Cytotoxic Evaluation of the Essential Oils from Korean Native Plant on Human Skin and Lung Cells
13
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
수치
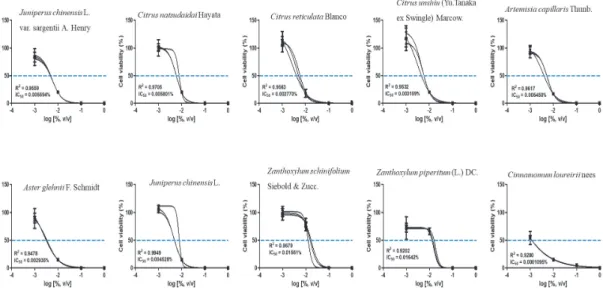
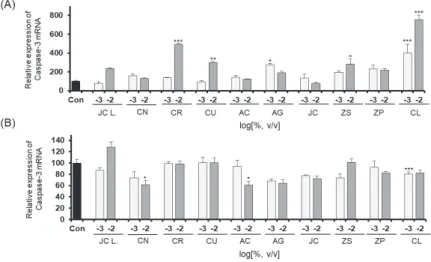

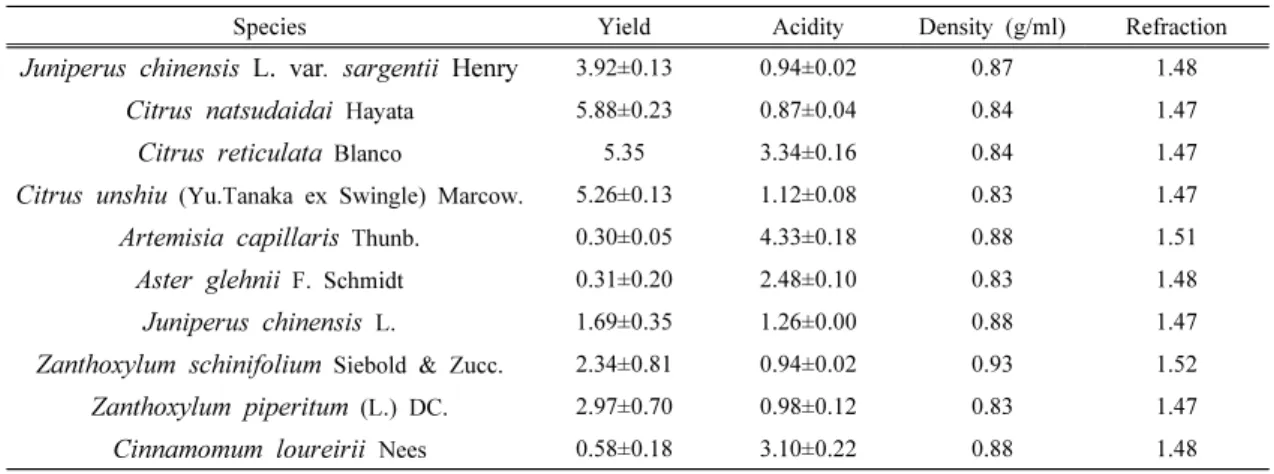
관련 문서
The expression levels of apoptotic related proteins and caspase dependent proteins by the ostreolysin purified from P.ostreatus on cell viability in FaDu human
Learning Korean is essential to understand Korean culture and live in Korea with fewer obstacles. Fortunately, there are many institutions in Korea offering Korean language
This paper discovered the Korean native Chinese characters which were flowed in Japanese, discussing the necessity of setting up the standards for judging
The strong role of Korean government from the days of economic development in the 1960s became the target of the regulatory reform from 1990s, the government reform and
For the data collection, we generated our gene expression profiles of Illumina platform and our gene expression profile demonstrated higher expression and connectivity than
william James Hall, M.D., Medical Missionary to the Slums of New York; Pioneer Mission of Korean Native Costumes, China Medical Journal, 1916.3; Medical Needs of Korean
In view of the essential relevance of mission in history (and not only in the history of religion), on the one hand, and in theology (and not only in missiology),
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
