246
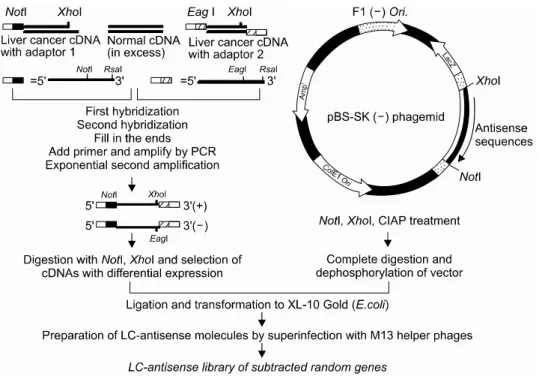
Identification of Genes Involved in Liver Cancer Cell Grow th U sing an A ntisense Library of Phage Genomic DN A
Yun Han Lee, M.S.
1,3, Young Ho Kim, Ph.D.
3and Jong Gu Park, Ph.D.
1,21
전체 글
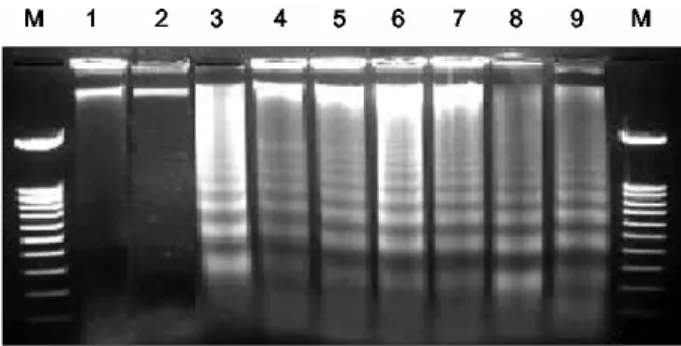
1
수치



관련 문서
Arabic Scripted Language Identification System in order to identify the language of the Arabic script by using PCA and KNN, here, the number of PCA features were optimized
In this study, the expression profiles of miRNAs were compared and analyzed for establishment of miRNAs related cancer cell growth inhibition in normal human oral
The expression of reporter genes driven by the ZAT1 promoter was observed in the tissues undergoing maturation such as border cell of root cap, endodermis,
We determined the nucleotide sequences of the mitochondrial DNA (mtDNA) control region using cloning and sequencing, and obtained the complete sequence from the cattle bones
Objective: The purpose of this study was to analyze recent trend in incidence of basal cell carcinoma and squamous cell carcinoma in patients from the Gwangju City
Embedded commands: database commands are embedded in a general-purpose programming language.. Library of database functions: available to the host language for
The 5-day-old seedlings overexpressing the AtTX12 -like genes originated from crops of Brassicaceae were used for the extraction of the total RNA from which cDNA was
In the present study, consistent to that report, up-regulation of VEGF mRNA was observed in the colon cancer cell with the acquired resistance to 5-FU and this