alone.
TNF- concentration (pg/mL)α IL-6 concentration (pg/mL)
None None VEGF
(0.1 ng/mL) VEGF (1 ng/mL)
VEGF (10 ng/mL) VEGF
(0.1 ng/mL)
VEGF (1 ng/
mL) VEGF (10 ng/
mL) Dexa (2 nM) Dexa
(200 nM) Non-adherent cells
Adherent cells
TNF-α IL-6
CD14 CD14
* * *
0 0
100 500
40 200
80 400
60 300
20 100
A
C
B
D
No stimulation No stimulation
VEGF 10 ng/mL VEGF 10 ng/mL
VEGF 1 ng/mL VEGF 1 ng/mL
LPS 5 g/mLµ LPS 5 g/mLµ
37.2%
43.8%
31.4%
11.4%
26.3%
18.4%
4.4%
1.7%
*
적으로 증가시켰다. 이와 유사하게 FLS를 VEGF로 자극 시 IL-6의 생성이 농도 의존적으로 증가되었다 (그림 3D).
또한, 부착세포를 10 ng/mL의 VEGF로 자극 시 서 로 다른 농도의 dexamethasone (2 and 200 nM)를 동 시에 처리할 경우, TNF-α 합성이 농도 의존적으로 감소하여 거의 기저 수준까지 억제되었다(그림 3C).
그러나 비부착세포(주로 림프구)는 VEGF자극에 대 해 전혀 반응하지 않았다(결과 생략).
고 찰
TNF-α와 IL-6과 같은 전 염증성 사이토카인은 RA의 병태생리에 결정적 역할을 한다고 알려져 있
다21-26). TNF-α는 prostaglandin과 같은 염증 매개물질
뿐만 아니라 여러 다른 염증성 사이토카인과 chemo- kine의 합성을 자극한다21,22). RA 환자에서 TNF-α의 농도는 혈청과 활액에서 증가되어 있으며 질환의 증 상과 예후와 밀접한 관련이 있다23). IL-6는 염증 반 Fig. 4. Model of vicious cycle between VEGF and TNF-α production, leading to perpetuation of rheumatoid
inflammation. VEGF is produced by infiltrating synoviocytes, near endothelial cells, and acts on them via interactions with its receptors. VEGF may recruit monocytes around the endothelial cells in the synovial membrane, where newly employed macrophages, in addition to the resident synoviocytes, can produce TNF-α and IL-6 by VEGF stimulation, or through cell contact with activated endothelial cells. TNF-α and IL-6, in turn, further enhance the capacity of macrophages and synoviocytes to secrete VEGF, and also stimulate the endothelial cells to induce the cell-contact-mediated macrophage activation, generating a positive feedback-loop. In this context, VEGF also may provide the mutual activation link between the endothelial cells and macrophages/synoviocytes. If this assumption is correct, the auto-amplification loop could be broken off by treatment with Dexa, which showed a potent down-regulatory effect on the VEGF production.
monocytes
T H
TC
B
High endothelial
venule
Synoviocyte
VEGF Flt-1
Flt-1
Flk-1 TNF-α, IL-6, PGE2
TNF-α, IL-6, PGE2
Dexa Anti-Flt-1 mAb
Anti-Flk-1 mAb
Dexa Anti-Flt-1 mAb
angiogenesis monocytes
T H T H
TC TC
B
High endothelial
venule
Synoviocyte
VEGF Flt-1
Flt-1
Flk-1 TNF-α, IL-6, PGE2
TNF-α, IL-6, PGE2
Dexa Anti-Flt-1 mAb
Anti-Flk-1 mAb
Dexa Anti-Flt-1 mAb
angiogenesis
응 초기에 주로 대식세포와 단구에 의해 생성되는 또 하나의 중요한 사이토카인으로 RA에서 조직파괴 를 유발하는 데 핵심적인 역할을 한다24). 또한 IL-6 는 B 세포의 성장과 활성화를 증진시키고 immuno- globulin의 합성을 강력히 유도한다25,26). 이들 두 사 이토카인들은 모두 단핵구 혹은 활막세포를 자극하 여 VEGF 생성을 유도할 수 있다3,4).
본 연구에서 VEGF는 인체 단핵구로부터 TNF-α 와 IL-6의 합성을 직접적으로 증가시켰으며 이러한 증가는 anti-Flt-1 mAb의 처리에 의해 거의 완전히 차단되었다. RA 환자의 SFMC는 정상인 PBMC에 비해 VEGF 자극에 대해 보다 강력한 반응을 보였 다. 단구와 FLS는 VEGF에 반응하여 TNF-α와 IL-6 를 생산하는 주된 세포였다. 본 연구에서 in vitro 실 험에 사용된 VEGF165는 40명의 RA 환자의 활액에서 측정된 VEGF165 농도인 0.1∼10 ng/mL의 범위 내에 있으므로, 상술한 결과는 환자 체내에서 일어날 수 있는 생리학적인 반응이라고 생각된다(그림 2C). 종 합하여 볼 때, 저자들의 결과는 RA 환자의 관절 내 에 과도하게 형성된 VEGF가 수용체인 Flt-1에 결합 하여 단구와 활막세포를 자극/활성화시키고, TNF-α 와 IL- 6와 같은 전 염증성 사이토카인의 생성을 증 가시킴으로써 만성염증 반응을 유지/증폭시키는 데 중요한 역할을 하고 있음을 시사한다.
RA 병인의 최근 모델은 사이토카인 분비와 세포 간 접촉을 통한 면역 세포 간의 복잡한 상호작용이 강조되고 있다. VEGF는 혈관 내피세포(endothelial cell)근처의 활막세포에 의해 합성되고 내피세포 수 용체에 결합할 경우 내피세포는 증식하거나 활성화 된다7,8). 최근 보고에 의하면, VEGF는 내피세포를 활성화시켜 MCP-1과 IL-8 같은 chemokine의 합성을 유도한다고 한다5,6). 이와 관련하여, 저자들의 연구는 기존의 RA 병인 모델에 부가적인 새로운 기전을 제 공한다. 먼저, 높은 농도의 VEGF는 증식된 내피세 포 주위로 단구를 모여들게 할 수 있고, 새로이 모 여든 단구/대식세포와 활막세포는 VEGF의 자극 또 는 활성화된 내피세포와 접촉을 통해 TNF-α와 IL-6 의 합성을 증가시킬 수 있다. 새로이 합성된 TNF-α 와 IL-6는 대식세포와 활막세포를 자극하여 VEGF를 분비할 수 있는 능력을 증가시키고, 증가된 VEGF는 내피세포를 또 다시 활성화시켜 염증반응의 악순환
고리(positive feedback-loop)가 형성된다(그림 4). 이와 유사한 방법으로 VEGF는 내피세포와 대식세포/활막 세포 사이의 상호 활성화 연결고리(mutual activation link)를 제공할 수 있다. 만일, 이러한 가정이 옳다 면, Dexa와 같은 약물은 VEGF의 자극에 의한 TNF-α 와 IL-6의 생성을 감소시킴으로써 상술한 염증반응 의 악순환의 고리가 절단하고, 이를 통해 관절염 치 료효과가 나타날 수도 있을 것이다(그림 4).
결 론
본 연구에서, 저자들은 신생혈관 자극인자 중 하 나인 VEGF가 류마티스 염증반응을 증폭시키는 매 개체로 직접 작용할 수 있는지를 조사하였다. RA 환자 단핵구와 활막세포를 배양하여 VEGF로 자극 한 결과 자극한 VEGF의 농도에 비례하여 배양 상 층액내 TNF-α와 IL-6생성이 증가됨을 발견하였다.
이러한 결과는 RA 환자의 관절 내에 과도하게 형성 된 VEGF가 신생혈관형성이라는 이미 알려진 기능 이외에도, 단구와 활막세포를 활성화시키고 전 염증 성 사이토카인의 생성을 촉진시킴으로써 관절 내 만 성염증 반응을 유지, 증폭시키는 데 중요한 역할을 하고 있음을 시사한다.
REFERENCES
1) Koch A. Angiogenesis: implications for rheumatoid arthritis. Arthritis Rheum 1998;41:951-62.
2) FitzGerald O, Soden M, Yanni G, Robinson R, Bresnihan B. Morphometric analysis of blood ves- sels in synovial membranes obtained from clini- cally affected and unaffected knee joints of pa- tients with rheumatoid arthritis. Ann Rheum Dis 1991;50:792-6.
3) Klagsbrun M, D'Amore PA. Regulators of angio- genesis. Annu Rev Physiol 1991;53:217-39.
4) Dvorak HF, Brown LF, Detmar M, Dvorak AM.
Vascular permeability factor/vascular endothelial growth factor, microvascular hypermeability and angiogenesis. Am J Pathol 1995;146:1029-39.
5) Marumo T, Schini-Kerth VB, Busse R. Vascular endothelial growth factor activates nuclear factor- κB and induces monocyte chemoattractant protein- 1 in bovine retinal endothelial cells. Diabetes 1999;
48:1131-7.
6) Lee TH, Avraham H, Lee SH, Avraham S. Vas- cular endothelial growth factor modulates neut- rophil transendothelial migration via up-regulation of interleukin-8 in human brain microvascular endothelial cells. J Biol Chem 2002;277:10445-51.
7) Fava RA, Olsen NJ, Spencer-Green G, Yeo KT, Yeo TK, Berse B, et al. Vascular permeability factor/endothelial growth factor (VPF/VEGF): accu- mulation and expression in human synovial fluids and rheumatoid synovial tissue. J Exp Med 1994;
180:341-6.
8) Nagashima M, Yoshino S, Ishiwata T, Asano G.
Role of vascular endothelial growth factor in angio- genesis of rheumatoid arthritis. J Rheumatol 1995;
22:1624-30.
9) Lee SS, Joo YS, Kim WU, Min DJ, Min JK, Park SH, et al. Vascular endothelial growth factor levels in the serum and synovial fluid of patients with rheumatoid arthritis. Clin Exp Rheumatol 2001;19:321-4.
10) Sone H, Kawakami Y, Sakauchi M, Nakamura Y, Takahashi H, Shimano H, et al. Neutralization of vascular endothelial growth factor prevents collagen- induced arthritis and ameliorates established disease in mice. Biochem Biophys Res Commun 2001;
281:562-8.
11) Miotla J, Maciewicz R, Kendrew J, Feldmann M, Paleolog E. Treatment with soluble VEGF receptor reduces disease severity in murine collagen-induced arthritis. Lab Invest 2002;80:1195-205.
12) Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z.
Vascular endothelial growth factor (VEGF) and its receptors. FASEB J 1999;13:9-22.
13) Autiero M, Luttun A, Tjwa M, Carmeliet P. Pla- cental growth factor and its receptor, vascular endothelial growth factor receptor-1: novel targets for stimulation of ischemic tissue revascularization and inhibition of angiogenic and inflammatory dis- orders. J Thromb Haemost 2003;1:1356-70.
14) Selvaraj SK, Giri RK, Perelman N, Johnson C, Malik R, Kalra VK. Mechanism of monocyte acti- vation and expression of proinflammatory cytochemo- kines by placenta growth factor. Blood 2003;102:
1515-24.
15) Iyer S, Leonidas DD, Swaminathan GJ, Maglione D, Battisti M, Tucci M, et al. The crystal struc- ture of human placenta growth factor-1 (PlGF-1), an angiogenic protein, at 2.0 A resolution. J Biol
Chem 2001;276:12153-61.
16) Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum 1988;31:315-24.
17) Park SH, Min DJ, Cho ML, Kim WU, Youn J, Park W, et al. Shift toward T helper 1 cytokines by type II collagen-reactive T cells in patients with rheumatoid arthritis. Arthritis Rheum 2001;44:
561-9.
18) Kalra VK, Ying Y, Deemer K, Natarajan R, Na- dler JL, Coates TD. Mechanism of cigarette smoke condensate induced adhesion of human monocytes to cultured endothelial cells. J Cell Physiol 1994;
160:154-62.
19) Cho CS, Cho ML, Min SY, Kim WU, Min DJ, Lee SS, et al. CD40 engagement on synovial fibroblast up-regulates production of vascular endo- thelial growth factor. J Immunol 2000;164:5055-61.
20) Cho ML, Kim WU, Min SY, Min DJ, Min JK, Lee SH, et al. Cyclosporine differentially regulates interleukin-10, interleukin-15, and tumor necrosis factor-a production by rheumatoid synoviocytes.
Arthritis Rheum 2002;46:42-51.
21) Brennan FM, Maini RN, Feldmann M. TNF α--a pivotal role in rheumatoid arthritis? Br J Rheu- matol 1992;31:293-8.
22) Dayer JM, Beutler B, Cerami A. Cachectin/tumor necrosis factor stimulates collagenase and pros- taglandin E2 production by human synovial cells and dermal fibroblasts. J Exp Med 1985;162:
2163-88.
23) Tetta C, Camussi G, Modena V, Di Vittorio C, Baglioni C. Tumour necrosis factor in serum and synovial fluid of patients with active and severe rheumatoid arthritis. Ann Rheum Dis 1990;49:665-7.
24) Guerne PA, Zuraw BL, Vaughan JH, Carson DA, Lotz M. Synovium as a source of interleukin 6 in vitro. Contribution to local and systemic manife- stations. J Clin Invest 1989;83:585-92.
25) Ito A, Itoh Y, Sasaguri Y, Morimatsu M, Mori Y.
Effects of interleukin-6 on the metabolism of connective tissue components in rheumatoid syno- vial fibroblasts. Arthritis Rheum 1992;35:1197-201.
26) Le JM, Vilcek J. Interleukin 6: a multifunctional cytokine regulating immune reactions and the acute phase protein response. Lab Invest 1989;61:588-602.