40 책임저자:최영현, 614-052, 부산시 진구 양정동 산 45
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4037 E-mail: [email protected]
접수일:2009년 3월 5일, 게재승인일:2009년 3월 16일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, College of Oriental Medicine, Graduate School, Dongeui University, Yangjung-dong, Busanjin-gu, Busan 614- 714, Korea
Tel: +82-51-850-7413, Fax: +82-51-853-4037 E-mail: [email protected]
Bcl-2 과발현 인체혈구암세포의 인진쑥 유래 Esculetin에 의한 Apoptosis 유발에서 Extracellular-regulated Kinase의 역할
1동의대학교 블루바이오 소재개발센터, 2자연과학대학 생명응용학과, 3생활과학대학 식품영양학과,
한의과대학 4생화학교실, 5생리학교실 및 6대학원 바이오물질제어학과,
7부산대학교 한의학전문대학원 경락구조의학부
박 철1ㆍ권현주1,2,6ㆍ황혜진1,3ㆍ이용태1,5ㆍ최병태7ㆍ김병우1,2,6ㆍ최영현1,4,6
Involvement of Extracellular-Regulated Kinase in Esculetin-Induced Apoptosis of Bcl-2 Overexpressing Human Leukemia U937 Cells
Cheol Park1, Hyun Ju Kwon1,2,6, Hye Jin Hwang1,3, Yong Tae Lee1,5, Byung Tae Choi7, Byung-Woo Kim1,2,6 and Yung Hyun Choi1,4,6
1Blue-Bio Industry RIC, 2Department of Life Science and Biotechnology, College of Natural Science,
3Department of Food and Nutrition, College of Human Ecology, Departments of 4Biochemistry and
5Physiology, College of Oriental Medicine, 6Department of Biomaterial Control, Graduate School, Dongeui University, Busan 614-714, 7Division of Meridian and Structural Medicine,
School of Oriental Medicine, Pusan National University, Busan 609-735, Korea
Bcl-2 is the prototypic anti-apoptotic protein involved in the regulation of apoptosis. Commonly, overexpression of Bcl-2 confers resistance to the apoptotic effect of chemo- and radiotherapy. Esculetin is a coumarin compound that is found in various natural plant products and induces apoptosis in several types of human cancer cells. However, the underlying mechanisms of its action are not completely understood. In the present study, we observed overexpressing Bcl-2 attenuated esculetin-induced up-regulation of death receptor 4 (DR4), down-regulation of X-linked inhibitor of apoptosis protein (XIAP) and β-catenin, and degradation of DNA fragmentation factor 45/inhibitor of caspase-activated DNase (DFF45/ICAD). However, HA14-1, a small molecule Bcl-2 antagonist, increased sensitivity to the apoptotic effect of the esculetin in Bcl-2 overexpressing U937/Bcl-2 cells that correlated with the loss of mitochondrial membrane potential (MMP). Furthermore, inactivation of extracellular-regulated kinase (ERK) by PD98059 significantly decreased esculetin-induced cell death and restored XIAP, β-catenin, phospholipase Cγ-1 (PLCγ1) and DFF45/ICAD in pretreatment with HA14-1 in Bcl-2 overexpressing U937/Bcl-2 cells. These results demonstrate an additional mechanism of regulation of cell survival mediated by Bcl-2, namely through ERK. Therefore, directed inhibition of Bcl-2 may alter diverse pathways controlling cell survival and overcome the apoptotic resistance that is the hallmark of human leukemia cells. (Cancer Prev Res 14, 40-47, 2009)
Key Words: Esculetin, Apoptosis, Bcl-2, ERK
서 론
Apoptosis는 배아의 형성, 면역과 신경계의 발달 및 암 발생 등과 같은 생체 내의 중요한 생리 및 병리학적인 상태에서 흔히 관찰되는 세포 죽음의 한 기전으로 중요 한 역할을 한다.1,2) 특히 종양의 자연 치유나 항암제에 의해 암세포의 죽음이 유발될 때 이러한 apoptosis가 많이 관찰되므로 항암제의 항종양기전 연구에 있어서 많은 연구자들의 관심을 받고 있다. Apoptosis 과정은 extrinsic pathway 및 intrinsic pathway로 크게 구별되어지는데, ex- trinsic pathway는 mitochondria 비의존적인 apoptosis 과정으 로서 apoptotic ligand가 Fas, TNF-related apoptosis-inducing ligand (TRAIL)-receptor 1 (TRAIL-R1, DR4), TRAIL-R2 (DR5) 등과 같은 death receptor와 결합함으로서 유발되어 진 다.3,4) 이와 반대로 intrinsic pathway는 mitochondria 의존적 인 과정으로서 여러 가지 외부 자극에 의하여 mitochond- rial membrane permeability (MMP, Δψm)의 변화가 유발되 어 mitochondria 내에 존재하는 여러 종류의 apoptotic molecule들이 세포질로 방출됨으로서 시작된다.5,6) MMP 변화를 유발하는데 있어서 Bcl-2 family가 중요한 역할을 하는 것으로 알려져 있는데, 특히 apoptosis 유발을 억제 하는 것으로 알려진 anti-apoptotic 유전자인 Bcl-2는 대부 분의 암세포에서 과발현되어 있으며 항암제 내성의 원 인이 된다고 알려져 있다.7∼9) 따라서 Bcl-2의 발현을 조 절하는 것은 암치료의 효율을 높이는 한 방법으로 유용 하게 사용될 수 있다.
한편 mitogen-activating protein kinases (MAPKs)는 serine/
threonine protein kinases로서 세포의 성장, 분화, 증식, 사 멸 및 스트레스 반응 등과 같은 여러 가지 세포 반응을 조절하는 것으로 알려져 있으며, c-Jun NH2-terminal kinase (JNK), p38-MAPK 및 extracellular signal-regulated kinase (ERK) 로 구성되어 있다.10∼12) 일반적으로 JNK 및 p38-MAPK는 proinflammatory cytokines, UV irradiation, heat, osmotic shock, hydrogen peroxide 및 DNA 손상 등에 의하여 활성화되어 스트레스 반응, 성장 억제 및 apoptosis 유발에 관여하는 반면에, ERK의 경우는 growth factors, cytokines 및 phorbol esters 등과 같은 mitogenic stimuli에 의하여 활성화 되어 세포의 성장 및 분화에 관여하는 것으로 알려져 있다.10∼14) 하지만 최근 연구에 따르면 세포가 처한 환경 변화에 따라 이들 MAPKs 모두 성장 억제 및 apoptosis 유발과 세포의 성장 및 분화에 각기 다르게 관여할 수 있다고 보고되어 지고 있다.15∼17)
Lipoxygenase inhibitor로 알려진 바 있는 esculetin (6,7-di-
hydroxycoumarin)은 인진쑥(Artemisia scoparia), 비쑥(A. ca- pillaries), 갯질경이(Ceratostigma willmottianum) 및 레몬(Citrus limonia)의 잎 등에 많이 함유되어 있는 페놀계 화합물이 다.18) 최근 연구들에 의하면 esculetin은 Chinese hamster lung fibroblast에서 H2O2에 의하여 유도되는 손상에 대한 항산화작용을 가지는 것으로 알려진 바 있으며, croton oil ear test에서 anti-inflammatory effect 및 peripheral analge- sic activity를 가지며 3T3-L1 세포에서 adipogenesis 억제효 과를 가지는 것으로 보고된 바 있다.19∼21) 또한 인체 혈 구암세포인 U937 세포에서 G1기에서 세포주기 억제 및 apoptosis를 유발하는 것으로 확인되었지만,16,22) 항암제 내성의 가장 큰 원인 중의 하나인 Bcl-2 과발현 암세포에 서 esculetin의 항암성에 대해서는 알려진 바 없다.
따라서 본 연구에서는 선행연구 결과에 준하여16,22) 인 진쑥에서 유래된 esculetin이 인체 혈구암세포인 U937 세 포 및 Bcl-2가 과발현된 U937/Bcl-2 세포에16) 미치는 apoptosis 유발 정도를 비교 분석하였고, Bcl-2 및 MAPKs 가 esculetin에 의하여 유발되는 apoptosis에 어떠한 영향을 미치는 지를 조사하여 유의적인 결과를 얻었기에 이를 보고하는 바이다.
재료 및 방법 1. 실험재료
Esculetin 및 5,5,6,6-tetrachloro-1,1,3,3-tetraethylbenzimida- zolyl carbocyanine iodide (JC-1)은 Sigma-Aldrich (St. Louis, MO, USA)에서 구입하였다. Bcl-2, FasL, death receptor (DR) 4, DR5, XIAP, β-catenin, DNA fragmentation factor 45/
inhibitor of caspase-activated DNase (DFF45/ICAD), phos- pholipase Cγ-1 (PLCγ1) 및 actin 항체는 Santa Cruz Biote- chnology Inc. (Santa Cruz, CA, USA) 및 CalBiochem (San Die- go, CA, USA)에서 구입하였으며, 2차 항체로 사용된 per- oxidase-labeled donkey anti-rabbit 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Life Science Corp.
(Arlington Heights, IL, USA)에서 구입하였다.
2. 세포 배양 및 형태관찰
인체혈구암 U937 세포는 American Type Culture Collec- tion (ATCC, Rockville, MD, USA)에서 분양 받았으며 90%의 RPMI-1640 배지(Gibco BRL, Grand Island, NY, USA), 10%
fetal bovine serum (FBS)에 2 mM glutamine, 100 U/ml penicil- lin 및 100μg/ml streptomycin (Gibco BRL)이 포함된 배지를 사용하여 5% CO2, 37°C의 조건하에서 배양하였다. 또한 Bcl-2가 과발현된 U937/Bcl-2 세포는 0.7μg/ml geneticin
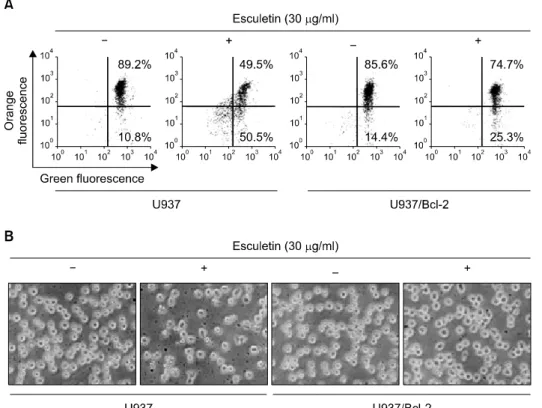
Fig. 1. Bcl-2 overexpression pro- tects against esculetin-induced loss of MMP and morphological changes in U937 cells. U937 and U937/Bcl-2 cells were seeded at 2×105 cells/ml and then incuba- ted with 30μg/ml of esculetin for 6 h. (A) The MMP was measured with the lipophilic cationic probe, JC-1. The cells were exposed esculetin, after which flow cytome- tric analyses were performed as described in materials and me- thods. Data are expressed as means of three independent ex- periments. (B) Morphological chan- ges of U937 and U937/Bcl-2 cells after treatment with esculetin were visualized by an inverted micro- scope. Magnification, ×200.
(G418 sulfate, Calbiochem)을 첨가하여 배양하였다.16) Esculetin 처리에 의한 U937 및 U937/Bcl-2 세포의 형태변 화는 적정 시간동안 esculetin을 처리하여 배양한 후, 도립 현미경(inverted microscope, Carl Zeiss, Germany)을 이용하 여 200배의 배율로 관찰하였다.
3. Flow cytometry 분석
Esculetin이 처리된 U937 및 U937/Bcl-2 세포를 모은 다 음 CycleTEST PLUS DNA REAGENT Kit (Becton Dickinson, San Jose, CA, USA)를 이용하여 4°C, 암실에서 30분 동안 고정 및 염색을 하였다. 염색된 세포를 35-mm mesh를 이 용하여 단일세포로 분리한 후 FACSCalibur (Becton Dickin- son)를 이용하여 형광반응에 따른 Cellular DNA content 및 histogram을 CellQuest software 및 ModiFit LT (Becton Dic- kinson) 프로그램을 이용하여 분석하였다.
4. Mitochondrial membrane potential (MMP, Δψm) 분석
Esculetin에 의한 MMP 변화 정도의 측정은 선행 방법 에 준하여16) 500μl의 PBS에 10μM JC-1을 처리하여 20 분간 37°C에서 반응시킨 다음 2,000 rpm으로 원심 분리 하여 상층액을 제거하고 다시 500μl의 차가운 PBS를 첨 가하여 35-mm mesh를 이용하여 단일세포로 분리한 후
FACSCalibur를 이용하여 분석하였다.
5. Protein extraction and western blot analysis
준비된 암세포에 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenyme- thylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 처 리하여 단백질을 분리한 다음 Bio-Rad 단백질 정량 시약 (Bio-Rad, Hercules, CA, USA)을 이용하여 정량하였다. 동 량의 단백질을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리한 다음 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이시켰다. 단백질이 전이된 mem- brane에 1차 및 2차 항체를 반응시킨 다음 Enhanced Che- miluminoesence (ECL) 용액(Amersham Life Science Corp., Ar- lington Heights, IL, USA)을 처리하여 암실에서 X-ray film 에 감광시켜 특정단백질의 양을 분석하였다.
결 과
1. Bcl-2 과발현이 esculetin에 의한 MMP 변화 및 형태에 미치는 영향
인체 혈구암세포인 U937 세포에서 esculetin에 의한 apoptosis 유발에 있어서 Bcl-2 과발현이 미치는 영향을
Fig. 3. Co-treatment with esculetin and HA14-1 restores apoptosis in U937/Bcl-2 cells. (A) After co-treat- ment with esculetin and HA14- 1 for 6 h, the cells were collected and stained with CycleTEST PLUS DNA REAGENT Kit for flow cyto- metry analysis. The percentage of cells with hypodiploid DNA (sub- G1 phase) content represent the fractions undergoing apoptotic DNA degradation. (B) Morphologi- cal changes of U937/Bcl-2 cells after co-treatment with esculetin and HA14-1 were visualized by an inverted microscope. Magnifica- tion, x200. (C) The cells were ex- posed esculetin and HA14-1, after which the levels of MMP were me- asured with the lipophilic cationic probe, JC-1, using flow cytometric analyses as described in Materi- als and methods. All data are ex- pressed as means of three inde- pendent experiments.
Fig. 2. Bcl-2 overexpression protects against esculetin-induced down-regulation or proteolytic cleavage of DR4, XIAP, β-cate- nin and DFF45/ICAD. After treatment with esculetin for 6 h, the cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
먼저 비교하였다. Fig. 1A에 나타낸 바와 같이 U937 세포 에서는 esculetin 처리에 의하여 MMP의 loss가 유발되는 것으로 나타났지만 Bcl-2가 과발현된 세포에서는 MMP 의 loss가 현저하게 억제되는 것으로 나타났으며, escule- tin 처리에 따른 형태변화도 Bcl-2 과발현에 의하여 억제 되는 것으로 관찰되었다. 아울러 esculetin이 처리된 U937 세포의 경우 전반적인 세포의 응축과 단편화 현상이 관 찰되었으나, Bcl-2가 과발현된 세포(U937/Bcl-2)에서는 이 러한 현상이 나타나지 않았다(Fig. 1B). 따라서 Bcl-2가 esculetin에 의하여 유발되는 mitochondria의 기능 손상을 억제함으로서 apoptosis 유발을 막아주는 것으로 생각된 다.
2. Bcl-2 과발현이 apoptosis 관련 단백질의 발현에 미치는 영향
U937 및 U937/Bcl-2 세포에서 esculetin이 apoptosis 관련 단백질들의 발현에 어떠한 영향을 미치는 조사한 결과 는 Fig. 2에 나타난 바와 같다. 결과에서 알 수 있듯이 Bcl-2, FasL 및 DR5 단백질의 발현에는 두 세포주 모두에
서 큰 변화가 관찰되지 않았지만 DR4, XIAP, β-catenin 및 DFF45/ICAD 단백질들은 esculetin 처리에 의한 발현 변화가 Bcl-2 과발현에 의하여 억제되는 것으로 나타났 다. 따라서 U937/Bcl-2 세포에서의 apoptosis 억제는 이들
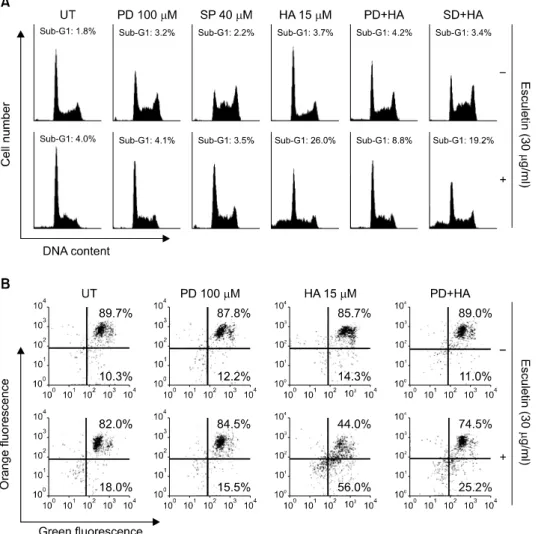
Fig. 4. Involvement of ERK path- way in esculetin plus HA14-1- induced apoptosis in U937/Bcl-2 cells. Cells were pretreated with PD98059 (100μM) or SP600125 (40μM) for 1 h before treatment with 30μg/ml of esculetin and 15 μM of HA14-1 for 6 h. (A) The cells were collected and stained with CycleTEST PLUS DNA REAG- ENT Kit for flow cytometry ana- lysis. The percentage of cells with hypodiploid DNA (sub-G1 phase) content represent the fractions undergoing apoptotic DNA degra- dation. (B) The MMP was mea- sured with the lipophilic cationic probe, JC-1. All data are express- ed as means of three indepen- dent experiments.
단백질들의 발현 변화 억제와 관련성이 있음을 추정할 수 있었다.
3. U937/Bcl-2 세포에서 esculetin 및 HA14-1 처리 에 의한 apoptosis 유발
U937 세포에서 esculetin에 의하여 유발되었던 MMP 및 apoptosis 관련 단백질들의 변화가 Bcl-2의 과발현에 의하 여 억제되는 것으로 관찰되었기에, 이러한 현상들을 재확 인하기 위하여 Bcl-2 억제제인 HA14-1을 이용하여 U937/
Bcl-2 세포에서 Bcl-2를 억제하였을 경우 esculetin에 의한 apoptosis가 다시 유발되는 지를 확인하였다. 먼저 Fig. 3A 및 B에 나타난 바와 같이 U937/Bcl-2 세포에서는 esculetin 에 의한 sub-G1기의 증가 및 형태 변화가 관찰되지 않았 지만 HA14-1을 처리하였을 경우에는 sub-G1기가 현저하 게 증가하였으며, 세포의 형태도 현저하게 변하는 것으 로 나타났다. 이러한 sub-G11기의 증가 및 형태 변화에 mitochondria의 연관성을 확인한 결과, Fig. 3C에 나타난 바와 같이 HA14-1 처리에 의하여 MMP의 loss도 증가하
는 것으로 나타났으므로 U937/Bcl-2 세포에서 esculetin 및 HA14-1 처리에 의한 apoptosis 유발에 있어서 mitochondria 가 중요한 역할을 하는 것으로 생각된다.
4. ERK pathway가 esculetin 및 HA14-1에 의한 sub-G1 및 MMP 변화에 미치는 영향
U937/Bcl-2 세포에서 esculetin 및 HA14-1 처리에 의하 여 유발되는 apoptosis에 있어서 MAPKs pathway가 어떠한 영향을 미치는 지를 확인한 결과는 Fig. 4에 나타난 바와 같다. 먼저 Fig. 4A에서와 같이 U937/Bcl-2 세포에 escule- tin을 단독으로 처리하였을 경우에는 sub-G1기의 증가가 관찰되지 않았지만 esculetin 및 HA14-1을 같이 처리하였 을 경우에는 sub-G1기가 현저하게 증가하여 약 26% 정 도로 나타났다. 하지만 ERK 억제제인 PD98059를 선처리 후 esculetin 및 HA14-1을 처리하였을 경우에는 sub-G1기 에 해당하는 세포가 약 8.8%로 매우 억제되었다. 또한 Fig. 4B에서와 같이 MMP의 loss를 관찰한 결과도 PD9 8059의 선처리에 의하여 현저하게 억제되었음을 알 수
Fig. 5. The ERK pathway plays important roles in esculetin plus HA14-1-induced down-regulation and proteolytic cleavage of XIAP, β-catenin, PLCγ1 and DFF45/ICAD in U937/Bcl-2 cells. Cells were pretreated with PD98059 (100μM) for 1 h before treatment with 30μg/ml of esculetin and 15μM of HA14-1 for 6 h. The cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
있었다. 그러나 p38-MAPK 억제제인 SP600125의 선처리 는 esculetin 및 HA14-1의 동시처리에 의한 apoptosis 유발 은 크게 억제하지 못하여, U937/Bcl-2 세포에서 ERK pa- thway가 esculetin 및 HA14-1에 의하여 유발되는 apoptosis 에 중요하게 관여하고 있음을 알 수 있었다.
5. ERK pathway가 esculetin 및 HA14-1에 의한 apoptosis 관련 단백질의 발현에 미치는 영향
U937/Bcl-2 세포에서 esculetin 및 HA14-1 처리에 의하 여 유발되는 apoptosis 관련 단백질들의 변화와 ERK pathway와의 관계를 조사한 결과는 Fig. 5에 나타난 바와 같다. 결과에서 알 수 있듯이 esculetin 및 HA14-1 처리에 의하여 유발되었던 XIAP와 caspase-3의 기질 단백질들인 β-catenin, PLCγ1 및 DFF45/ICAD의 발현 감소 및 단편화 현상이 PD98059 처리에 의한 ERK pathway의 억제에 의 하여 원래대로 회복되는 것으로 나타났다. 이는 Bcl-2 과 발현 U937 세포에서 esculetin 및 HA14-1에 의하여 유도 되는 apoptosis에 ERK pathway가 관여하고 있음을 확인시 켜 주는 결과라고 할 수 있다.
고 찰
본 연구팀은 선행연구에서 esculetin이 인체 혈구암세 포인 U937 세포의 증식을 억제하였으며, 이는 세포주기
G1 arrest 및 apoptosis의 유발과 연관성이 있음을 제시한 바 있다.16,22) 본 연구에서는 이를 바탕으로 하여 대부분 의 인체 암세포에 과발현되어 있으며 여러 가지 항암제 에 의한 암치료에 있어서 저항성을 가지는 원인 중 하나 인 Bcl-2를 과발현시켰을 경우 esculetin에 의하여 유발되 는 apoptosis가 억제되는 지를 확인하였다. 이를 위하여 U937 세포 및 Bcl-2가 과발현된 U937/Bcl-2 세포에 각각 30μg/ml 농도의 esculetin을 6시간 동안 처리한 다음 MMP의 변화 및 형태 변화의 정도를 조사한 결과, U937 세포에서는 MMP의 loss가 증가하면서 심한 형태적 변형 이 관찰되었지만 U937/Bcl-2 세포에서는 MMP의 loss가 현저하게 억제되었고 형태적 변형도 거의 나타나지 않 는 것으로 관찰되었다(Fig. 1). 그러나 Bcl-2 저해제인 HA14-1을 esculetin과 동시에 처리하였을 경우 Bcl-2가 과 발현된 U937 세포에서도 apoptosis가 유도되었으며, 이때 ERK pathway과 관여하는 것으로 나타났다.
Apoptosis 유발에는 여러 가지 단백질들이 관여하지만 특히 DR4의 경우는 TRAIL과의 결합을 통하여 extrinsic pathway를 통한 apoptosis 유발에 중요한 역할을 하며, IAP family 중 가장 강력한 유전자인 XIAP의 경우에는 활성 화된 caspase와의 높은 친화력을 가지며 apoptosis를 억제 하는 것으로 알려져 있다.23,24) 또한 caspase가 활성화 되 면 여러 가지 기질 단백질들을 분해시킴으로서 apoptosis 를 유발하게 되는데 대표적인 기질 단백질로는 세포내 골격의 유지와 다양한 부착성 세포의 전사 조절에 관여 하는 β-catenin과 DNA 단편화에 관여하는 DFF45/ICAD 등이 있다.25∼27) 따라서 U937 및 U937/Bcl-2 세포에서 es- culetin이 이러한 apoptosis 유발 관련 단백질들의 발현에 어떠한 영향을 미치는 지를 확인한 결과, U937 세포에서 는 DR4, XIAP, β-catenin 및 DFF45/ICAD의 발현 감소와 단편화 현상이 관찰되었지만 U937/Bcl-2 세포에서는 아 무런 변화가 나타나지 않았다(Fig. 2). 이는 Bcl-2의 과발 현에 의하여 esculetin에 의한 U937 세포의 apoptosis 유도 가 억제되었음을 직접적으로 보여주는 증거로서 escule- tin에 의한 apoptosis에 Bcl-2가 중심적인 역할을 할 수 있 음을 의미하며, U937 세포에서 Bcl-2의 과발현으로 escu- letin에 대한 세포독성 저항성이 획득되었음을 의미한다.
아울러 U937/Bcl-2 세포에서의 apoptosis 억제가 Bcl-2의 과발현과 직접적인 연관이 있는 지를 재확인하기 위하 여 HA14-1을 이용하여 확인하였는데, HA14-1은 Bcl-2와 결합함으로서 Bcl-2의 기능을 억제하여 apoptosis 유발을 막는 것으로 보고된 바 있는 강력한 Bcl-2 억제제이다.28,29) Fig. 3에 나타난 바와 같이 esculetin 단독 처리군에 비하 여 HA14-1과 esculetin을 동시에 처리하였을 경우에는
sub-G1기 세포 빈도, 형태 변화 및 MMP loss의 정도가 현저하게 증가되어 Bcl-2의 과발현으로 항암제 저항성을 지닐 수 있는 암세포에서 Bcl-2 활성 억제제의 동시 처리 는 항암제 내성을 극복할 수 있는 새로운 방법이 될 수 있음을 알 수 보여주었다. 한편 이러한 현상들이 세포의 성장, 분화, 증식 및 사멸 등과 같은 여러 가지 세포 반응 조절에 관여하는 MAPKs와 어떠한 관련이 있는지를 확 인하기 위하여 몇 가지 MAPK 특이적 저해제를 이용하 여 조사한 결과, U937/Bcl-2 세포에서 HA14-1 및 esculetin 처리에 의해 유발되었던 apoptosis가 ERK inhibitor인 PD 98059의 선처리에 의하여 다시 억제되었으며 MMP loss 의 정도도 현저하게 감소하는 것으로 나타났다(Fig. 4).
또한 Bcl-2의 과발현에 의하여 변화가 억제되었던 apoptosis 유발 관련 단백질들인 XIAP, β-catenin, PLCγ1 및 DFF45/ICAD의 발현 감소와 단편화 현상이 다시 나타 남을 알 수 있었다(Fig. 5). 이는 Bcl-2 과발현에 esculetin의 항암 내성 극복을 위한 HA14-1과의 동시 처리에 ERK pathway가 직접적으로 관여하고 있음을 보여주는 결과 이다.
이상의 결과들을 살펴볼 때 esculetin은 U937 세포에서 DR4의 발현 증가, 미토콘드리아막 전위의 변화 및 caspa- se 기질 단백질들의 분해 등을 통하여 apoptosis를 유발하 였음을 알 수 있었다. 또한 Bcl-2 과발현에 따른 esculetin 내성 극복을 위한 Bcl-2 저해제의 혼용처리에 의한 apop- tosis 유발에는 ERK pathway가 관여하고 있음을 알 수 있 었다.
결 론
Apoptosis 조절에 중요한 역할을 하는 Bcl-2는 prototypic anti-apoptotic 단백질로서 작용하며, 일반적으로 Bcl-2 유 전자의 과발현은 화학요법 및 방사선 조사에 의한 암세 포의 apoptosis 유발에 저항인자로 작용한다. 인진쑥에서 유래된 esculetin에 의한 인체혈구암 U937 세포의 apop- tosis 유발은 DR4의 발현 증가, XIAP와 β-catenin 발현의 감소 및 DFF45/ICAD 단백질의 단편화를 수반하였으나, Bcl-2의 과발현은 esculetin에 의한 apoptosis 유발 효과를 차단시켰다. 그러나 Bcl-2 antagonist인 HA14-1을 Bcl-2가 과발현된 U937/Bcl-2 세포에 esculetin과 동시에 처리했을 경우 apoptosis가 다시 유발되었으며, 이는 MMP의 감소 와 연관성이 있었다. 또한 ERK 저해제 처리는 U937/
Bcl-2 세포에서 apoptosis 유발 효과를 더욱 증가시켰다.
본 연구의 결과는 Bcl-2 과발현에 의한 생존전략에 ERK 경로가 관여함을 보여주는 것이며, Bcl-2 발현의 조절을
통하여 암세포의 약재 저항성을 극복할 수 있음을 시사 하여 준다.
감사의 글
이 연구는 지식경제부·부산광역시 지원 지역혁신센터 사업 동의대학교 블루바이오 소재 개발 및 실용화 지원 센터(RIC08-06-07) 및 2008년도 부산테크노파크 지원에 의하여 이루어진 결과입니다.
참 고 문 헌
1) Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics.
Br J Cancer 26, 239-257, 1972.
2) Evans VG. Multiple pathways to apoptosis. Cell Biol Int 17, 461-476, 1993.
3) Ashkenazi A. Targeting death and decoy receptors of the tumour-necrosis factor superfamily. Nat Rev Cancer 2, 420- 430, 2002.
4) Kischkel FC, Lawrence DA, Chuntharapai A, Schow P, Kim KJ, Ashkenazi A. Apo2L/TRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity 12, 611-620, 2000.
5) Schultz DR, Harrington WJ Jr. Apoptosis: programmed cell death at a molecular level. Semin Arthritis Rheum 32, 345-369, 2003.
6) Zamzami N, Susin SA, Marchetti P, Hirsch T, Gómez-Monterrey I, Castedo M, Kroemer G. Mitochondrial control of nuclear apoptosis. J Exp Med 183, 1533-1544, 1996.
7) Jeong SY, Seol DW. The role of mitochondria in apoptosis.
BMB Rep 41, 11-22, 2008.
8) Cory S, Huang DC, Adams JM. The Bcl-2 family: roles in cell survival and oncogenesis. Oncogene 22, 8590-8607, 2003.
9) Bouillet P, Huang DC, O'Reilly LA, Puthalakath H, O'Connor L, Cory S, Adams JM, Strasser A. The role of the pro-apoptotic Bcl-2 family member bim in physiological cell death. Ann N Y Acad Sci 926, 83-89, 2000.
10) Sigoillot FD, Evans DR, Guy HI. Growth-dependent re- gulation of mammalian pyrimidine biosynthesis by the protein kinase A and MAPK signaling cascades. J Biol Chem 277, 15745-15751, 2002.
11) Chang L, Karin M. Mammalian MAP kinase signaling cas- cades. Nature 410, 37-40, 2001.
12) Geilen CC, Wieprecht M, Orfanos CE. The mitogen-activated protein kinases system (MAP kinase cascade): its role in skin signal transduction. A review. J Dermatol Sci 12, 255-262, 1996.
13) Ichijo H. From receptors to stress-activated MAP kinases.
Oncogene 18, 6087-6093, 1999.
14) Beyaert R, Cuenda A, Vanden Berghe W, Plaisance S, Lee JC, Haegeman G, Cohen P, Fiers W. The p38/RKmitogen activated protein kinase pathway regulates interleukin-6 synthesis response to tumor necrosis factor. EMBO J 15, 1914-1923, 1996.
15) Lunghi P, Giuliani N, Mazzera L, Lombardi G, Ricca M, Corradi A, Cantoni AM, Salvatore L, Riccioni R, Costanzo A, Testa U, Levrero M, Rizzoli V, Bonati A. Targeting MEK/
MAPK signal transduction module potentiates ATO-induced apoptosis in multiple myeloma cells through multiple signa- ling pathways. Blood 112, 2450-2462, 2008.
16) Park C, Jin CY, Kim GY, Choi IW, Kwon TK, Choi BT, Lee SJ, Lee WH, Choi YH. Induction of apoptosis by esculetin in human leukemia U937 cells through activation of JNK and ERK. Toxicol Appl Pharmacol 227, 219-228, 2008.
17) Thoms HC, Dunlop MG, Stark LA. p38-mediated inactivation of cyclin D1/cyclin-dependent kinase 4 stimulates nucleolar translocation of RelA and apoptosis in colorectal cancer cells.
Cancer Res 67, 1660-1669, 2007.
18) Morita I, Takahashi R, Saito Y, Murota S. Stimulation of eicosapentaenoic acid metabolism in washed human platelets by 12-hydroperoxyeicosatetraenoic acid. J Biol Chem 258, 10197-10199, 1983.
19) Kim SH, Kang KA, Zhang R, Piao MJ, Ko DO, Wang ZH, Chae SW, Kang SS, Lee KH, Kang HK, Kang HW, Hyun JW. Protective effect of esculetin against oxidative stress-in- duced cell damage via scavenging reactive oxygen species. Acta Pharmacol Sin 29, 1319-1326, 2008.
20) Yang JY, Della-Fera MA, Hartzell DL, Nelson-Dooley C, Hausman DB, Baile CA. Esculetin induces apoptosis and inhibits adipogenesis in 3T3-L1 cells. Obesity (Silver Spring) 14, 1691-1699, 2006.
21) Tubaro A, Del Negro P, Ragazzi E, Zampiron S, Della Loggia R. Anti-inflammatory and peripheral analgesic activity of
esculetin in vivo. Pharmacol Res Commun 20(5 Suppl), 83-85, 1988.
22) Lee SH, Park C, Jin CY, Kim GY, Moon SK, Hyun JW, Lee WH, Choi BT, Kwon TK, Yoo YH, Choi YH.
Involvement of extracellular signal-related kinase signaling in esculetin induced G1 arrest of human leukemia U937 cells.
Biomed Pharmacother 62, 723-729, 2008.
23) Scott FL, Denault JB, Riedl SJ, Shin H, Renatus M, Salvesen GS. XIAP inhibits caspase-3 and -7 using two binding sites:
evolutionarily conserved mechanism of IAPs. EMBO J 24, 645-655, 2005.
24) Kimberley FC, Screaton GR. Following a TRAIL: update on a ligand and its five receptors. Cell Res 14, 359-372, 2004.
25) Thomas DA, Du C, Xu M, Wang X, Ley TJ. DFF45/ICAD can be directly processed by granzyme B during the induction of apoptosis. Immunity 12, 621-632, 2000.
26) Wolf BB, Schuler M, Echeverri F, Green DR. Caspase-3 is the primary activator of apoptotic DNA fragmentation via DNA fragmentation factor-45/inhibitor of caspase-activated DNase inactivation. J Biol Chem 274, 30651-30656, 1999.
27) Fukuda K. Apoptosis-associated cleavage of β-catenin in human colon cancer and rat hepatoma cells. Int J Biochem Cell Biol 31, 519-529, 1999.
28) Manero F, Gautier F, Gallenne T, Cauquil N, Grée D, Cartron PF, Geneste O, Grée R, Vallette FM, Juin P. The small organic compound HA14-1 prevents Bcl-2 interaction with Bax to sensitize malignant glioma cells to induction of cell death. Cancer Res 66, 2757-2764, 2006.
29) Pei XY, Dai Y, Grant S. The small-molecule Bcl-2 inhibitor HA14-1 interacts synergistically with flavopiridol to induce mitochondrial injury and apoptosis in human myeloma cells through a free radical-dependent and Jun NH2-terminal kinase-dependent mechanism. Mol Cancer Ther 3, 1513-1524, 2004.