91
1)
본 연구는 한국과학재단 목적기초연구(R01-2000-00150) 및 2001년도 충남대학교병원 공모과제연구비의 지원으로 수행되었음.
접수 : 2003년 3월 15일, 수정 : 2003년 5월 21일, 승인 : 2003년 5월 25일 교신저자 : 조덕연, 충남대학교 의과대학 내과학교실
Tel : 042)220-7162, Fax : 042)257-5753, E-mail : [email protected]
Interferon-gamma, Tumor Necrosis Factor-α및
Transforming Growth Factor-β
1에 의한 골수세포의 유착분자 발현 및 Stromal Cell-Derived Factor-1 생성의 조절
충남대학교 의과대학 내과학교실1, 이화여자대학교 의과대학 내과학교실2
조덕연
1・황진희
1・정효균
1・박수진
1・박상은
1・곽승근
1・윤환중
1・성주명
2・김삼용
1Regulation of Adhesion Molecule Expression and Stromal Cell-Derived Factor-1 Production in Human Bone Marrow Cells
by Interferon-gamma, Tumor Necrosis Factor-α, and Transforming Growth Factor-β1 : Implications in Bone Marrow
Homing of Hematopoietic Cells
Deog Yeon Jo, M.D.1, Jin Hee Hwang1, Hyo Kyun Chung1, Sang Eun Park, M.D.1, Soo Jin Park, M.D.1, Seung Keon Kwak, M.D.1, Hwan Jung Yun, M.D.1,
Chu Myoung Seong, M.D.2 and Samyong Kim, M.D.1
1Department of Internal Medicine, Chungnam National University College of Medicine, Daejon
2Department of Internal Medicine, Ewha Womans University College of Medicine, Seoul, Korea
Background : It is well known that harmonious interactions among adhesion molecules and stromal cell-derived factor-1 (SDF-1)-mediated chemoattraction signalling via CXCR4 are needed for bone marrow homing of hematopoietic stem cells and progenitor cells. The aim of this study was to define the role of interferon-gamma (IFN-γ), tumor necrosis factor-alpha (TNF-α), and transforming growth factor-beta 1 (TFG-β1), known as hematopoiesis-inhibitory cytokines, in the regulation of the molecules in relation to the homing.
Methods : We investigated the effects of these cytokines on the expression of CXCR4 and adhesion molecules and the production of SDF-1 in bone marrow cells including CD34+ cells, bone marrow endothelial cells (BMEC-1 cells), and bone marrow stromal cells (BMSCs). We also examined whether the cytokines influence in vitro transmigration of hematopoietic pro- genitors.
Results : None of the cytokines influenced CXCR4 expression on CD34+ cells or SDF-1- mediated chemotaxis of the cells. IFN-γ and TNF-α, but not TGF-β up-regulated the ex- pression of L-selectin, ICAM-1, and VLA-4 on CD34+ cells. However, the up-regulation was not translated into the enhanced transendothelial migration. IFN-γ and TNF-α up-regulated the expression of VCAM-1 and ICAM-1 on BMEC-1 cells, and rendered the endothelium more suitable for transendothelial migration of hematopoietic progenitors. IFN-γ and TNF-α also up-regulated the expression of VCAM-1 and ICAM-1 on primary human BMSCs. All three cytokines significantly attenuated SDF-1 production from primary BMSCs, and TNF-α dimini- shed SDF-1 production from BMEC-1 cells.
Conclusion : These data indicate that IFN-γ, TNF-α, and TGF-β1 play a role in the
서 론
조혈모세포는 빠르고 효율적으로 골수로 귀소(homing)한 다.1) 이 과정에서 조혈세포와 혈관내피세포 또는 골수기질 세포의 유착분자(adhesion molecule)의 유기적인 상호작 용이 필요하다.2) 또한 골수기질세포에서 항시 생성되고 분 비되는 케모카인(chemokine)인 stromal cell-derived factor-1 (SDF-1)이 수용체 CXCR4를 통한 화학주성 (chemotaxis)을 유발함으로써 조혈세포의 골수귀소에 결 정적인 역할을 한다.3,4) 이와 같이 골수귀소의 기전은 어느 정도 밝혀지고 있으나, 그 기전이 어떻게 조절되는지에 대 해서는 알려진 바가 많지 않다. 골수귀소 조절 기전의 규명 은 곧 조혈모세포이식 및 조혈모세포의 말초가동화의 최적 화를 위한 기초 자료로 활용될 수 있을 것이다. 그동안 조 혈성장인자가 조혈세포의 근원적 특성인 골수귀소의 조절 에 관여할 수 있을 것이라는 개연성을 확인하려는 연구가 상당 수 있었으나 일치된 결론에 이르지는 못했다. 즉, Peled 등3)은 조혈세포를 체외에서 단기간 조혈성장인자로 처리함으로써 골수 생착이 향상됨을 보고한 반면, 오히려 이러한 조작이 골수 생착을 저해한다는 보고5,6)가 있는 등 상반된 견해가 있다. 본 연구진은 최근 조혈세포에 대한 단 기간의 조혈성장인자 처리가 세포유착분자와 CXCR4 발현 및 SDF-1에 의한 경내피세포 화학주성(transendothelial chemotaxis)에 영향을 주지 않음을 규명하여 보고한 바 있 다.7)
Interferon-gamma (IFN-γ), tumor necrosis factor- alpha (TNF-α) 및 transforming growth factor-beta1
(TGF-β1)는 조혈억제 인자로 분류되는 싸이토카인이다.
이들 싸이토카인은 백혈구와 혈관내피세포 유착분자의 발 현8) 및 특정 세포에서 일부 케모카인9∼11)과 그 수용체12∼
15)의 조절에 관여한다. 조혈성장인자가 조혈세포의 골수귀 소 미치는 영향에 대해서는 어느 정도 알려져 있으나, 조혈 억제인자가 골수귀소의 조절에 관여하는지에 대해서는 잘 알려져 있지 않다. 본 연구에서는 IFN-γ, TNF-α 및 TGF-β1 등의 싸이토카인이 조혈세포의 골수귀소에 관여 하는 세포 즉, 조혈세포, 골수혈관내피세포 및 골수기질세 포의 귀소에 관여하는 분자 즉, 유착분자, CXCR4와 SDF-1의 발현 및 생성에 어떠한 영향을 미치는지를 규명
하고 골수귀소와 관련된 의미를 고찰하고자 하였다.
재료 및 방법 1. 세포 및 세포주
골수세포는 동종골수이식 시 동의를 구한 후 정상 공여 자로부터 얻었다. Ficoll을 사용하여 단핵세포를 분리한 후 MiniMACS (Miltenyi Biotec, Auburn, CA, USA)를 이 용해 지침에 따라 CD34 양성세포를 정제하였으며, FITC 가 부착된 항CD34 단클론항체(HPCA-2; Becton-Dikin- son, San Diego, CA, USA)를 반응시킨 후 유세포분석을 통해 순도를 확인하였다. 순도가 90% 이상인 세포만을 실 험에 사용하였다.
사람 골수기질세포는 Verfaillie 등16)이 제시한 방법에 따라 얻었다. 즉, 골수 단핵구로부터 CD34 양성세포를 분리하고 남은 CD34 음성세포를 MEM-α (GIBCO-BRL Life Technologies, Grand Island, NY, USA)에, 12.5%의 말 혈청(GIBCO-BRL), 12.5%의 우태혈청(GIBCO-BRL), 10-6M의 hydrocortisone (Sigma, St. Louis, MO, USA) 및 5×10-5 M의 2-mercaptoethalnol (Sigma)를 첨가한 장 기배양배지를 사용하여 배양하였다. 부착세포가 밀집 상태에 도달한 후 수확하고, 장기배양배지를 사용하여 주 1회 계대배 양하였다. 3회 계대배양한 후 CD45 양성세포는 거의 발견되 지 않았다.
골수기질세포주로는 생쥐 골수기질세포주인 MS-517)를 사용하였다. 10% 우태혈청(GIBCO-BRL)를 함유한 alpha-MEM (GIBCO-BRL)을 배지로 하여 주 1회 계대 배양하였다. MS-5는 사람의 미성숙 조혈세포의 장기 배양 을 지지하며 SDF-1을 분비한다.6)
골수내피세포로는 사람 골수 내피세포에 ‘SV large T 항 원’ 유전자를 이입한 세포주인 BMEC-118)을 사용하였다.
세포는 20% 우태혈청을 함유하는 M-199 (GIBCO- BRL) 배지를 사용하여 주 1회 계대배양하였다. 경내피세포 이동 을 위해서는 구경이 5µm인 미세공막(TranswellTM; Corn- ing-Costar, Cambridge, MA, USA)에 내피세포 단층을 형성시켜 사용하였다. 이 때 6혈 배양판(Corning-Costar) 의 경우 4×105 세포를 넣고 3일이 경과한 후 단층의 형성 regulation of bone marrow homing of hematopoietic cells via up-regulation of adhesion molecule expression and down-modulation of SDF-1 production in bone marrow cells. (Korean J Hematol 2003;38:91~99)
Key Words : Stromal cell-derived factor-1, CXCR4, adhesion molecules, interferon-gamma, tumor necrosis factor-α, transforming growth factor-β1
여부를 Quick staining kit (HEMA3TM, Fisher Scientific, Swedesboro, NJ, USA)를 사용하여 염색한 후 현미경으 로 확인한 후 사용하였다.
제대혈관내피세포는 만기 정상분만을 위한 제왕절개 시 환자 및 가족의 동의하에 무균적으로 제대를 얻어 37℃에 서 10분간 0.1% collagenase를 처리하여 얻었다. 획득한 내피세포는 50µg/mL endothelial cell growth supple- ments (ECGS; Biomedical Technologies, Inc. Stou- ghton, MA, USA), 5U/mL heparin (Sigma) 및 20%
FBS 등을 함유한 Medium-199 (GIBCO-BRL)을 사용하 여 배양하였다. 실험에는 2∼3회 계대배양한 세포를 사용 하였다. CXCR4를 발현하는 백혈병세포주인 MO7e는 10
%의 FBS와 10ng/mL의 granuocyte, macrophage colony-stimulating factor (GM-CSF; Lucky, Daejon, Korea)를 함유하는 IMDM을 사용하여 배양하였다.
2. 각종 세포의 싸이토카인 처리
골수 CD34 양성세포, BMEC-1 및 골수기질세포를 무 혈청배지(X-VIVO, BioWhittacker, Walkerville, MA, USA)에 배양하면서 IFN-γ, TNF-α 및 TGF-β1을 각각 10ng/mL, 100 U/mL 및 10ng/mL의 농도로 첨가하여 배 양한 후 분석하였다. 일부 실험에서는 interleukin-1β (IL-1β; 10ng/mL), IL-3 (50ng/mL), granulocyte col- ony-stimulating factor (G-CSF; 100ng/mL) 및 stem cell factor (SCF; 20ng/mL)을 첨가하였다. G-CSF (Kirin, Tokyo, Japan)를 제외한 모든 싸이토카인은 R&D sys- tems사(Minneapolis, MN, USA) 제품을 사용하였다.
3. 유세포분석
CXCR4 발현의 측정을 위해서는 5×104∼1×105세포를 phycoerythrin (PE)이 부착된 항CXCR4 단클론성항체 (12G5; BD PharMingen, San Diego, CA, USA)를 첨가 하여 4℃에서 30분간 반응시킨 후 Coulter Elite flow cytometer (Coulter Electronics Ltd., Hialeah, FL, USA)를 사용하여 분석하였다. 세포유착분자 CD62L (L- selectin), CD106 (VACM-1), CD39d (VLA-4), CD31 (PECAM-1), CD11a (LFA-1), CD54 (ICAM-1) 등에 대한 PE가 부착된 단클론항체(BD PharMingen)를 이용하 여 유세포분석을 시행하였다. Isotype 항체를 대조군으로 사용하였다.
4. 골수기질세포주(MS-5)를 향한 경내피세포 화학주성
Jo 등4)이 기술한 방법에 따라 시행하였다. 약술하면, 6 혈 배양판에 MS-5 단층을 형성시킨 후 배지를 완전히 교 환하고 3일간 37℃, 5% CO2, 완전 습윤 상태에서 배양하 였다. BMEC-1 단층이 형성된, 또는 BMEC-1 단층 없는 5µm의 미세공 막의 6혈 TranswellTM의 상방에 분석하고 자 하는 세포를 넣고(혈당 0.5∼1×104 CD34 양성세포), MS-5 단층이 형성된 6혈 배양판에 옮긴 후 37℃, 5%
CO2, 완전 습윤 상태에 두었다. 24시간 후 상방을 제거하 고 이동한 세포를 함유하고 있는 하방은 그대로 장기배양 을 시행하였다. 5주 후 부유세포 및 부착세포를 수확하여 집락형성세포의 함량을 계측하여 장기배양기시세포(long- term culture-initiating cell, LTC-IC)의 함량을 산출하였 다. 이 때 TranswellTM의 상방에 넣은 세포와 동일한 수의 세포를 별도의 MS-5단층이 형성된 6혈 배양판에 넣고 장 기배양을 병행하여 분석하고자 하는 세포의 cobblestone area-forming cell (CAFC)19)과 LTC-IC의 함량을 측정함 으로써 하방으로 이동한 세포의 백분율을 산출하였다. 배 양 중 1주일에 1회씩 부유 세포를 포함한 상층액의 절반을 제거하고 새로운 배지로 교환하였다. 장기배양기시세포의 함량을 측정하고 이동한 세포의 백분율을 산출하였다. 집 락형성세포분석(colony-forming cell assay)은 30% 우태 혈청, 5×10-5M의 2-mercaptoethalnol 및 5mM의 hemin (Sigma)를 함유하는 1.2% methylcellulose (Sigma)를 35 mm Petri dish (Becton-Dikinson)에 1mL의 양으로 SCF (20ng/mL), erythropoietin (EPO; 6U/mL) 및 IL-3 (50 ng/mL) 등의 조혈성장인자 첨가하에 분석하고자 하는 세 포를 넣어 37℃ 세포배양기에서 2주간 배양한 후 각각의 집 락을 표준적 기준에 따라 도립현미경하에서 세었다.
5. ELISA 분석
세포 배양 상층액의 SDF-1α의 농도를 ELISA kit (R&D Systems)를 사용하여 측정하였다. 검체를 제조사 가 제시한 지침에 따라 처리한 후 spectrophotometer (Molecular Device Co., Sunnyvale, CA, USA)를 이용 하여 optical density를 얻고, Softmax프로그램 (Molecular Device Co.)을 이용하여 표준곡선과 SDF-1치를 얻었다.
6. Northern blot
통상적인 방법에 따라 골수기질세포로부터 RNA를 추출 하였다. 20µg의 RNA를 2.4% formaldehyde를 함유한
1.2% agarose겔에서 전기영동을 시행한 후 나일론막 (Amersham Life Science, Arlington Heights, IL, USA) 에 옮겼다. SDF-1 probe로는 역전사중합효소연쇄반응을 통해 얻은 산물을 random priming kit (Amersham Life Science)를 사용하여 라벨링한 것을 사용하였다. Probe를 나일론막과 24시간 반응시켜 hybridization을 시행하고 세 척한 후 필름을 감광시켜 사진을 얻었다.
7. 통계처리
통계량은 Student-t test로 검정하였으며, P값이 0.05 미만인 경우 통계적으로 의미가 있다고 판정하였다.
결 과
1. 조혈세포의 CXCR4 및 유착분자의 발현 및 경내피세포 이동
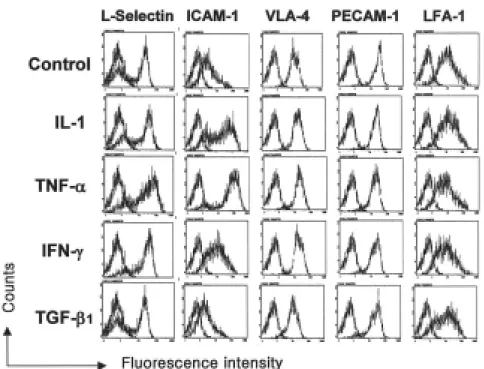
골수 CD34 양성세포를 IFN-γ (100U/mL), TNF-α (10ng/mL) 및 TFG-β (10ng/mL) 등의 싸이토카인으로 무혈청배지에서 24시간 처리한 후 관찰한 CXCR4 발현은 변화가 없었다(Fig. 1). 이들 싸이토카인의 농도를 10배 높게 하거나 낮게 한 경우와 처리 시간을 48시간까지 연장 한 경우에도 CXCR4 발현의 변화는 관찰되지 않았다(결과 제시되지 않음). 골수 CD34 양성세포는 L-selectin, VLA-4, PECAM-1 및 LFA-1을 모두 강하게 발현하였으 며, ICAM-1은 중등도로 발현하였다. TNF-α 및 IFN-γ 는 ICAM-1의 발현을 강력하게 상향 조절하였으며, L- selectin 및 VLA-4의 발현은 경미하게 상향 조절하였다.
TGF-β는 어떠한 유착분자의 발현에도 영향을 미치지 않 았다(Fig. 2).
골수 CD34 양성세포를 무혈청배지에 배양하며 IFN-γ (100U/mL), TNF-α (10ng/mL), 및 TFG-β1 (10ng/
mL)으로 24시간 처리한 후 5주차 CAFC의 MS-5를 향한 경내피세포 이동을 확인하였다. 싸이토카인의 첨가 없이 24시간 배양된 세포의 경우 30±7%의 5주차 CAFC이 하
Fig. 2. Expression of adhesion molecules on CD34+ cells and its modulation by cytokines. Bone marrow CD34+ cells were incubated in serum-free medium for 24 hours with or without a cytokine (10ng/mL), and then analyzed using flow cytometry.
Fig. 1. CXCR4 expression on bone marrow CD34+ cells.
The cells were incubated in serum-free medium with or without a cytokine (10ng/mL) for 24 hours, and analyzed using flow cytometry. None of the three cytokines influenced the expression of CXCR4.
방으로 이동하였으며, 싸이토카인의 처리는 이러한 이동에 영향을 주지 않았다(Fig. 3).
2. 골수내피세포의 CXCR4 발현
유세포분석에서 BMEC-1 세포와 일차성 사람 골수혈관 내피세포 모두에서 CXCR4 발현은 관찰되지 않았다(Fig.
4). 무혈청배지에서 BMEC-1세포를 IFN-γ (100U/
mL), TNF-α (10ng/mL) 및 TFG-β1 (10ng/mL)으로 40시간까지 처리한 경우에도 그 발현이 유도되지 않았다.
3. 골수내피세포의 유착분자 발현 및 조혈세포 투과성의 조절
BMEC-1 세포에서 ICAM-1은 약하게 발현되었으며, TNF-α 처리 후 뚜렷하게 증가하였다. PECAM-1은 강하 게 발현되었으며 싸이토카인 처리 후 변동은 없었다(Fig.
4). 이러한 결과는 싸이토카인의 농도를 10배 높게 또는 10배 낮게 하거나, 작용시간을 짧게 또는 48시간까지 연장 하여도 마찬가지였다(결과 제시되지 않음).
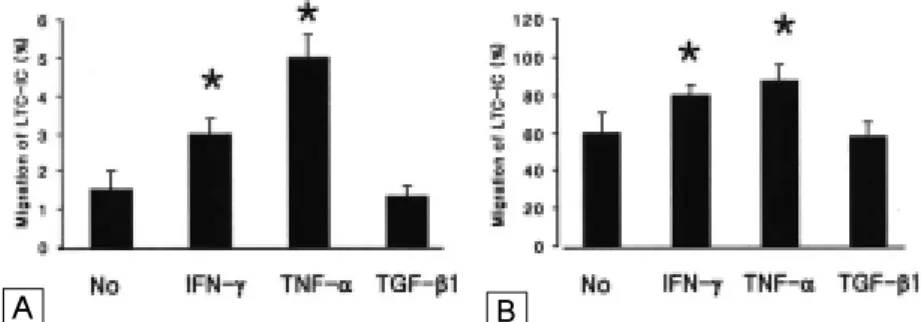
조혈억제인자가 골수내피세포의 조혈세포 투과성에 미 치는 영향을 알아보기 위하여 TranswellTM의 상방에 이미 형성된 BMEC-1세포 단층을 24시간 동안 싸이토카인으로 처리한 후 하방에 무혈청 배지만을 첨가하여 장기배양기시 세포의 24시간 동안 하방으로의 이동 즉, 자발적 이동과 하 방의 MS-5를 향한 이동을 측정하였다. 자발적 이동의 경 우 1.5±0.5%의 장기배양기시세포가 이동하였으며, BMEC-1 단층을 IFN-γ (100U/mL) 또는 TNF-α (10 ng/mL)의 처리한 경우에는 각각 2.9±0.3% 및 4.8±
0.6%로 증가하였다(P<0.05; Fig. 5A). MS-5를 향한 이 동에 있어 싸이토카인을 처리하지 않은 대조군의 장기배양
Fig. 5. Twenty four hour-migration of bone marrow CD34+ cell-derived LTC-ICs through cytokine-treated BMEC-1 monolayer. The BMEC-1 monolayers were treated with IFN-γ, TNF-α, or TGF-β1 (10ng/mL each) for 24 hours prior to the transmigration assay. (A) Spontaneous migration. (B) Migration toward the lower chamber containing MS-5 monolayer and its conditioned medium. *P<0.05 compared wtih the migration through non-cytokine-treated BMEC-1 monolayer.
Fig. 4. Expression of adhesion molecules on BMEC-1 cells.
The cells did not express CXCR4. ICAM-1 expression was negligible in basal status, but it was up-regulated by incu- bation of the cells with TNF-α (10ng/mL), but not with IFN-γ or TGF-β1, in serum-free medium for 24 hours.
BMEC-1 cells strongly expressed PECAM-1, which was not modulated by any cytokines.
Fig. 3. Transendothelial migration of cytokin-treated CD34+
cells toward MS-5. Bone marrow CD34+ cells were incubat- ed with or without a cytokine (10ng/mL) in serum-free me- dium for 24 hours. And then, the cells were loaded into the upper chamber of TranswellTM containing BMEC-1 monolayer, and allowed to migrate toward the lower chamber containing MS-5 monolayer. After 24 hours, the upper chamber was removed, and the lower chamber directly underwent long-term culture for 5 weeks. Data are presented as the mean +S.D.
of the percent migration of LTC-ICs from triplicate experi- ments.
A B
기시세포는 60±5%가 하방으로 이동하였으며, BMEC-1 단층의 IFN-γ (100U/mL) 또는 TNF-α (10ng/mL)의 처리로 각각 79±3% 및 82±4%로 증가하였다(P<0.05;
Fig. 5B).
4. 골수기질세포의 유착분자 발현의 조절
골수기질세포는 VCAM-1과 ICAM-1을 약하게 발현하 였으며 PECAM-1은 발현하지 않았다. 기질세포를 무혈청 배지에서 24시간 동안 IFN-γ (100U/mL), TNF-α (10 ng/mL) 및 TFG-β1 (10ng/mL)으로 각각 처리한 후 시 행한 유세포분석에서 VACM-1의 발현은 TNF-α에 의해 뚜렷하게 증가하였으며, ICAM-1의 발현은 IFN-γ 및 TNF-α 모두에 의해 뚜렷하게 증가하였다. TGF-β1은 유 착분자의 발현에 아무런 영향이 없었다(Fig. 6). 이러한 결과는 싸이토카인의 농도 또는 작용시간에 상관없이 동일 하였다(결과 제시되지 않음).
5. 골수내피세포 및 골수기질세포의 SDF-1 생성 조절
싸이토카인이 골수혈관내피세포 및 골수기질세포의 SDF-1 생성 또는 분비의 조절에 관여하는지를 확인하기 위하여 이들 세포를 무혈청배지에 72시간 동안 IFN-γ (100U/mL), TNF-α (10ng/mL) 및 TFG-β1 (10ng/
mL)을 각각 첨가하여 배양하고, 이로부터 얻은 상층액의 SDF-1의 농도를 측정하였다. 3일간 배양한 BMEC-1 상층
액의 SDF-1 농도는 1,710±205pg/mL 이었다. 이러한 SDF-1의 생성은 IL-1β와 SCF의 첨가로 각각 6,152±
611pg/mL와 4,068±368pg/mL로 증가하였으며, TNF-α 의 첨가로 278±81pg/mL로 현저하게 감소하였다(Table 1). 골수기질세포의 경우 대조군의 25.2±3.9ng/mL에 비 해 IFN-γ, TNF-α와 TGF-β1 처리로 SDF-1의 농도는 각각 12.8±0.9ng/mL, 10.9±1.8ng/mL 및 9.4±4.1 ng/mL로 뚜렷하게 감소하였다(Table 1). 단, BMEC-1 세포와는 달리 IL-1β 또는 SCF 등 조혈성장인자에 의한 Fig. 6. Expression of adhesion molecules on primary bone marrow
stromal cells. The cells were incubated in serum-free medium with or without cytokine (10ng/mL) for 24 hours, and analyzed using flow cytometry.
Table 1. SDF-1α levels in cytokine-treated bone marrow endothelial cell and stromal cell condi- tioned medium
Cytokines SDF-1α (pg/mL)
BMEC-1 Primary BMSC
No 1,710±204 25,234±3,913
IL-1β (10ng/mL) 6,151±611* 24,140±3,636 IL-3 (50ng/mL) 1,537±122 24,136±3,503 IL-6 (20ng/mL) 1,984±204 NT G-CSF (100ng/mL) 2,157±205 22,863±3,729 SCF (100ng/mL) 4,068±368* 26,167±6,132 IFN-γ (100U/mL) 1,276±82 12,817±858* TNF-α (10ng/mL) 278±81* 10.861±1,787* TGF-β1 (10ng/mL) 1,492±245 9,368±4,076* The cells were incubated in serum-free medium with or without cytokines as indicated for 3 days, and the concentrations of SDF-1α in the supernatant were measured using ELISA. Data are representative of three independent experiments (mean±SD).
*P<0.05 compared to no-cytokine-treatment groups.
Abbreviations: BMSC, bone marrow stromal cells; NT, not tested.
SDF-1의 생성 증가는 관찰되지 않았다. Northern blot 분 석에서 이러한 SDF-1의 생성 감소가 이들 싸이토카인에 의한 SDF-1 mRNA 전사의 하향 조절에 기인함을 확인하 였다(Fig. 7).
고 찰
TNF-α는 수지상세포와 astrocyte에서는 CXCR4 발현 을 하향 조절하는 반면,13,15) U937 세포에서는 이를 상향 조절한다.14) TGF-β1은 TNF-α에 의한 수지상세포의 CXCR4 하향조절을 강화하고,13) IFN-γ는 말초혈액 단핵 세포의 CXCR 4발현을 하향 조절하는12) 등 다른 종류의 세 포에서는 조혈억제인자가 CXCR4의 발현의 조절에 관여한 다. 그러나 본 연구를 통해 IFN-γ, TNF-α 및 TGF-β1
등이 골수 CD34 양성세포의 CXCR4 발현에는 영향을 주지 않으며 SDF-1에 의한 화학주성의 변화도 유발하지 않음을 확인하였다. 이러한 사실은 본 연구진이 이미 보고한 바 있 는 CD34 양성세포를 각종 조혈성장인자로 단기간 처리하 는 경우에도 CXCR4 의 발현 및 SDF-1에 대한 화학주성에 변화가 없다는 점7)을 상기할 때 체외에서 조혈세포에 싸이 토카인을 처리함으로써 CXCR4 발현을 증강시키고 골수귀 소를 증진하고자하는 것이 어렵다는 점을 시사한다. 최근, 오히려 조혈세포를 체외에서 SDF-1으로 단기간 처리하는 경우 골수귀소가 향상됨이 확인된 바 있다.20) CXCR4를 발 현하는 세포에 SDF-1을 가하는 경우 CXCR4가 빠르게 하 향 조절된다는 잘 입증된 사실에 비추어 볼 때 생체 내 골 수귀소의 증진에 CXCR4의 상향조절이 필수적인 것은 아닌 것으로 보인다. 아울러 SDF-1이 골수귀소뿐 아니라 조혈
모세포 및 전구세포의 생존 및 증식에 관여한다는 최근 보
고21,22)를 참고할 때, SDF-1의 단기간 처리에 의한 골수귀
소의 증진은 화학주성의 향상보다는 조혈모세포 생존 능력 의 향상에 의한 것으로 평가된다.
IFN-γ 및 TNF-α가 CD34양성세포의 E-selectin 및 VLA-4의 발현을 경미하나마 향상시키는 것을 확인할 수 있었으나 이것이 SDF-1에 의한 경내피세포이동의 향상으 로는 연결되지 않았다. 많은 다른 종류의 세포에서와 마찬 가지로 IFN-γ 및 TNF-α에 의해 ICAM-1의 발현은 강력 하게 상향 조절되었는데, 골수귀소를 포함한 조혈세포 생 리에 ICAM-1이 어떠한 역할을 하는지는 잘 알려져 있지 않다. 다만 ICAM-1의 현저한 발현 증강에도 불구하고 자 발적 및 SDF-1 매개 경내피세포 이동 모두에서 변화가 없 었다는 것은 ICMA-1이 골수귀소에 중요한 역할을 할 가능 성은 크지 않다는 것을 의미한다.
별도의 연구에서 골수조직표본을 대상으로 한 면역조직 화학염색에서는 골수혈관내피세포에서 CXCR4의 발현이 확인되었으나(자료 제시되지 않음), BMEC-1 세포 및 사 람 골수에서 분리, 배양된 내피세포는 CXCR4를 발현하지 않았는데 그 이유는 뚜렷하지 않다. 다만 생체 내 3차원적 구조에서 벗어나면서 발현을 잃었거나, 아니면 본 연구에 서 사용한 내피세포가 실제 혈관에서 유래한 것이 아니고 조혈세포에서 유래했기 때문일 가능성을 생각해 볼 수 있 다. IFN-γ와 TNF-α가 일반적으로 혈관내피세포의 각종 유착분자 발현을 상향 조절하고 내피세포단층의 투과성을 증진시키는 것은 잘 알려진 사실로,8) 본 연구에서는 골수 혈관내피세포도 예외가 아님을 확인할 수 있었다.
골수기질세포는 VCAM-1과 ICAM-1을 약하게 발현하 고, 이러한 발현은 IFN-γ 및 TNF-α에 의해 뚜렷하게 증 가하였는 바, 기존의 보고23)와 일치하는 것이다. VCAM-1 과 ICAM-1이 각각 VLA-4 및 LFA-1을 통해 조혈세포의 기질세포에 대한 부착에 중심적인 역할을 한다는 점24,25)을 상기할 때, 이러한 조혈억제인자가 조혈세포의 골수 내 거 주의 조절에 관여한다는 것을 강력하게 시사한다.
몇몇 케모카인의 발현과 생성의 상향 또는 하향조절에 IFN-γ 및 TNF-α가 관여한다. 즉, IFN-γ와 TNF-α는 HT29-19A세포에서 MCP-1과 RANTES 발현을 유발하거 나 촉진한다.10) TNF-α는 BEAS-2B세포에서 eotaxin의 생성을 촉진한다.11) TGF-β1은 골수기질세포에서 IFN-γ 에 의한 MCP-1의 발현 증강을 강화시킨다.9) 그러나 골수 기질세포 또는 내피세포에서 SDF-1의 생성 및 분비가 어 떻게 조절되는 지에 대해서는 전혀 알려져 있지 않다. 본 연구에서는 최초로 IFN-γ, TNF-α 및 TGF-β1 등 공통 적으로 조혈억제인자로 작용하는 이들 싸이토카인이 골수 Fig. 7. Down-regulation of SDF-1 mRNA expression on
bone marrow stromal cells induced by TNF-α, IFN-γ, and TGF-β1. Primary human bone marrow stromal cells were incubated in serum-free medium with the cytokines (10 ng/mL). At indicated time points, the cells were harvested, and subjected to Northern blot analysis.
기질세포의 SDF-1 mRNA를 하향 조절함으로써 SDF-1 생성을 억제함을 보고하였다. 최근 G-CSF가 조골세포를 포함한 골수기질세포의 SDF-1 mRNA의 발현을 감소시킨 다는 보고26)가 있으나 이들 세포에 G-CSF 수용체가 발현 되는지 불분명한 상황이어서 다시 확인되어야 할 것이다.
잘 알려진 SDF-1의 기능 즉, CXCR4를 통한 화학주성이 조혈모세포의 골수귀소에 결정적인 역할을 한다는 점, 그 리고 골수와 말초혈액의 SDF-1 농도 환경의 변화 즉, 혈 중 SDF-1 농도가 높아지거나 골수 내 SDF-1 농도가 낮아 지는 경우 조혈모세포 및 전구세포의 말초구동화가 유발된 다는 점26,27)을 참조할 때 골수기질세포의 SDF-1 생성의 저하는 곧 골수귀소의 저하 또는 조혈세포의 말초 구동화 를 촉발할 수도 있을 것이다. 한편, SDF-1이 조혈모세포/
전구세포의 생존인자로 작용한다는 근간의 보고22)에 기초 할 때 이러한 조혈억제인자가 단순히 조혈세포에 직접 작 용하여 세포주기를 변조하거나 apoptosis를 촉진시킴으로 써 조혈을 억제하는 이외에, 기질세포로부터 SDF-1의 생 성을 억제하여 간접적으로 조혈억제에 관여할 가능성도 배 제할 수 없다. SDF-1은 전단류(shear flow) 하에서 integrin 유착분자의 유착력을 증강시켜 조혈전구세포의 혈 관내피세포에 공고한 유착을 유발한다.28) 생체 내 상황에 서 조혈세포가 골수로 귀소하기 위해서는 먼저 골수혈관내 피세포에 공고히 유착되어야 함은 주지의 사실로, TNF-α 에 의한 골수혈관내피세포의 뚜렷한 SDF-1 생성 저하는 골수귀소의 저하를 유발할 수 있을 것으로 생각된다.
요약하면, IFN-γ, TNF-α및 TGF-β1 등 조혈억제인 자는 선별적으로 조혈세포, 골수혈관내피세포 및 골수기질 세포의 일부 유착분자 발현을 증강시키는 한편, 기질세포 및 내피세포의 SDF-1 생성을 저하시킴으로써 조혈세포의 골수귀소의 조절에 관여하는 것으로 판단된다. 이러한 조 절이 조혈모세포이식, 조혈세포의 말초가동화 및 각종 혈 액질환에서 갖는 의미의 규명을 위해서는 보다 심화된 연 구가 필요하리라 생각된다.
요 약
배 경 : 조혈세포의 골수귀소가 각종 유착분자, CXCR4 및 SDF-1의 유기적인 상호작용을 통해 이루어진다는 것은 잘 알려져 있다. 그러나 이들 요소가 어떻게 조절되는지에 대해서는 잘 알려져 있지 않다.
방 법 : 본 연구에서는 골수귀소에 관여하는 세포 즉, 조 혈세포(골수 CD34 양성세포), 골수혈관내피세포(사람 골 수 혈관내피세포주 BMEC-1) 및 사람 골수기질세포를 대 상으로 조혈억제인자로 잘 알려져 있는 IFN-γ, TNF-α
및 TGF-β1이 귀소에 관여하는 요소 즉, 유착분자, CXCR4 및 SDF-1의 발현 또는 생성에 어떠한 영향을 미치 는지 시험관내에서 규명하고, 골수귀소 측면에서 그 의미 를 고찰하였다.
결 과 : IFN-γ, TNF-α 및 TGF-β1 모두 골수 CD34 양성세포의 CXCR4의 발현 또는 SDF-1매개 화학주성에 영향이 없었다. IFN-γ와 TNF-α는 CD34양성세포의 ICAM-1의 발현을 현저하게 증강시켰으나 그 의미는 뚜렷 치 않다. 이 두 가지 싸이토카인은 BMEC-1의 ICAM-1 발 현을 증강시키고 BMEC-1 단층의 조혈세포에 대한 투과성 을 증가시켰으며, 기질세포의 VCAM-1과 ICAM-1의 발현 을 크게 증가시켰다. TNF-α 는 내피세포의 SDF-1 생성 을, 그리고 IFN-γ, TNF-α 및 TGF-β1 모두는 기질세 포의 SDF-1 생성을 현저하게 감소시켰다.
결 론 : IFN-γ, TNF-α 및 TGF-β1 등 조혈억제인자 는 골수세포의 유착분자 발현을 증진시키고 SDF-1 생성을 감소시켜 골수귀소의 조절에 관여한다는 것을 시사한다.
조혈모세포이식, 조혈모세포의 구동화 또는 혈액질환에서 의 이러한 조절, 특히 기질세포의 SDF-1 생성의 하향적 조절이 내포하는 임상적 의의에 대한 추가적인 연구가 필 요하리라 생각된다.
참 고 문 헌
1) Hendrix PJ, Martens ACM, Hagenbeek A, Keij JF, Visser JWM : Homing of fluorescently labeled mu- rine hematopoietic stem cells. Exp Hematol 24:129- 140, 1996
2) Papayannopoulou T, Carddock C : Homing and traf- ficking of hematopoietic progenitor cells. Acta Hematol 97:97-104, 1997
3) Peled A, Petit I, Kollet O, Magid M, Ponomaryov T, Byk T, Nagler A, Ben-Hur H, Many A, Shultz L, Lider O, Alon R, Zipori D, Lapidot T : Depen- dence of human stem cell engraftment and repo- pulation of NOD/SCID mice on CXCR4. Science 283:845-848, 1999
4) Jo DY, Rafii S, Hamada T, Moore MAS : Chemo- taxis of primitive hematopoietic cells in response to stromal cell-derived factor-1. J Clin Invest 105:
101-111, 2000
5) van der Loo JCM, Ploemacher RE : Marrow- and spleen-seeding efficiencies of all murine hemato- poietic stem cell subsets are decreased by prein- cubation with hematopoietic growth factors. Blood 85:2598-2606, 1995
6) Gothot A, van der Loo JCM, Clapp W, Srour EF : Cell cycle-related changes in repopulating capacity of human mobilized peripheral blood CD34+ cells
in non-obese diabetic/severe combined immune- deficient mice. Blood 92:2641-2649, 1998
7) Jo DY, Hwang JH, Kwak SK, Shin HY, Kim SE, Yun HJ, Seong CM, Kim S : Effects of short-term incubation of hematopoietic cells with hematopoietic growth factors on CXCR4 expression and stromal cell-derived factor-1-mediated transendothelial mi- gration. Korean J Hematol 36:324-334, 2001 8) Carlos TM, Harlan JM : Leukocyte-endothelial ad-
hesion molecules. Blood 84:2068-2101, 1994
9) Gautam SC, Noth CJ, Janakiraman N, Pindolia KR, Chapman RA : Induction of chemokine mRNA in bone marrow stromal cells: modulation by TGF- beta and IL-4. Exp Hematol 23:482-491, 1995 10) Warhurst AC, Hopkins SJ, Warhusrt G : Interferon
gamma induces differential upregulation of alpha and beta chemokine secretion in colonic epithelial cell lines. Gut 42:208-213, 1998
11) Stellato C, Matsukura S, Fal A, white J, Beck LA, Proud D, Schleimer RP : Differential regulation of epithelial-derived C-C chemokine expression by IL-4 and the glucocorticoid budesonide. J Immunol 163: 5624-5632, 1999
12) Shirazi Y, Pitha PM : Interferon down-regulates CXCR4(fusin) gene expression in peripheral blood mononuclear cells. J Hum Virol 1:69-76, 1998 13) Sato K, Kawasaki H, Nagayama H, Enomoto M,
Morimoto C, Tadokoro K, Juji T, Takahashi TA : TGF-beta 1 reciprocally controls chmotaxis of hu- man peripheral blood monocyte-derived dendritic cells via chemokine receptors. J Immunol 164:
2285-2295, 2000
14) Biswas P, Mantelli B, Delfanti F, Cota M, Vallanti G, de Filippi C, Menozzi M, Vienzi E, Lazzarin A, Poli G : Tumor necrosis factor-alpha drives HIV-1 replication in U937 cell clones and upregulates CXCR4. Cytokine 7:55-59, 2001
15) Han Y, Wang J, He T, Ransohoff RM : TNF-alpha down-regulates CXCR4 expression in primary mu- rine astrocytes. Brain Res 888:1-10, 2001
16) Verfaillie C, Blakolmer K, McGlave P : Purified pri- mitive human haematopoietic progenitor cells with long-term in-vitro repopulating capacity adhere selectively to irradiated bone marrow stroma. J Exp Med 172:509-520, 1990
17) Itoh K, Tezuka H, Sakoda H, Konno M, magata K, Uchiyama T, Uchino H, Mori KJ : Reproducible establishment of hemopoietic supportive cells lines from murine bone marrow. Exp Hematol 17:145- 153, 1989
18) Candal FJ, Rafii A, Parker JT, Ades EW, Ferris B, Nachman RL, Kellar KL : BMEC-1: a human bone marrow microvascular endothelial cell line with primary cell characteristics. Microvasc Res 52:221- 234, 1996
19) Breems DA, Blokland EAW, Neben S, Ploemacher RE : Frequency analysis of human primitive hema- topoietic stem cell subsets using a cobblestone area forming cell assay. Leukemia 8:1095-1104, 1994
20) Plette PA, Frankovitz SM, Wolber FM, Abonour R, Orschell-Traycoff CM : Treatment of circulating CD34+ cells with SDF-1α or anti-CXCR4 anti- body enhances migration and NOD/SCID repopu- lating potential. Exp Hematol 30:1061-1069, 2002 21) Cashman J, Clark-Lewis I, Eaves A, Eaves C :
Stromal-derived factor-1 inhibited the cycling of very primitive human hematopoietic cells in vitro and in NOD/SCID mice. Blood 99:792-799, 2002 22) Lstaillade JJ, Clay D, Bourin P, Herodin F, Dupuy
C, Jasmin C, Le Bousse-Kerdiles M-C : Stromal cell-derived factor 1 regulates primitive hemato- poiesis by suppressing apoptosis and by promoting G0/G1 transition in CD34+ cells: Evidence for an autocrine/paracrine mechnism. Blood 99:1117-1129, 2002
23) Hideshima T, Chauhan D, Schlossman R, Richard- son P, Andeson KC : The role of tumor necrosis factor alpha in the pathophysiology of multiple myeloma: Therapeutic applications. Oncogene 20:
4519-4527, 2001
24) Teixido J, Jemler ME, Greeberger JS, Naklesaria P : Role of β1 and β2 integrins in the adhesion of human CD34hi stem cells to bone marrow stroma.
J Clin Invest 90:358-367, 1992
25) Simmons PJ, Masinovsky B, Longenecker BM, Berenson R, Torok-Storb B, Gallatin WM : Vascular cell adhesion molecule-1 expressed by bone mar- row stromal cells mediates the binding of hema- topoietic progenitor cells. Blood 80:388-395, 1992 26) Petit I, Szyer-Kravitz M, Nagler A, Lahav M, Peled
A, Haber L, Ponomaryov T, Taichman RS, Aren- zana-Seisdedos F, Fujii N, Sandbank J, Zipori D, Lapidot T : G-CSF induces stem cell mobilization by decreasing bone marrow SDF-1 and up-re- gulating CXCR4. Nature Immunol 3:687-694, 2002 27) Hattori K, Heissig B, Tashiro K, Honjo T, Tateno
M, Shieh JH, Hackett NR, Quitoriano, MS, Crystal RG, Rafii S, Moore MAS : Plasma elevation of stromal cell-derived factor-1 induces mobilization of mature and immature hematopoietic progenitor and stem cells. Blood 97:3354-3360, 2002
28) Peled A, Brabovsky V, Habler L, Sandbank J, Arezana-Seisdedos F, Petit I, Ben-Hur H, Lapidot T, Alon R : The chemokine SDF-1 stimulates inte- grin-mediated arrest of CD34+ cells on vascular endothelium under shear flow. J Clin Invest 104:
1199-1211, 1999