HepG2 세포에서 Isoeugenol의 PCSK9 억제 효능
최효경․정민유
한국식품연구원 식품기능연구본부
Isoeugenol Inhibits PCSK9 in HepG2 Cells
Hyo-Kyoung Choi and Min-Yu Chung
Research Division of Food Functionality, Korea Food Research Institute
ABSTRACT Hypercholesterolemia is a major cause of a number of different chronic diseases, including atherosclerosis.
Statins for the treatment of hypercholesterolemia induce transactivation of low-density lipoprotein receptor (LDLR) and proprotein convertase subtilisin/kexin type 9 (PCSK9). PCSK9 interferes with LDL cholesterol and its receptor binding, promoting LDLR degradation and thereby increasing circulating LDL cholesterol. In the current study, HepG2 cells were treated with 10% delipidated serum (DLPS) in the presence or absence of isoeugenol. Lipid depletion in- creased both LDLR and PCSK9 protein expression levels in HepG2 cells, and isoeugenol inhibited PCSK9 in HepG2 cells. The objective of this study was to investigate the mechanism by which isoeugenol inhibits PCSK9 and subsequently increases LDLR protein expression HepG2 cells under lipid depletion conditions. A non-toxic level of isoeugenol reduced PCSK9 protein expression without affecting LDLR protein expression. Lipid depletion enhanced gene ex- pression of LDLR, PCSK9, and their transcription factors SREBP2 and HNF1α in HepG2 cells. Isoeugenol significantly attenuated LDLR, PCSK9, and SREBP2 gene expression (P<0.0001), but not HNF1α. Consistently, isoeugenol also markedly reduced SREBP2 protein expression, but not HNF1α. This suggests that isoeugenol-mediated SREBP2 down- regulation reduced LDLR and PCSK9 gene expression, and attenuated PCSK9 expression contributed to maintenance of LDLR protein expression in HepG2 cells under lipid depletion conditions. In DLPS-treated HepG2 cells, isoeugenol dose-dependently reduced statin-mediated elevation of PCSK9 protein expression. This study suggests isoeugenol as a potential statin co-treatment material and PCSK9 inhibitor.
Key words: isoeugenol, PCSK9, LDL receptor, SREBP2, HepG2
Received 15 May 2020; Accepted 10 June 2020
Corresponding author: Min-Yu Chung, Korea Food Research Insti- tute, Wanju, Jeonbuk 55365, Korea
E-mail: [email protected], Phone: +82-63-219-9371
서 론
고콜레스테롤 혈증은 동맥경화 및 심장병의 주요 위험요 인이다(Morsy와 Fouad, 2008). 다량의 저밀도 지단백 콜레 스테롤(low-density lipoprotein cholesterol; LDL cho- lesterol)은 동맥 내 축적되어 죽종을 형성하고, 혈관세포에 독성을 주어 동맥경화, 고혈압, 비만, 당뇨뿐만 아니라 간, 심장, 신장 기능의 저하를 일으킨다(Jain 등, 2010). 따라서 혈중 LDL 콜레스테롤의 감소는 심장병 및 관련 질환의 발병 률을 낮출 수 있다(Ichihashi 등, 1998; Shi 등, 2000). 동맥경 화 및 심장병 고위험군 환자들에게 가장 오랫동안 그리고 널 리 처방되어온 약물은 스타틴이다(Shahreyar 등, 2018). 스 타틴은 3-hydroxy-3-methyl-glutaryl-CoA(HMG-CoA) 환원효소의 억제제로써 LDL 콜레스테롤의 생합성을 막아 혈중 LDL 콜레스테롤 수치를 낮추는 약물이다(Shahreyar
등, 2018). 가족성 고콜레스테롤혈증(familial hypercho- lesterolemia) 환자나, 스타틴 약물을 오랫동안 복용하여 스 타틴 저항성을 가진 환자들에게 고농도의 스타틴이나 다른 콜레스테롤 저감 약물을 함께 복용해도 혈중 LDL 저감 효능 이 나타나지 않는 경우가 많다(Shahreyar 등, 2018). 스타틴 복용 후 LDL 콜레스테롤 수치가 낮아진 동맥경화 환자 5명 중 1명은 5년 이내에 두 번째 동맥경화 증상이 나타난다는 연구 결과 또한 보고되었다(Alkhalil, 2019). 이는 모두 LDL 수용체(LDL receptor; LDLR)의 결함과 관련이 있으며, 스 타틴 약물의 한계점으로도 지적되어 왔다.
Proprotein convertase subtilisin/kexin type 9a(PCSK9) 는 전사 후 변형을 통하여 LDLR을 조절하고, 결과적으로 혈 중 콜레스테롤 함량에 영향을 미친다(Maxwell과 Breslow, 2004). PCSK9 유전자의 기능상실돌연변이(loss-of-func- tion mutation)는 세포 표면의 LDLR을 증가시키고 LDLR 의 세포내 이동을 감소시켰으며, 기능획득돌연변이(gain- of-function mutation)는 세포표면의 LDLR 단백질의 발현 을 감소시켰다는 연구 결과가 이를 부분적으로 뒷받침 한다 (Berge 등, 2006; Naoumova 등, 2005; Cameron 등, 2006).
Rashid 등(2005)은 PCSK9 knockout 마우스의 간에서 LDLR 단백질 발현이 증가하였고, 이로 인한 혈중 LDL 콜레 스테롤이 감소하였음을 보고하였다. 스타틴은 SREBP2 (sterol regulatory element-binding protein-2) 전사인자 의 발현을 증가시킴으로써 LDLR과 PCSK9 유전자의 활성 을 증가시킨다. PCSK9(-/-) 마우스에 투여된 스타틴은 간 조직 내 LDLR 단백질 발현을 증가시켜 혈중 LDL 콜레스테 롤을 감소시킨다. 즉, PCSK9은 간 조직에서 LDLR 단백질 발현을 억제하며, PCSK9 억제제는 스타틴과 동시 투여 시 LDLR의 증진 및 나아가 혈중 LDL 콜레스테롤을 감소시킨 다(Rashid 등, 2005). 고콜레스테롤 혈증 환자 중 스타틴 약물을 복용하고 있으나 LDL 콜레스테롤 저감 효능이 나타 나지 않은 환자를 대상으로 한 임상실험 결과, PCSK9 억제 제인 alirocumab(Robinson 등, 2015)와 evlocumab(Sa- batine 등, 2015)는 LDL 콜레스테롤 저감 효과를 60% 이상 증가시켰다. 이후 Lopez(2008)와 Seidah(2009)에 의해 PCSK9은 고콜레스테롤 혈증 치료에 있어서 스타틴의 한계 를 극복할 수 있는 새로운 표적단백질로 제안되어 왔다.
PCSK9 억제제인 alirocumab와 evolocumab는 2015년 미 국 FDA의 승인을 받은 물질로서, LDL 콜레스테롤 저감 효 능이 스타틴을 단독 복용할 때보다 뛰어나다(Shahreyar 등, 2018).
PCSK9 억제제는 스타틴 저항성 혹은 스타틴 단독 복용 으로 LDL 콜레스테롤 수치가 잘 감소하지 않는 환자들에게 스타틴을 대신할 수 있는 약물이다(Shahreyar 등, 2018).
PCSK9을 억제하는 천연 소재는 alirocumab, evolocumab 등의 PCSK9 억제제가 가지는 여러 가지 단점들인 정신적 문제, 집중력 저하, 경제적 부담 등을 보완하면서 스타틴 복 용과 함께 혹은 독립적으로 고콜레스테롤 혈증 혹은 콜레스 테롤 저항성을 개선할 수 있는 안전한 소재로 제안될 수 있다.
Eugenol(4-allyl-1-hydroxy-2-methoxybenzene)은 정향, 시나몬, 바질, 넛메그 등의 식물로부터 추출한 에센셜 오일에 함유된 안전성이 입증된(GRAS; generally recog- nized as safe) 페놀 성분으로써, 항산화(Nam과 Kim, 2013), 항박테리아(Devi 등, 2010), 항염(Fonseca 등, 2016), 뇌 질환 예방(Prasad와 Muralidhara, 2012) 등의 다양한 생리 활성을 지니고 있다. Eugenol의 이성체인 isoeugenol(2- methoxy-4-propenyl-phenol) 또한 여러 향신료에 함유 되어 방향제 및 장기저장제로 쓰이며, eugenol과 유사한 생 리활성을 지닌다고 보고되어 왔다(Atsumi 등, 2005; Bor- tolomeazzi 등, 2010; Melles 등, 2013). Eugenol과 iso- eugenol은 o-methoxy phenol계 화합물로서(Hidalgo와 De la Rosa, 2009; Choi 등, 2007), 생리활성은 유사하지만 각각 다른 메커니즘을 통해서 그 활성을 나타내기도 한다.
현재까지 동맥경화 유발 식이를 먹인 랫 모델에서 eugenol 의 항동맥경화 및 항산화 효능이 입증된 바 있고(Venka- deswaran 등, 2014), eugenol의 LDL 저감 활성 또한 고콜 레스테롤혈증 랫(Triton WR 1339-induced hypercholes-
terolemic rat) 모델에서 입증되었다(Venkadeswaran 등, 2014). 하지만 현재까지 isoeugenol의 기능성에 대한 연구 는 이루어지지 않았다. 본 연구는 eugenol의 이성체인 iso- eugenol의 PCSK9 억제에 관여하는 전사인자 측정을 통해 기전을 밝히고, 스타틴과 동시 처리 시 isoeugenol의 PCSK9 억제 정도를 측정함으로써 스타틴의 한계를 보완할 수 있는 소재로 제안될 수 있는 가능성을 제시하고 있다.
재료 및 방법
HepG2 세포 배양
HepG2 세포는 ATCC(HB-8065; Manassas, VA, USA) 에서 구매하였다. HepG2 세포는 10% fetal bovine serum (FBS; Welgene Inc., Gyeongsan, Korea)과 1% antibi- otics/antimycotics(Welgene Inc.)를 첨가한 배지(DMEM- high glucose; Corning, Manassas, VA, USA)를 사용하여 37°C, 5% CO2 조건에서 배양하였다.
Western blot
HepG2 세포가 약 70% confluence에 도달했을 때, 6- well plate에 세포를 분주(5×104/wells)하였다(day 0). 24 시간 후 세포를 cold Dulbecco’s phosphate-buffered sal- ine(DPBS; Welgene Inc.)으로 2회 세척하고 fresh DMEM 배지로 교체하였다. 이때 음성 대조군에는 DMEM+10%
FBS를, 양성 대조군에는 DMEM에 지방이 제거된 FBS(de- lipidated serum, DLPS, 10%)와 1% antibiotics/anti- mycotics(DMEM+10% DLPS)를 첨가하여 24시간 동안 배양하였다(day 1). DLPS는 선행논문을 바탕으로 지방(지 방산, 콜레스테롤, 중성지방)이 제거된 상태로 제조되었다 (Hannah 등, 2001). 이후 배지를 제거하고 cold DPBS로 2회 세척한 후, fresh DMEM+10% FBS 혹은 DMEM+
10% DLPS로 각각 교환하였다. DMEM+10% DLPS에 배양 한 세포에만 isoeugenol을 50, 100 µM로 처리하였다(day 2). 18시간 배양 후(day 3), cold DPBS로 2회 세척하고 세 포를 회수하였다. 원심분리(1,200 rpm, 3분)하여 얻은 pel- let을 cell lysis buffer(Cell Signaling Technology, Dan- vers, MA, USA)로 lysis하고, Bradford assay법으로 단백 질을 측정하였다(Bio-Rad Laboratories, Hercules, CA, USA). 총 15 μg의 단백질을 8% SDS gel에 전기영동하고, 항체[PCSK9(Circulex, Medical & Biological Laborato- ries Co., Ltd., Woburn, MA, USA), LDLR(BioVision Inc., Milpitas, CA, USA), hepatic nuclear factor 1α(HNF1α;
Cell Signaling Technology), SREBP2(Cayman Chemical, Ann Arbor, MI, USA), β-actin(Bethyl Laboratories, Montgomery, TX, USA)]를 이용하여 western blot을 하 였다. ECL solution(Pierce Biotechnology, Rockford, IL, USA)에 배양한 후, 표적 단백질의 밴드를 관찰하였다(Bio- Rad Laboratories).
MTT assay
HepG2 세포가 약 70% confluence에 도달했을 때, tryp- sin-EDTA(Welgene Inc.)를 이용하여 세포를 떼어내어 96-well plate에 분주(5×104/wells)하였다(day 0). 24시 간 후, 세포를 cold DPBS(Welgene Inc.)로 2회 세척하고 fresh DMEM 배지로 교체하였다. 이때 음성 대조군에는 DMEM+10% FBS를, 양성 대조군에는 DMEM+10% DLPS 를 첨가하여 24시간 동안 배양하였다(day 1). 배지를 제거 하고 cold DPBS로 2회 세척한 후, fresh DMEM+10% FBS 혹은 DMEM+10% DLPS로 각각 교환하였다. DMEM+10
% DLPS에 배양한 세포에만 isoeugenol을 농도별(12.5, 25, 50, 100 μM)로 처리하였다(day 2). 18시간 배양 후(day 3), cold DPBS로 2회 세척하고 FBS가 포함되지 않은 배지 로 교체한 뒤 3-(4,5-dimethylthiazol-2-yl)-2,5-diphe- nyltetrazolium bromide(MTT, Sigma-Aldrich, St. Louis, MO, USA) solution을 배지에 첨가하여 두 시간 배양하였 다. 형성된 blue formazan을 DMSO(Sigma-Aldrich)에 용 해하여 570 nm와 630 nm에서 세포 생존률을 측정하였다 (microplate reader; Molecular Devices, San Jose, CA, USA).
Total RNA의 추출 및 qRT-PCR
HepG2 세포가 약 70% confluence에 도달했을 때, 6- well plate에 세포를 배양(5×104/wells)하였다(day 0). 24 시간 후, 세포를 cold DPBS로 2회 세척하고 fresh DMEM 배지로 교체하였다. 이때 음성 대조군에는 DMEM+10%
FBS를, 양성 대조군에는 DMEM+10% DLPS를 첨가하여 24시간 동안 배양하였다(day 1). 배지를 제거하고 cold DPBS로 2회 세척한 후, fresh DMEM+10% FBS 혹은 DMEM+10% DLPS로 각각 교환하였다. DMEM+10%
DLPS에 배양한 세포에만 isoeugenol 12.5, 25, 50, 100 μM을 처리하였다(day 2). 18시간 배양 후(day 3), cold DPBS로 2회 세척하였다. 회수한 세포 pellet에서 total RNA 를 추출하고 cDNA를 합성하였다. qRT-PCR 분석은 SYBR Green PCR Master Mix reagent(Thermo Fisher Scientific, Waltham, MA, USA)를 이용하여 다음과 같은 조건으로 시 행하였다(Bio-Rad CFX Connect Real-Time PCR Detec- tion System, Bio-Rad Laboratories). 모든 결과는 GAPDH 에 따라 상대적으로 정량화되었다. PCR은 95°C에서 30초, 56°C에서 30초, 72°C에서 30초, 40 cycle로 시행되었으 며, 마지막 72°C에서 10분간 추가로 반응하여 안정화시켰 다. 증폭의 모든 데이터는 매 cycle의 72°C, 30초 단계에서 수집되었다. 본 분석에 사용된 프라이머는 다음과 같다.
PCSK9은 Forward 5′-GGCAGGTTGGCAGCTGTTT- 3′, Reverse 5′-CGTGTAGGCCCCGAGTGT-3′, LDLR은 Forward 5′-AGGAGACGTGCTTGTCTGTC-3′, Reverse 5′-CTGAGCCGTTGTCGCAGT-3′, SREBP2는 Forward 5′-CGGTAATGATCACGCCAACAT-3′, Reverse 5′-TG
GTATATCAAAGGCTGCTGGAT-3′, HNF1α는 Forward 5′-TGGCGCAGCAGTTCACCCAT-3′, Reverse 5′-TGA AACGGTTCCTCCGCCCC-3′, 그리고 GAPDH는 Forward 5′-ATGTTCGTCATGGGTGTGAAC-3′, Reverse 5′-GC ATGGACTGTGGTCATGAGT-3′가 사용되었다. 모든 반 응은 3회 반복으로 시행되었으며, 상대적 발현 및 SD value 는 comparative method를 사용하여 계산하였다.
스타틴 처리
HepG2 세포가 약 70% confluence에 도달했을 때, 6- well plate에 세포를 분주(5×104/wells)하였다(day 0). 24 시간 후, 세포를 cold DPBS로 2회 세척하고 fresh DMEM+
10% DLPS를 첨가하여 24시간 동안 배양하였다(day 1). 배 지를 제거하고 cold DPBS로 2회 세척한 후, fresh DMEM +10% DLPS 처리 혹은 DMEM+10% DLPS+스타틴[ro- suvastatin 0.2 μM+mevalonate 50 μM]을 처리하였다. 스 타틴 처리군에 isoeugenol을 0, 12.5, 25, 50, 100 μM을 동시 처리하였다(day 2). 18시간 배양 후(day 3), cold DPBS로 2회 세척하고 세포를 회수하였다. 원심분리(1,200 rpm, 3분) 후 세포 pellet을 얻었다. 위와 동일한 방법으로 western blot을 시행하였다.
통계 분석
본 연구에서는 최소 3회 반복에 대한 평균 및 표준편차로 표시했으며, 각 실험군을 대조군에 대한 백분율로 나타내었 다. 각 실험군 간의 통계적 유의성은 Student’s t-test를 사 용하여 검정하였다. 통계검정은 GraphPad Prism(version 5.0, GraphPad Software, San Diego, CA, USA)을 이용하 였다.
결과 및 고찰
HepG2 세포에서 isoeugenol의 LDLR, PCSK9 단백질 발 현 조절
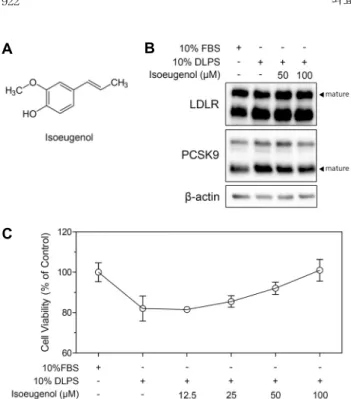
Isoeugenol(Fig. 1A)을 DLPS 첨가로 지방이 제거된 환 경의 HepG2 세포에 처리한 후, total lysates에서 LDLR과 PCSK9 단백질 발현을 western blot으로 측정하였다. 그 결과, isoeugenol(50, 100 μM)은 HepG2 세포에서 DLPS 처리에 의해 증가한 LDLR 단백질 발현에 영향을 주지 않았 다(Fig. 1B). 한편, HepG2 세포에서 DLPS 처리는 PCSK9 단백질 발현을 현저히 증가시켰고, isoeugenol(50, 100 μM) 은 PCSK9의 단백질 발현을 농도 의존적으로 감소시켰다 (Fig. 1B). HepG2 세포에 DLPS를 처리하여 지방 제거 환경 으로 만들고, isoeugenol을 100 μM 농도까지 처리하여 앞 서 관찰된 PCSK9의 단백질 발현 감소가 isoeugenol에 의 한 세포 생존 억제에 의한 것인지 측정하기 위해 MTT assay 를 수행하였다. 그 결과, DLPS 처리 시 isoeugenol은 100 μM 농도까지 세포 생존률에 영향을 주지 않았다(Fig. 1C).
A B
C
Fig. 1. Chemical structure of isoeugenol (A). LDLR, PCSK9, and β-actin proteins expression regulated by isoeugenol (B).
Cellular viability of HepG2 cells treated with DLPS and/or iso- eugenol (12.5∼100 μM) (C).
A B
C D Fig. 2. Gene expression levels of LDLR
(A), PCSK9 (B), SREBP2 (C), and HNF1α (D) in HepG2 cells treated with DLPS or isoeugenol (12.5∼100 μM). All data are expressed as mean±SD from the samples of each group. # indicates a significant differ- ence compared to FBS (#P<0.05, ###P<0.001).
* indicates a significant difference compared to DLPS (*P<0.05, **P<0.01, ***P<0.001).
지방제거 환경에서 세포는 항상성 유지를 위해 SREBP2 의 활성을 통하여 LDLR과 PCSK9의 전사를 증가시킨다.
본 연구에서는 isoeugenol의 PCSK9 억제 정도를 관찰하기 위해 세포를 지방 제거 상태로 만들어 PCSK9의 발현을 임 의적으로 증가시켰으며, Fig. 1B가 이러한 결과를 뒷받침하 고 있다. 따라서 지방 제거 환경에 의해 이미 증가한 LDLR
단백질 발현이 isoeugenol 처리에 의해 추가적으로 증가하 지는 않았음을 관찰하였다. 특히, isoeugenol은 PCSK9 억 제제로써 지방 제거 환경에서 LDLR을 degradation 시키지 않고 단백질 발현량을 유지하도록 작용을 한 것에 의의가 있겠다.
HepG2 세포에서 isoeugenol의 LDLR, PCSK9, SREBP2, HNF1α유전자 발현 조절
PCSK9과 LDLR 및 주요 조절 전사인자인 SREBP2, HNF1α의 유전자 발현 조절을 qRT-PCR로 측정하였다. 지 방이 제거된 상태에서 HepG2 세포의 PCSK9, LDLR 그리 고 전사인자인 SREBP2, HNF1α의 유전자 발현량은 모두 대조군 대비 유의적으로 높았다(P<0.01; Fig. 2A-D). Iso- eugenol(12.5~100 μM) 처리는 농도 의존적으로 LDLR, PCSK9, SREBP2의 유전자 발현량을 감소시켰으며, 이는 통계적으로 유의하였다(P<0.001; Fig. 2A-C). 하지만 지방 제거에 의해 증가한 HNF1α는 isoeugenol에 의해 유전자 발현이 조절되지 않았다(Fig. 2D).
PCSK9과 LDLR은 프로모터에 sterol regulatory ele- ments(SREs)를 가지고 있으므로 SREBP pathway의 활성 조절을 통해 세포 내 콜레스테롤 항상성을 유지시킨다. 콜레 스테롤 고갈 상태에서 세포는 SREBP2 전사인자의 활성 증 진을 통해 HMGCR, LDLR, PCSK9 등의 유전자 발현을 증 가시켜 세포 내 콜레스테롤을 증진시킨다(Horton, 2002).
SREBP 외에 PCSK9의 전사 활성을 독립적으로 조절하는 전사인자인 HNF1α에 대한 연구가 최근 활발히 이루어지고 있다. PCSK9은 프로모터에 HNF1α binding element를 가 지고 있어서 HNF1α가 PCSK9의 전사 조절에 관여함이
Fig. 3. Protein expression of HNF1α, SREBP2, and β-actin regulated by isoeugenol at 50 and 100 μM in HepG2 cells.
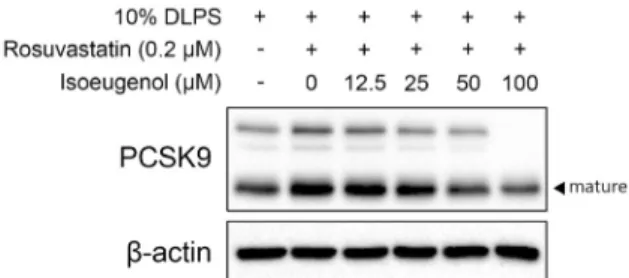
Fig. 4. PCSK9 protein expression in HepG2 cells with DLPS in the absence or presence of rosuvastatin (0.2 µM of rosuvasta- tin and 50 µM of mevalonate) and isoeugenol (0, 12.5, 25, 50, 100 µM).
HepG2 세포에서 밝혀진 바 있다(Li 등, 2009). PCSK9의 천연 억제제로 연구되어 온 berberine은 HNF1α 활성 억제 를 통해 혈중 LDLR clearance를 최종 증가시켰으며(Li 등, 2009), curcumin의 PCSK9 전사 억제능 또한 SREBP2가 아닌 HNF1α를 통해 이루어짐이 발표된 바 있다(Tai 등, 2014). 본 연구에서는 isoeugenol에 의한 PCSK9과 LDLR 의 전사 억제는 HNF1α가 아닌 SREBP2를 통하여 이루어짐 을 알 수 있었다.
HepG2 세포에서 isoeugenol의 HNF1α, SREBP2 단백질 발현 조절
Isoeugenol(50, 100 μM)을 HepG2 세포에 처리하여 PCSK9, LDLR 발현 조절에 관여하는 주요 전사인자의 단백 질 발현을 western blot으로 측정하였다. HepG2 세포에서 지방 제거에 의해 증가한 HNF1α 단백질 발현은 isoeugenol 처리(50, 100 μM)에 의해 농도 의존적으로 증가하는 경향 을 보였다(Fig. 3). 반면, SREBP2는 isoeugenol 처리(50, 100 μM)에 의해 단백질 발현이 현저하게 감소하였다(Fig.
3). 지방이 고갈된 상태의 HepG2 세포에서 isoeugenol의 PCSK9 조절은 SREBP2를 통해서 이루어짐을 알 수 있었다.
PCSK9은 세포 밖으로 배출된 이후에 세포 표면에서 LDLR의 epidermal growth factor-like receptor A 도메 인에 결합하여 LDLR의 post transcritiponal degradation 을 유도하는데, LDLR의 세포 표면으로 recycle 하는 것을 막고 LDLR의 lysosome으로 유도하여 degradation을 일으 키도록 한다(Zhang 등, 2007). Isoeugenol에 의해서 억제 된 PCSK9의 발현은 LDLR 단백질의 degradation을 막았 음을 유추할 수 있다.
HepG2 세포에서 isoeugenol과 스타틴 동시 처리 시 PCSK9, LDLR의 발현 조절
지방이 제거된 HepG2 세포에서 스타틴(rosuvastatin 0.2 μM+mevalonate 50 μM) 처리는 PCSK9 단백질의 발현을 증진시켰다(Fig. 4). 스타틴 처리에 의해 증가한 PCSK9 발 현은 isoeugenol(12.5~100 μM)에 의해 농도 의존적으로 감소하였다(Fig. 4).
스타틴은 HMG CoA 환원효소의 활성을 억제하는 약물로 써 SREBP2를 통해 LDLR의 전사를 증진시키며, 이때 SREBP2에 의해 함께 전사되는 PCSK9가 LDLR의 degra- dation을 촉진시키게 된다(Shahreyar 등, 2018; Alkhalil, 2019). 따라서 스타틴 약물의 오랜 복용이 스타틴 저항성이 나 LDL 콜레스테롤의 저감 효과가 떨어지는 등의 한계를 보 인 바 있다(Shahreyar 등, 2018; Alkhalil, 2019). 본 연구 에서 스타틴과 isoeugenol의 동시 처리는 스타틴에 의해 증 가한 PCSK9의 발현을 억제했으며, 이를 통해 isoeugenol 은 스타틴 약물의 한계를 보완시킬 수 있는 소재로서의 가능 성을 제시하고 있다.
요 약
본 논문에서는 isoeugenol의 PCSK9 억제능 관련 메커니즘 을 규명하였다. HepG2 세포를 지방 부족 환경으로 만들고 isoeugenol을 처리하여 PCSK9과 LDLR, 그리고 전사인자 의 단백질 및 유전자 발현 조절을 측정하였다. 독성 없는 농도의 isoeugenol(~100 μM)은 LDLR의 단백질 발현 조절 에 관여하지 않았지만, PCSK9의 단백질 및 유전자 발현을 감소시켰다. Isoeugenol은 PCSK9 조절에 독립적으로 관여 하는 전사인자인 HNF1α의 유전자 및 단백질 조절에는 관여 하지 않았다. 따라서 isoeugenol은 SREBP2를 통해 PCSK9과 LDLR의 유전자 발현을 억제하였음을 알 수 있었 다. 또한 DLPS로 지방 고갈이 유도된 HepG2 세포에 스타틴 (rosuvastatin 0.2 μM)과 isoeugenol을 동시 처리했을 때, isoeugenol은 스타틴에 의해 증가한 PCSK9의 발현을 농도 의존적으로 감소시켰다. 고콜레스테롤혈증 동물 모델에서 도 isoeugenol의 이러한 PCSK9 억제 효능이 관찰되는지에 대한 추후 연구가 필요하다. 본 연구를 통하여 isoeugenol 은 PCSK9 억제 활성을 나타내며 스타틴 치료와 병행하여 그 한계점을 보완할 수 있는 소재로서 제안될 수 있다.
감사의 글
본 연구는 한국식품연구원 사업연구비(과제번호 E0150301
-06)의 지원을 받아 수행한 연구 결과로 이에 감사드립니 다.
REFERENCES
Alkhalil M. Proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitors, reality or dream in managing patients with cardio- vascular disease. Curr Drug Metab. 2019. 20:72-82.
Atsumi T, Fujisawa S, Tonosaki K. A comparative study of the antioxidant/prooxidant activities of eugenol and isoeugenol with various concentrations and oxidation conditions. Toxicol In Vitro. 2005. 19:1025-1033.
Berge KE, Ose L, Leren TP. Missense mutations in the PCSK9 gene are associated with hypocholesterolemia and possibly increased response to statin therapy. Arterioscler Thromb Vasc Biol. 2006. 26:1094-1100.
Bortolomeazzi R, Verardo G, Liessi A, Callea A. Formation of dehydrodiisoeugenol and dehydrodieugenol from the reaction of isoeugenol and eugenol with DPPH radical and their role in the radical scavenging activity. Food Chem. 2010. 118:256- 265.
Cameron J, Holla ØL, Ranheim T, Kulseth MA, Berge KE, Leren TP. Effect of mutations in the PCSK9 gene on the cell surface LDL receptors. Hum Mol Genet. 2006. 15:1551-1558.
Choi YC, Park KR, Lee JH, Jeon YJ, Liu KH, Oh S, et al.
Isoeugenol suppression of inducible nitric oxide synthase ex- pression is mediated by down-regulation of NF-κB, ERK1/2, and p38 kinase. Eur J Pharmacol. 2007. 576:151-159.
Devi KP, Nisha SA, Sakthivel R, Pandian SK. Eugenol (an es- sential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J Ethnopharmacol. 2010. 130:107-115.
Fonsêca DV, Salgado PR, Aragão Neto HC, Golzio AMFO, Caldas Filho MRD, Melo CGF, et al. Ortho-eugenol exhibits anti-nociceptive and anti-inflammatory activities. Int Immu- nopharmacol. 2016. 38:402-408.
Hannah VC, Ou J, Luong A, Goldstein JL, Brown MS. Unsatu- rated fatty acids down-regulate SREBP isoforms 1a and 1c by two mechanisms in HEK-293 cells. J Biol Chem. 2001.
276:4365-4372.
Hidalgo ME, De la Rosa C. Antioxidant capacity of eugenol derivatives. Quim Nova. 2009. 32:1467-1470.
Horton JD. Sterol regulatory element-binding proteins: tran- scriptional activators of lipid synthesis. Biochem Soc Trans.
2002. 30:1091-1095.
Ichihashi T, Izawa M, Miyata K, Mizui T, Hirano K, Takagishi Y. Mechanism of hypocholesterolemic action of S-8921 in rats: S-8921 inhibits ileal bile acid absorption. J Pharmacol Exp Ther. 1998. 284:43-50.
Jain KS, Kulkarni RR, Jain DP. Current drug targets for anti- hyperlipidemic therapy. Mini-Rev Med Chem. 2010. 10:232- 262.
Li H, Dong B, Park SW, Lee HS, Chen W, Liu J. Hepatocyte nuclear factor 1alpha plays a critical role in PCSK9 gene tran- scription and regulation by the natural hypocholesterolemic compound berberine. J Biol. 2009. 284:28885-28895.
Lopez D. Inhibition of PCSK9 as a novel strategy for the treat- ment of hypercholesterolemia. Drug News Perspect. 2008. 21:
323-330.
Maxwell KN, Breslow JL. Adenoviral-mediated expression of
Pcsk9 in mice results in a low-density lipoprotein receptor knockout phenotype. Proc Natl Acad Sci U S A. 2004. 101:
7100-7105.
Melles D, Vielhaber T, Baumann A, Zazzeroni R, Karst U. In chemico evaluation of skin metabolism: Investigation of euge- nol and isoeugenol by electrochemistry coupled to liquid chro- matography and mass spectrometry. J Chromatogr B. 2013.
913-914:106-112.
Morsy MA, Fouad AA. Mechanisms of gastroprotective effect of eugenol in indomethacin-induced ulcer in rats. Phytother Res. 2008. 22:1361-1366.
Nam H, Kim MM. Eugenol with antioxidant activity inhibits MMP-9 related to metastasis in human fibrosarcoma cells.
Food Chem Toxicol. 2013. 55:106-112.
Naoumova RP, Tosi I, Patel D, Neuwirth C, Horswell SD, Marais AD, et al. Severe hypercholesterolemia in four British fami- lies with the D374Y mutation in the PCSK9 gene: long-term follow-up and treatment response. Arterioscler Thromb Vasc Biol. 2005. 25:2654-2660.
Prasad SN, Muralidhara. Neuroprotective efficacy of eugenol and isoeugenol in acrylamide-induced neuropathy in rats: be- havioral and biochemical evidence. Neurochem Res. 2012. 38:
330-345.
Rashid S, Curtis DE, Garuti R, Anderson NN, Bashmakov Y, Ho YK, et al. Decreased plasma cholesterol and hypersensi- tivity to statins in mice lacking Pcsk9. Proc Natl Acad Sci U S A. 2005. 102:5374-5379.
Robinson JG, Farnier M, Krempf M, Bergeron J, Luc G, Averna M, et al. Efficacy and safety of alirocumab in reducing lipids and cardiovascular events. N Engl J Med. 2015. 372:1489- 1499.
Sabatine MS, Giugliano RP, Wiviott SD, Raal FJ, Blom DJ, Robinson J, et al. Efficacy and safety of evolocumab in re- ducing lipids and cardiovascular events. N Engl J Med. 2015.
372:1500-1509.
Seidah NG. PCSK9 as a therapeutic target of dyslipidemia.
Expert Opin Ther Targets. 2009. 13:19-28.
Shahreyar M, Salem SA, Nayyar M, George LK, Garg N, Koshy SKG. Hyperlipidemia: Management with proprotein conver- tase subtilisin/kexin type 9 (PCSK9) Inhibitors. J Am Board Fam Med. 2018. 31:628-634.
Shi W, Haberland ME, Jien ML, Shih DM, Lusis AJ. Endothe- lial responses to oxidized lipoproteins determine genetic sus- ceptibility to atherosclerosis in mice. Circulation. 2000. 102:
75-81.
Tai MH, Chen PK, Chen PY, Wu MJ, Ho CT, Yen JH. Curcu- min enhances cell-surface LDLR level and promotes LDL up- take through downregulation of PCSK9 gene expression in HepG2 cells. Mol Nutr Food Res. 2014. 58:2133-2145.
Venkadeswaran K, Muralidharan AR, Annadurai T, Ruban VV, Sundararajan M, Anandhi R, et al. Antihypercholesterolemic and antioxidative potential of an extract of the plant, piper betle, and its active constituent, eugenol, in triton WR-1339- induced hypercholesterolemia in experimental rats. Evid Based Complement Alternat Med. 2014. Article ID 478973.
https://doi.org/10.1155/2014/478973
Zhang DW, Lagace TA, Garuti R, Zhao Z, McDonald M, Horton JD, et al. Binding of proprotein convertase subtilisin/kexin type 9 to epidermal growth factor-like repeat A of low density lipoprotein receptor decreases receptor recycling and increases degradation. J Biol Chem. 2007. 282:18602-18612.