Author contributions: I.H.B., C.S.L., and D.Y.K. designed the study. I.H.B., S.H.L., S.O., H.W.C., P.A.M., J.W.Y., J.Y.K., and E.S.L. performed the experiments.
I.H.B., T.R.L., C.S.L., and D.Y.K. analyzed the data and contributed to writ- ing the manuscript. All authors read and approved the final version of the manuscript.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Copyright © Korean J Physiol Pharmacol, pISSN 1226-4512, eISSN 2093-3827
INTRODUCTION
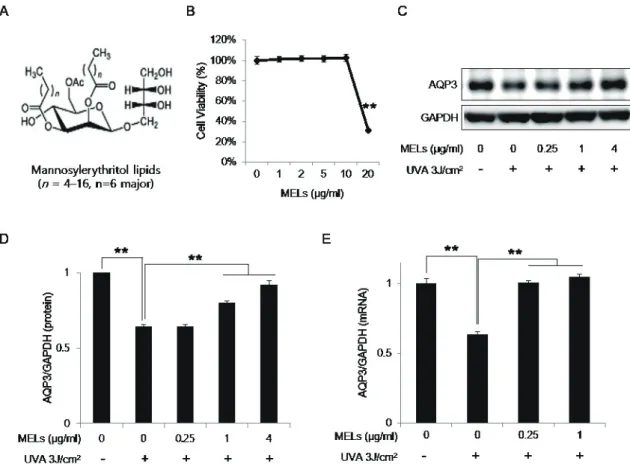
Mannosylerythritol lipids (MELs) are extracellular glycolipids that are produced from different vegetable oil substrates by the yeast strains of genus Pseudozyma [1]. MELs comprise a hydro- philic headgroup containing 4-O--D-mannopyranosyl eryth-
ritol or 1-O--D-mannopyranosyl erythritol and fatty acid-con- taining hydrophobic chains [1]. MELs were originally developed as a highly biodegradable biosurfactant [2], but were subsequently found to have additional biological properties such as anti- inflammation and apoptosis of melanoma cells [3-7]. As the pos- sible applications of MELs expanded, some researchers focused
Original Article
Mannosylerythritol lipids ameliorate ultraviolet A-induced aquaporin-3 downregulation by suppressing c-Jun N-terminal kinase phosphorylation in cultured human keratinocytes
Il-Hong Bae 1,3 , Sung Hoon Lee 1 , Soojung Oh 1 , Hyeongwon Choi 1 , Paulo A. Marinho 1 , Jae Won Yoo 1 , Jae Young Ko 1 , Eun-Soo Lee 1 , Tae Ryong Lee 1 , Chang Seok Lee 2, *, and Dae-Yong Kim 3, *
1