33 책임저자:김군도
608-737, 부산시 남구 용소로 45 부경대학교 자연과학대학 미생물학과 Tel: 051-629-5618, Fax: 051-629-5619 E-mail: [email protected] 박남규
608-737, 부산시 남구 용소로 45 부경대학교 수산과학대학 생물공학과 Tel: 051-629-5867, Fax: 051-629-5863 E-mail: [email protected]
접수일:2013년 1월 28일, 1차 수정일:2013년 1월 31일, 2차 수정일:2013년 2월 5일, 게재승인일:2013년 2월 7일
*The first two authors equally contributed to this work.
Correspondence to:Gun-Do Kim
Department of Microbiology, College of Natural Sciences, Pukyong National University, 45, Yongso-ro, Nam-gu, Busan 608-737, Korea Tel: +82-51-629-5618, Fax: +82-51-629-5619
E-mail: [email protected]
Co-corresponding author:Nam Gyu Park
Department of Biotechnology, College of Fisheries Sciences, Pukyong National University, 45, Yongso-ro, Nam-gu, Busan 608-737, Korea Tel: +82-51-629-5867, Fax: +82-51-629-5863
E-mail: [email protected]
Anticancer Mechanism of AP1, Abalone Peptide by Cell Cycle Arrest in Human Gastric Cancer Cells
Cho Won Kim1*, Hye-Jin Go2*, Jung-Kil Seo3, Nam Gyu Park2 and Gun-Do Kim1
1Department of Microbiology, College of Natural Sciences, 2Department of Biotechnology, College of Fisheries Sciences, Pukyong National University, Busan 608-737, 3Department of Food Science & Biotechnology, College of Ocean Science
and Engineering, Kunsan National University, Gunsan 573-701, Korea
Gastric cancer is one of the most common malignant tumors and the second cause of cancer related death in the world. Here, we evaluated the effects of abalone peptide (AP1) on cellular mediators involved in cell cycle arrest in gastric cancer cells. AGS and MKN-28 cell lines were used as human gastric cancer cells and the mechanism study showed that AP1 decreased cell population by cell cycle arrest and then led to apoptotic cell death of MKN-28 and AGS cells. Cell cycles were analyzed by flow cytometry and western blotting. Treatment with AP1 arrested the cell cycle in G1/S phase. AP1 decreased expressions of not only CDK2 and CDK6 but also Cyclin D1 and Cyclin D3. Along with those decrease, expressions of phospho-Rb, E2F1, phospho-chk1 and Cdc25A also decreased as well. Activation of caspases-3, -6 and -7 was also observed. In conclusion, the findings in this study suggested that AP1 have potential anticancer efficacy via cell cycle arrest in G1/S phase and induction of apoptosis. Its potential role to be a candidate of anticancer agent is worth being further investigated. (Cancer Prev Res 18, 33-40, 2013) Key Words: Abalone peptide, Gastric cancer cells, MKN-28, AGS, Cell cycle, Apoptosis
INTRODUCTION
Gastric cancer is one of the most common malignant tumors and the second cause of cancer related death in the world.1) Despite recent advances in targeted therapy and improved un- derstanding of the biology, progress in the treatment of gastric cancer has been limited.2) Therefore, it continuously incur to be a major unresolved health problem and new approaches for the management of gastric cancer are needed.3) For patients with unrespectable advanced or recurrent gastric cancers, the
main therapeutic option is systemic chemotherapy. Although a series of trials have produced evidence that chemotherapy can prolong the survival of patients with advanced gastric cancers, a globally accepted standard chemotherapy and the optimal regimen have not yet been determined.3) Furthermore, systemic chemotherapy usually results in resistance arising in gastric can- cer cells, which is associated with poor prognosis. Therefore, novel targets and therapeutic agents from natural resources are necessary for the treatment of gastric cancers.
Abalones are edible sea snails, marine gastropod mollusks in the family of Haliotidae and its genus is Haliotis.4) Abalones
are largely cultivated and used as valuable food source in East Asian countries. In Korea, not only the protein rich body part of abalone but also the viscera portion is taken in the form of sashimi or pickle and used as stamina food from ancient times. Although the nutritional composition of abalone visceral extract have not yet been identified in depth, since abalone lives on brown algae like Ecklonia, Laminaria and Undaria for cultivation,5) it is regarded that the visceral portion of abalone may contain concentrated nutritional components derived from sea weeds. Besides, it is well known that polysaccharides and glycoproteins of the brown algae possess potential immune- stimulant, anti-tumoral and anti-viral activity.6)
The regulation of cell cycle plays an important role in pro- liferation and survival of human gastric cancer cells. The eu- karyotic cell cycle is regulated by signal transduction pathways mediated by a series of cell cycle regulators, including cyclins, cyclin dependent kinases (CDKs) and CDK inhibitors (CDKIs).
Cyclin molecules regulate the progression of each phase of cell cycle by associating with its corresponding specific CDKs.
Cyclin K activates CDK6, which help maintaining and pro- gressing through the early G1 phase of the cell cycle. Cyclin E and CDK2 proteins play a role in the transition from the G1 to S phase of the cell cycle.7) Increased expression of CDKs and cyclins have been observed in the most of cancer cells.
Especially, the loss of cell cycle control in G1 has been im- plicated in tumor development and proliferation.8,9) Therefore, induction of cell cycle arrest by chemo-preventive drugs could be an effective approach to treat uncontrolled cell proliferation and survival of tumor cells. Recently, many studies have shown that induction of cell cycle arrest via the inhibition of CDK activity is one of the promising strategies for the discovery and design of novel anticancer agents.8,10)
In this study, we investigated the molecular mechanism of abalone peptide (AP1) against human gastric cancer cells. We demonstrated that AP1 exhibited antitumor activity by induc- ing cell cycle arrest in the phase of G1/S and apoptosis.
Antitumor effects of AP1 were characterized by down-regu- lation of CDK2, CDK6, Cyclin D1, Cyclin D3, phospho-Rb (Ser807/811) and E2F1. We also found that AP1 induced apoptosis through the activation of caspases. Based on the re- sults, the potentiality of AP1 to be a candidate of anticancer agent is worth being further investigated.
MATERIALS AND METHODS 1. Design of peptide
Analogue peptide was designed from the primary structure of a novel antimicrobial peptide purified from the gill of the abalone, Haliotis discus, as a parent peptide (unpublished data).
The parent peptide is composed of 47 amino acids containing richly some specific amino acids such as Ala and Lys and showed high sequence homology with the N-terminus of his- tone H1. It showed potent antimicrobial activity against Gram- positive and negative bacteria without hemolytic activity. To design the analogue peptide, three basical points of views in- cluding αα-helical structure having the optimal amphipathic character, C-terminal amidation increasing net positive charge and stability, and small size reducing cost were firstly considered.
To select the optimal αα-helical region in the parent peptide, the secondary structure was predicted by the GOR method (http://
www.expasy.org/tools/). In addition, to optimize the amphipathcity and adjust the sequence length, a Schiffer Edmunson plot was gen- erated by the DNASIS v2.5 demo program (Hitachi Software Engineering Co. Ltd., Tokyo, Japan). The theoretical isoelectric point and molecular mass were estimated with ExPASy (http://www.expa- sy.ch/tools/peptide-mass.html). Hydrophobicity and hydropho- bic moment were obtained from the DNASIS v2.5 demo pro- gram (Hitachi Software Engineering Co. Ltd.).
2. Peptide synthesis
The designed peptides were synthesized by the solid-phase synthesis method on an ASP48S Pepsynthesizer (Peptron Inc, Daejeon, Korea) using 9-flurorenylmethoxycarbonyl (Fmoc)- polypeptide active ester chemistry. Two forms for the peptides, one amidated (NH2) and the other with a free carboxy terminus (COOH), were synthesized with Fmoc-NH-SAL resin or Fmoc-resin. The synthetic peptides were purified on a Vydac Everest C18 column (10 μm, 300 Å, 22×250 mm) with a water-acetonitrile (ACN) linear gradient of 3∼40% ACN in 0.1% trifluoroacetic acid (TFA). Molecular weights and purities (>95%) of the synthetic peptides were confirmed using an LC/MS (Agilent HP1100 series, Agilent Technologies, Wald- bronn, Germany) and the reverse phase HPLC using a CapCell-Pak C18 reversed-phase column (5 μm, 300 Å, 4.6×
250 mm, Shiseido, Tokyo, Japan).
3. Physical parameter of the designed peptide (AP1)
The physical properties of the designed peptide were meas- ured by several different methods as mentioned above. The de- signed peptide shows an αα-helical amphipatic structure con- sisting of 14 amino acid residues (1466.9 Da) with 4 hydo- phobic residues and 5 hydrophilic residues containing 3 basic residues and contained a Trp residue for fluorescence work.
This peptide have a positive net charge (+5), high hydro- phobicity and hydrophobic moment. The molecular weights and the calculated formula of the synthetic peptide were as fol- lows: 1466.9 Da and C71H123N19O14. The peptide was dis- solved in acidified water (0.01% acetic acid) to make stock sol- utions and then further diluted in the same solution for test.
4. Cell culture
Human gastric cancer cell lines, MKN-28 and AGS, were obtained from the Korean Cell Line Bank (Seoul, Korea) and human embryonic kidney cell line, HEK293, was from American Tissue Culture Collection (ATCC, Manassas, VA, USA). RPMI 1640 medium, Deulbecco's modified Eagle's me- dium with high glucose (DMEM/high glucose), fetal bovine se- rum (FBS) and penicillin-streptomycin were purchased from Cellgro Mediatech Inc., (Manassas, VA, USA). MKN-28 and AGS cells were cultured in RPMI 1640 medium supplemented with 10% heat-inactivated FBS, 100 units/ml penicillin, and 0.1 mg/ml streptomycin. HEK293 cells were cultured in Dul- becco's modified Eagle's medium with high glucose (DMEM/
high glucose), 10% heat-inactivated FBS and 1% 100 U/ml penicillin and 100 μg/ml streptomycin. The cells were main- tained under standard cell culture conditions at 37oC and 5%
CO2 in a humid environment.
5. Cell viability assay
The cell viability of AP1 treated cancer cells and normal cells were determined using WST-1 assay. Briefly, cells (1×104 cells/
well) were seeded in 96-well plates and cultured for 24 h, followed by various concentrations of AP1 (2∼14 μM) treatment for 24 h. 10 μl of WST-1 solution was added and incubated for ad- ditional 3 h. The absorbance of the reaction was measured us- ing ELISA reader (Molecular Devices, Sunnyvale, CA, USA) at 460 nm and inhibitory rates were calculated.
6. Cell cycle analysis by flow cytometry
MKN-28 and AGS cells were plated in 60 mm culture dishes. After an overnight incubation, the cells were treated with indicated concentrations of the agent (4 μM of AP1) and the control cells were untreated with the agent. After in- cubation for 24 h, the cells were washed twice with cold phos- phate-buffered saline (PBS), detached with 0.25% tryp- sin-EDTA, and pelleted. The pellets were suspended in cold PBS and the cells were fixed in a final concentration of 70%
ethanol for overnight at 4oC, and then resuspended in PBS added 0.2 mg/ml (final concentration) RNase A for 1 h at 37oC. DNA was stained with 40 μg/ml (final concentration) propidium iodide for 30 min and the cells were examined by FACS Calibur (Becton Dickinson, Mountain View, CA, USA).
7. Western blot analysis
For Western blotting, the MKN-28 and AGS cells were cul- tured as mentioned above and treated with 8, 10 and 12 μM of AP1, and then harvested after 24 h. The cells were collected by centrifugation, lysed in ice-cold lysis buffer [50 mM Tris-Cl (pH 7.5), 150 mM NaCl, 1 mM DTT, 0.5% NP-40, 1%
Triton X-100, 1% deoxycholate, 0.1% SDS and proteinase in- hibitors (PMSF, EDTA, aprotinin, leupeptin, prostatin A)]
(Intron Biotechnology, Seongnam, Korea). After incubation on ice for 30 min, the insoluble materials were removed by cen- trifugation at 14,000 rpm for 20 min at 4oC. The protein con- tent of the cell lysates were determined by a Protein Quantifi- cation Kit (CBB Solution) with bovine serum albumen (BSA) as standard. An aliquot from each sample was boiled with sam- ple buffer for 5 min, and then resolved by 12% SDS-poly- acrylamide gel-eletrophoresis (SDS-PAGE). The proteins were electrotransferred to a nitrocellulose membrane and then blo- cked in PBST buffer (135 mM sodium chloride, 2.7 mM potas- sium chloride, 4.3 mM sodium phosphate, 1.4 mM potassium dihydrogen phosphate, 0.5% Tween-20) containing 5% skim milk for overnight at 4oC. The blots were probed with the pri- mary antibodies (anti-CDK2, anti-CDK6, anti-E2F1, anti-phos- pho-Rb, anti-β-actin, anti-GAPDH, anti-Cyclin D1, anti- Cyclin D3, anti-Cdc25A, anti-phospho-Chk1) and after that, washed three times in PBST, followed by incubation for 1 h with horseradish peroxidase-coupled anti-rabbit IgG or an- ti-mouse IgG as second antibodies (Cell Signaling Technology Inc., Danvers, MA, USA). The blots were washed in PBST and
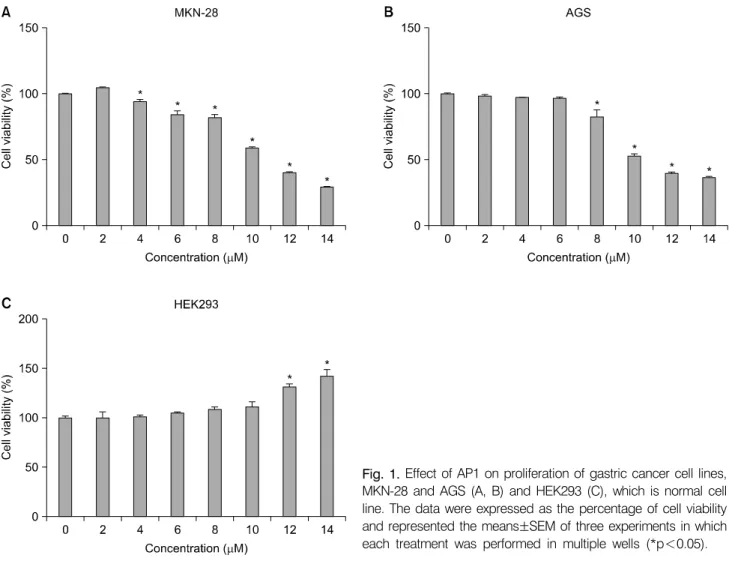
Fig. 1. Effect of AP1 on proliferation of gastric cancer cell lines, MKN-28 and AGS (A, B) and HEK293 (C), which is normal cell line. The data were expressed as the percentage of cell viability and represented the means±SEM of three experiments in which each treatment was performed in multiple wells (*p<0.05).
visualized by an enhanced chemiluminescent (ECL) detection solutions.
8. Statistical analysis
The Prism 6.0 Software (Graphpad, San Diego, CA, USA) for window was used to determine the statistical significance of differences between values for various experimental and con- trol group. Determinations were performed in triplicate and re- sults are expressed as mean values±standard error of the mean (SEM) and analyzed using one-way ANOVA.
RESULTS
1. Effect of AP1 on cell proliferation
In order to investigate the growth inhibitory properties of AP1, we performed WST-1 assay by treating AP1 on human gastric cancer cells versus non-cancerous HEK293 cells. MKN-28, AGS
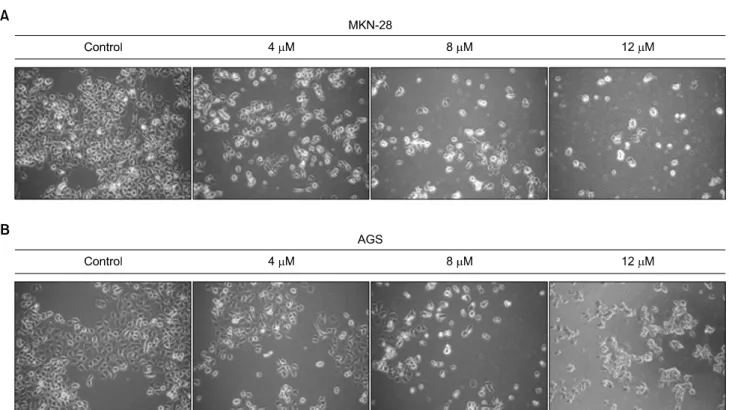
and HEK293 cells were seeded in 96-well culture plates in- dependently (1×104 cells/well). After incubation for 24 h, the cells were treated with various concentrations of AP1 or only medium as control. After incubation for 24 h, the cells were subjected to WST-1 assay (Fig. 1). The viability of MKN-28 and AGS cells was inhibited by the treatment of AP1 (2∼14 μM) and cell death was induced as dose increased (Fig. 1A, 1B). However, there was no inhibitory effect on cell prolifera- tion of non-cancerous HEK293 cells suggesting AP1 has no cytotoxicity on non-cancerous cells (Fig. 1C). MKN-28 and AGS cells treated with or without AP1 were visualized under inverted microscope (×100) to investigate its morphological changes (Fig. 2). It is confirmed that with the higher concen- tration of AP1, the more cells died and floated.
2. Effects of AP1 on cell cycle progression We analyzed the cell cycle to characterize the inhibition of
Fig. 2. Morphological changes of MKN-28 (A) and AGS (B) cells were analyzed as increase of AP1 concentration. They were treated for 24 h and the pictures are captured by a phase contrast microscope (magnification, ×200).
gastric cancer cell growth induced by AP1 and to relate this to cell cycle progression. Compared with control, AP1 treat- ment resulted accumulation of MKN-28 and AGS cells in the G1/S phase of the cell cycle (Fig. 3). For treatment at 4 μM, the accumulation levels of cells in the G1/S phase relative to control cells were increased from 2.13 to 31.53% in MKN-28 and from 3.32 to 14.37% in AGS cells.
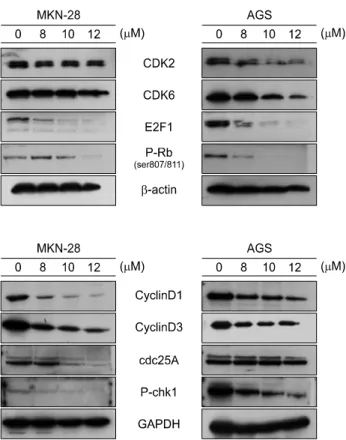
As shown in Fig. 4, the results revealed that when the cells were treated with 8, 10, 12 μM of AP1 for 24 h, it leads to changes of the expression of proteins which are involved in cell cycle progression. Treatment of MKN-28 and AGS cells with different concentrations of AP1 for 24 h led to decrease in the CDK2, CDK6, phospho-Rb and E2F1 protein expression levels. Also, cyclin D1 and D3, function during G1/S, Cdc25A and phospho-Chk1 were also decreased was reduced by AP1, which means by suppressing progression through the G1/S phase.
3. Effects of AP1 on expression of apoptosis- related proteins in gastric cancer cells
To investigate whether AP1 induced apoptosis was asso- ciated with the caspase family proteins. The expressions of
cleaved caspase-3, cleaved caspase-6 and cleaved caspase-7 were examined after 24 h of AP1 treatment. Especially, the levels of the cleaved caspase-3 were significantly increased, indicating the activation of the caspase (Fig. 5).
DISCUSSION
In this study, we investigated the inhibitory effect of AP1 on proliferation of human gastric cancer cells and its possible action mechanism. The study demonstrated that AP1 inhibited proliferation of gastric cancer MKN-28 and AGS cells by cell cycle arrest and caspases-mediated apoptosis.
Inhibition of cell cycle progression is an important step in cancer therapy because abnormal cell proliferation is observed in cancer cells.11) In order to elucidate the mechanism for the effects of AP1 on gastric cancer cells, the cell cycle distribution was examined using FACS analysis. When MKN-28 and AGS cells were treated with AP1, the number of treated cells in G1
phase was higher than the control cells with the decrease of S phase population (Fig. 3). These results suggest that AP1 arrests the cell cycle progression of both cells at the G1 phase.
In addition, the expression of cell cycle regulatory proteins were
Fig. 3. Effects of AP1 on the cell cycle in human gastric cancer cells. Gastric cancer MKN-28 (A) and AGS (B) cells were treated with AP1 (4 μM) for 24 h. The distribution histogram of the nuclear DNA detected by flow cytometry was indicated the arrest of cell cycle.
examined by Western blot analysis (Fig. 4).
Regulation of proteins that mediate critical events of the cell cycle may be a useful anti-tumor target.12) Cyclins and cy- clin-dependent kinases (Cdks) complexes are regulators of cell cycle progression where cyclins are regulatory units and Cdks are catalytic units, and these proteins are regulated by in- hibitory proteins.13) The retinoblastoma (Rb) protein is a key regulator of the G1/S transition in the cell cycle and it is asso- ciated with CDKs such as CDK2 and CDK6, which are im- portant cyclin-dependent kinases that regulate G1 progression and G1/S transition, respectively. Treatment of MKN-28 and AGS cells with different concentrations of AP1 decrease in the phospho-Rb protein expression. Phosphorylated Rb controls cell cycle by binding to and inhibiting the E2F transcription factor that regulates the G1/S cell cycle checkpoint.14) Treatment of
MKN-28 and AGS cells with different concentrations of AP1 for 24 h led to decrease in the E2F1 protein expression level.
Cdc25 protein phosphatase family plays a critical role in acti- vating cyclin-dependent kinases (CDKs) via dephosphorylation of conserved Thr14/Tyr15 inhibitory phosphorylation sites. The primary substrate of Cdc25A is CDK2, which, when active, al- lows progression through the G1/S and intra-S checkpoints.15) During normal cell cycle progression, elevated c-Myc and E2F transcription factor levels lead to increased Cdc25A expre- ssion.16) When conditions are favorable for DNA synthesis, Cdc25A and CDK2 form an activation loop, wherein each acti- vates the other enzyme.15) DNA damage, on the other hand, leads to multi-site phosphorylation at inhibitory sites (Ser123, Ser177, Ser278, Ser292, and Thr506) by Chk1, which result in 14-3-3 binding and ubiquitin-mediated degradation.17)
Fig. 4. The expression levels of cell cycle regulatory proteins were examined by Western blot analysis with indicated antibodies. The cells were treated with 8, 10, 12 μM of AP1 for 24 h.
Fig. 5. Expression of apoptosis related caspases was deter- mined by Western blot analysis after MKN-28 and AGS cells were exposed to various concentrations (8, 10 and 12 μM) of AP1 for 24 h.
Treatment with AP1 slightly decreased the expression of Cdc25A and phosphorylated Chk1, which is activated form.
The effect of AP1 on cell cycle arrest might be through multi- ple direct and indirect mechanisms, and the primary target identification is very important for small molecular anticancer compound development and its further application.
Many studies have shown that the outcome of cell cycle ar- rest could be lead to cell death by apoptosis.18,19) To demon- strate the rapid response of the MKN-28 and AGS cells, we evaluated the AP1 induced apoptosis. Apoptosis is a type of cell death regulated in an orderly way by a series of signal cas- cades under certain situations. It plays an essential role in regu- lating growth, development and clearing redundant or abnor- mal cells in organisms. The induction and execution of apopto- sis required the cooperation of a series of molecules including signal molecules, receptors, enzymes and gene regulating pro- teins. Among them, the caspase cascade signaling system is vi- tal in the process of apoptosis.20) The most prevalent caspase in the cell is caspase-3. It is the one ultimately responsible for
the majority of the apoptotic effects, although it is supported by two others, caspase-6 and caspase-7. Together, these three executioner caspases presumably cause the apoptotic phenotype by cleavage or degradation of several important substrates.21) We analyzed the proteins involved in apoptosis, and found that caspase-3, caspase-6 and caspase-7, the key molecular in apop- tosis, was activated. Thus, apoptotic changes we observed in the cells might be attributed to the fate of cell cycle arrest.
CONCLUSION
In conclusion, we were obliged to evaluate the possibility of cell cycle to reduce proliferation for MKN-28 and AGS cells by AP1. For this reason, cell cycle analysis was conducted using flow cytometry and Western blot for cell cycle regulatory pro- teins. We demonstrated that AP1 arrested the cell cycle at the G1/S phase, inhibited cell proliferation in human gastric cancer cell lines, MKN-28 and AGS. We also established the mecha- nism of action of this cell cycle arrest and apoptosis. Since de- regulation of both the cell cycle is closely linked to cancer pro- gression,22) our study might provide to serve as potential candi- dates of cell cycle arrest to be used in the treatment or pre- vention of human gastric cancers.
ACKNOWLEDGEMENT
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (grant number: No. 2010-0006123).
REFERENCES
1) Park JM, Xian XS, Choi MG, Park H, Cho YK, Lee IS, Kim SW, Chung IS. Antiproliferative mechanism of a cannabinoid agonist by cell cycle arrest in human gastric cancer cells. J Cell Biochem 112, 1192-1205, 2011.
2) Shah MA, Kelsen DP. Gastric cancer: a primer on the epidemiology and biology of the disease and an overview of the medical management of advanced disease. J Natl Compr Cancer Netw 8, 437-447, 2010.
3) Lou LH, Jing DD, Lai YX, Lu YY, Li JK, Wu K. 15-PGDH is reduced and induces apoptosis and cell cycle arrest in gastric carcinoma. World J Gastroenterol 18, 1028-1037, 2012.
4) Lin A, Meyers MA. Growth and structure in abalone shell.
Material Sci Engineering A 390, 27-41, 2005.
5) Hwang E, Baek J, Park C. The mass cultivation of Ecklonia stolonifera Okmura as a summer feed for the abalone industry in Korea. J Appl Phycol 21, 585-590, 2009.
6) Smit AJ. Medicinal and pharamaceutical uses of seaweed natural products. A review. J Appl Phycol 16, 245-262, 2004.
7) Massague J. G1 cell-cycle control and cancer. Nature 432, 298-306, 2004.
8) Malumbres N, Pevarello P, Barbacid M, Bischoff JR. Cdk inhibitors in cancer therapy: What is next? Trends Pharmacol Sci 29, 16-21, 2008.
9) Musgrove EA, Caldon CE, Barraclough J, Stone A. Sutherland RL. Cyclin D as a therapeutic target in cancer. Nat Rev Cancer 11, 558-572, 2011.
10) Tong WG, Chen R, Plunkett W, Siegel D, Sinha R, Harvey RD, Badros AZ, Popplewell L, Coutre S, Fox JA, Mahadocon K, Chen TL, Kegley P, Hoch U, Wierda WG. Phase I and pharmacologic study of SNS-032, a potent and selective Cdk2, 7, and 9 inhibitor, in patients with advanced chronic lym- phocytic leukemia and multiple myeloma. J Clin Oncol 28, 3015-3022, 2010.
11) Lopez-Saez J, De La Torre C, Pincheira J, Gimenez-Martin G. Cell proliferation and cancer. Histol Histopathol 13, 1197- 1214, 1998.
12) Stewart ZA, Westfall MD, Pietenpol JA. Cell-cycle dysre-
gulation and anticancer therapy. Trends Pharmacol Sci 24, 139- 145, 2003.
13) Chilampalli C, Guillermo R, Kaushik RS, Young A, Chan- drasekher G, Fahmy H, Dwivedi C. Honokiol, a chemopre- ventive agent against skin cancer, induces cell cycle arrest and apoptosis in human epidermoid A431 cells. Exp Biol Med 236, 1351-1359, 2011.
14) DeGregori J, Kowalik T, Nevins JR. Cellular targets for activation by the E2F1 transcription factor include DNA synthesis- and G1/S-regulatory genes. Mol Cell Biol 15, 4215- 4224, 1995.
15) Hoffmann I, Draetta G, Karsenti E. Activation of the pho- sphatase activity of human Cdc25A by a Cdc2-cyclin E depen- dent phosphorylation at the G1/S transition. EMBO J 13, 4302-4310, 1994.
16) Vigo E, Müller H, Prosperini E, Hateboer G, Cartwright P, Moroni MC, Helin K. Cdc25A phosphatase is a target of E2F and is required for efficient E2F-induced S phase. Mol Cell Biol 19, 6379-6395, 1999.
17) Chen MS, Ryan CE, Piwnica-Worms H. Chk1 kinase negatively regulates mitotic function of Cdc25A phosphatase through 14-3-3 binding. Mol Cell Biol 23, 7488-7497, 2003.
18) Ye M, Liu J, Lu Z, Zhao Y, Liu S, Li L, Tan M, Weng X, Li W, Cao Y. Grifolin, a, potential antitumor natural product from the mushroom albatrellus confluens, inhibits tumor cell growth by inducing apoptosis in vitro. FEBS Lett 579, 3437- 3443, 2005.
19) Krishnan A, Harigaran R, Nair SA, Pillai MR. Fluoxetine mediates G0/G1 arrest by inducing functional inhibition of cyclin dependent kinase subunit (CKS)1. Bioc Pharmacol 75, 1924-1934, 2008.
20) Launay S, Hermine O, Fontenay M, Kroemer G, Solary E, Garrido C. Vital functions for lethal caspases. Oncogene 24, 5137-5148, 2005.
21) Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 1, 43-50, 1998.
22) Davidson G, Niehrs C. Emerging links between CDK cell cycle regulators and Wnt signaling. Trends Cell Biol 20, 453- 460, 2010.