350 책임저자:최영현, 614-052, 부산시 진구 양정동 산 45번지
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4036 E-mail: [email protected]
접수일:2009년 12월 8일, 게재승인일:2009년 12월 15일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, College of Oriental Medicine, Dongeui University, San 45, Yangjung-dong, Busanjin-gu, Busan 614-714, Korea Tel: +82-51-850-7413, Fax: +82-51-853-4036
E-mail: [email protected]
흑마늘 추출물에 의한 AGS 인체 위암세포의 Apoptosis 유도에 관한 연구
1동의대학교 한의과대학 생화학교실, 대학원 바이오물질제어학과 및 블루바이오 소재개발센터,
2국립원예특작과학원 채소과, 3부산대학교 생명자원과학대학 원예생명과학과, 4경남도립남해대학
호텔조리제빵과, 5인제대학교 식품생명과학부
신동역1ㆍ윤무경2ㆍ최영환3ㆍ권오천4ㆍ김정인5ㆍ최영현1
Apoptosis Induction by Aged Black Garlic Extracts in AGS Human Gastric Carcinoma Cells
Dong Yeok Shin1, Moo Kyoung Yoom2, Young-Whan Choi3, Oh-Cheon Gweon4, Jung-In Kim5 and Yung Hyun Choi1
1Department of Biochemistry, College of Oriental Medicine, Department of Biomaterial Control, Graduate School and Blue-Bio Industry Regional Innovation Center, Dongeui University, Busan 614-714, 2Vegetable Research Division, National Institute of Horticulture & Herbal Science, Suwon 440-706, 3Department of Horticultural Bioscience, College of Natural Resource and Life Sciences, Pusan National University, Miryang 627-706, 4Department of Hotel Culinary Arts & Bakery, Gyeongnam Provincial Namhae College, Namhae 668-801, 5School of Food and Life Science, Inje University, Gimhae 621-749, Korea
Garlic is generally used as a therapeutic reagent against various diseases, and many studies have indicated that garlic and its derivatives can reduce the risk of various types of human cancer. In this study, the potential of water extract of raw garlic (RG) and water extract of aged black garlic (ABG) to induce apoptosis in human gastric carcinoma AGS cells was investigated. Treatment of AGS cells to ABG, but not RG, resulted in the inhibition of cell growth and viability and the induction of apoptosis, which could be proved by MTT assay, DAPI staining and flow cytometry analysis. The increase in apoptosis that was induced by ABG was correlated with down-regulation of anti-apoptotic Bcl-2 and Bcl-xL expression, up-regulation of pro-apoptotic Bax, Fas and Fas ligand, and a decrease in the mitochondrial membrane potential. ABG treatment induced the proteolytic activation of caspases and a concomitant degradation of poly (ADP-ribose)-polymerase. Cleavage of Bid and a down-regulation of the inhibitor of apoptosis family proteins were also observed in ABG-treated AGS cells. Taken together, the data from this study indicates that ABG induces the apoptosis of AGS cells through a signaling cascade of death receptor-mediated extrinsic and mitochondria-mediated intrinsic caspase pathways. (Cancer Prev Res 14, 350-358, 2009)
Key Words: Aged black garlic, AGS, Apoptosis, Caspases
서 론
현재 암 치료를 위해서 사용하고 있는 항암제들은 암
의 종류에 따라 효능이 다를 뿐만 아니라 치료 후의 경 과나 부작용에서 많은 문제점을 가지고 있기 때문에 치 료효능이 뛰어나면서 부작용이 거의 없는 새로운 물질 의 개발이 필요한 실정이다. 이러한 필요성에 의해서 많
은 연구자들은 천연물들에 관심을 가지게 되었고, 일부 천연물에서 항암 및 면역조절 효과가 뛰어나다는 사실 이 밝혀지고 있다. 마늘(garlic, Allium sativum)은 오랫동안 식용뿐만 아니라 건강 기능성 식품으로 많은 관심을 모 으고 있는 식품의 하나로서, 마늘 성분 중 allicin은 alliin (S-allyl-L-cysteine sulfoxide)이 alliinase에 의한 촉매작용의 산물에 의한 것이다. Allicin은 상온에서 매우 불안정해서 여러 가지 다른 allyl sulfur 화합물로 분해되는 것으로 알 려져 있으며, 최근 이들 분해 산물들을 이용한 암예방ㆍ 항암ㆍ항염증 및 심혈관질환 조절제의 개발에 많은 연 구가 집중되고 있다.1∼4) 한편 흑마늘(aged black garlic)은 일정한 온도와 습도 하에서 숙성ㆍ발효시켜 특유의 자 극적인 냄새와 맛을 제거하고 폴리페놀함량을 증가시킨 것으로서 특히 항산화력이 매우 높은 것으로 보고되어 져 왔는데, 이는 아마도 S-allyl cysteine의 함량 증가와 연 관성이 있는 것으로 추측된다.5∼9) 따라서 흑마늘 추출물 은 다양한 질병의 예방과 치료에 효과적일 것으로 추정 되지만 현재까지 흑마늘 추출물의 생리ㆍ생화학적 기전 에 대한 연구는 상대적으로 매우 미비한 실정이다.
일반적으로 세포의 죽음은 세포의 형태적 및 생리ㆍ 생화학적 특성에 따라 크게 necrosis와 apoptosis로 구분되 며, necrosis는 생리적, 화학적인 외상에 의한 세포의 죽음 이고, apoptosis는 개체의 정상적인 발달과 분화에 관여하 며, 태아의 형태형성, 난자의 배란, 신경세포의 시냅스 형성 등과 연관된 체내 비정상적인 세포들을 제거하는 기전이다.10∼12) 특히 necrosis에 비하여 apoptosis 유발에는 세포질 및 염색질 응축, 세포막 수포화 현상, DNA 단편 화 등이 수반되는데 이러한 현상은 세포내부의 정교한 신호전달에 의해 조절된다.13) Apoptosis 유발에 관여하는 대표적인 유전자로 death-receptor 연관 유전자들, Bcl-2 및 inhibitor of apoptosis (IAP) family 인자들, caspase 효소군 등 으로 구분될 수 있으며, 이들의 양적인 발현 및 활성의 변화를 통하여 apoptosis의 유발 여부가 결정된다.14,15) 이 러한 정교한 유전자의 발현조절 하에 이루어지는 apop- tosis는 항암제 개발이나 잠재적인 항암성을 가질 수 있 는 새로운 물질들의 활성 검정 및 그들의 기전해석에 필 수적으로 이용되어지고 있다.
본 연구에서는 선행 연구를 통하여 강력한 항염증 효 능이 있을 것으로 추정되었던 흑마늘 추출물이16) 인체위 암세포의 증식에 미치는 영향을 생마늘 추출물과 비교 조사하였다. 아울러 흑마늘 추출물 처리에 의한 위암세 포의 증식억제가 apoptosis 유발과 연관성이 있는지를 확 인하였으며, apoptosis 과정에 관여하는 몇 가지 중요한 유전자들의 발현 변화를 조사하여 유의적인 결과를 얻
었기에 이를 보고하는 바이다.
재료 및 방법 1. AGS 세포배양
본 실험에 사용된 AGS 인체 위암세포는 한국생명공학 연구소에서 분양받았으며, 10% fetal bovine serum (FBS)에 1%의 penicillin 및 streptomycin (Biofluids, Rockville, MD, USA)이 포함된 RPMI-1640 배지(Gibco BRL, Grand Island, NY, USA)를 사용하여 37°C 및 5% CO2 조건 하에서 배양 하였다.
2. 마늘 추출물 처리
본 실험에 사용된 생마늘(water extract of raw garlic, RG) 및 흑마늘 열수 추출물(water extract of aged black garlic, ABG)로서 인제대학교 김정인 교수연구실에서 제공받았 다.16) 이들을 처리하기 위하여 AGS 세포를 6 well에 1×105개/ml 정도 분주하여 24시간 동안 안정화시켰다.
RG 및 ABG는 세포에 처리하기 직전 적정 농도로 배지 에 첨가하여 녹인 후 주사기용 필터 유닛(0.22μm의 pore size)을 사용하거나 1회용 펌프 필터 유닛을 사용하여 미 생물 및 불순물을 걸러낸 다음, 배지를 갈아주면서 직접 처리하였다.
3. MTT assay에 의한 세포 성장억제 조사
24시간 동안 안정화시킨 AGS 세포에 48시간 동안 RG 및 ABG를 처리한 후 배지를 제거하고 tetrazolium bromide salt (MTT, Sigma Chemical Co., St. Louis, MO, USA)를 0.5 mg/ml 농도로 희석하여 200μl씩 분주하고 3시간 동안 배양하였다. 배양이 끝난 다음 MTT 시약을 제거하고 dimethylsulfoxide (DMSO, Sigma)를 100μl씩 분주하여 well 에 생성된 formazin을 모두 녹인 후 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡광도를 측 정하였다. 측정은 모두 세 번을 하였으며, 그에 대한 평 균값과 표준 오차를 Sigma Plot 4.0 프로그램 (SPSS Ins.)으 로 구하였다.
4. AGS 세포의 형태 변화 관찰
동일 조건에서 배양된 AGS 세포의 전체적인 형태변화 는 도립 현미경(inverted microscope, Carl Zeiss, Germany)을 이용하여 200배의 배율에서 관찰하였다. Apoptosis 유발 여부를 조사하기 위한 핵의 형태적 변화 관찰을 위하여 준비된 세포를 모은 다음 37% formaldehyde 용액과 PBS 를 1:9의 비율로 섞은 fixing solution을 이용하여 상온에서
Table 1. Gene-specific primers for RT-PCR
Gene name Sequence
GAPDH
TRAIL
DR4
DR5
Fas
FasL
Bcl-2
Bcl-xL
Bax
XIAP
cIAP-1
cIAP-2
Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense Sense Antisense
5'-CGG AGT CAA CGG ATT TGG TCG TAT-3' 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3' 5'-ATG GCT ATG ATG GAG TCC AG-3' 5'-TTG TCC TGC ATC TGC TTC AGC-3' 5'-CAG AAC GTC CTG GAG CCT GTA AC-3' 5'-ATG TCC ATT GCC TGA TTC TTT GTG-3' 5'-GGG AAG AAG ATT CTC CTG AGA TGT G-3' 5'-ACA TTG TCC TCA GCC CCA GGT CG-3' 5'-TCT AAC TTG GGG TGG CTT TGT CTT C-3' 5'-GTG TCA TAC GCT TTC TTT CCA T-3' 5'-GGA TTG GGC CTG GGG ATG TTT CA-3' 5'-AGC CCA GTT TCA TTG ATC ACA AGG-3' 5'-CAG CTG CAC CTG ACG-3'
5'-ATG CAC CTA CCC AGC-3'
5'-CGG GCA TTC AGT GAC CTG AC-3' 5'-TCA GGA ACC AGC GGT TGA AG-3' 5'-ATG GAC GGG TCC GGG GAG-3' 5'-TCA GCC CAT CTT CTT CCA-3' 5'-GAA GAC CCT TGG GAA CAA CA-3' 5'-CGC CTT AGC TGC TCT CTT CAG T-3' 5'-TGA GCA TGC AGA CAC ATG C-3' 5'-TGA CGG ATG AAC TCC TGT CC-3' 5'-CAG AAT TGG CAA GAG CTG G-3' 5'-CAC TTG CAA GCT GCT CAG G-3' 10분 동안 고정하였다. 이를 4',6-diamidino-2-phenylindole
(DAPI, Sigma) 용액으로 염색 후 형광 현미경(fluorescent microscope, Carl Zeiss)을 이용하여 400배의 배율로 각 처 리 농도에 따른 핵의 형태 변화를 관찰하였다.
5. DNA flow cytometry에 의한 세포 주기의 분석
정상 배지와 RG 및 ABG가 처리한 배지에서 48시간 동안 배양시킨 AGS 세포를 핵산에 특이적으로 결합하는 형광 물질인 DNA intercalating dye propidium iodide (PI, concentration, 50μg/ml; Sigma)와 0.1 mg/ml의 RNase (Sig- ma)를 처리하여 암실 (4°C)에서 1시간 동안 염색 과정을 거쳤다. 이를 DNA flow cytometry (Becton Dickinson, San Jose, CA, USA)에 적용시켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dickinson) program을 사용하여 분석하 였다.
6. Reverse transcription-polymerase chain reac- tion (RT-PCR)에 의한 mRNA 발현의 분석
동일한 조건에서 준비된 AGS 세포를 대상으로 Trizol B (Invitrogen, Carlsbad, CA, USA)를 이용하여 total RNA를 분리하였다. 분리된 RNA를 정량한 후, ONE-STEP RT-
PCR PreMix Kit (Intron, Biotechnology, Korea)를 이용하여 2μg의 RNA에서 cDNA를 합성하였다. 이 cDNA를 tem- plate로 사용하여 관찰 대상 유전자(Table 1)를 polymerase chain reaction (PCR) 방법으로 증폭하였다. 이때 house- keeping 유전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 유전자를 internal control로 사용하였다. 각 PCR 산물들을 1% agarose gel을 이용하여 전기영동하고 ethi- dium bromide (EtBr, Sigma)로 염색한 후 ultra violet (UV) 하에서 확인하였다.
7. Western blot analysis에 의한 단백질 발현의 분석
RG 또는 ABG가 처리된 배지에서 자란 세포들을 모은 다음 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethy- lsulfonyl fluoride, 5 mM dithiothreitol]를 첨가하여 4°C에서 30분간 반응시킨 후, 14,000 rpm으로 30분간 원심분리하 여 그 상층액을 분리하였다. 상층액의 단백질 농도는 Bio-Rad 단백질 정량 시약(Bio-Rad, Hercules, CA, USA)과 그 사용방법에 따라 정량한 다음 동량의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들어 sodium dodecyl sulfate (SDS)-polyacrylamide gel을 이용하여 전기영동으로
Fig. 1. Inhibition of cell viability in human gastric carcinoma AGS cells after treatment with water extract of raw garlic (RG) and water extract of aged black garlic (ABG). AGS cells were seeded at 1×105/ml in a 6-well plate and treated with variable concentrations of RG and ABG for 48 h. The growth inhibition was measured by the metabolic-dye-based MTT assay.
분리하였다. 분리된 단백질을 함유한 acrylamide gel을 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이시킨 후, 적정 1차 항 체를 처리하여 상온에서 1시간 이상 또는 4°C에서 over night 시킨 다음 PBS-T로 세척하고 처리된 1차 항체에 맞 는 2차 항체를 사용하여 상온에서 1시간 정도 반응시켰 다. 다시 PBS-T로 세척하고 enhanced chemiluminoesence (ECL) 용액(Amersham Life Science Corp., Arlington Heights, IL, USA)을 적용시킨 다음 암실에서 X-ray film에 감광시 켜 특정단백질의 양을 분석하였다.
8. In vitro caspase-3, caspase-8 및 caspase- 9의 activity 측정
Caspases의 in vitro 활성 측정을 위한 colorimetric assay kits는 R&D Systems (Minneapolis, MN, USA)에서 구입하였 으며, 제시된 방법에 준하여 caspase 활성의 증감 여부를 조사하였다. 이를 위하여 정상 및 RG 또는 ABG가 처리 된 배지에서 48시간 배양된 세포를 모은 뒤 단백질을 추 출하고 정량하여 각각 100μg의 단백질을 fluorogenic peptide 기질 100μM이 함유된 extraction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1 mM EDTA, 0.2%
NP-40 and 10 mM DL-DTT] 50μl에 혼합하였으며, microtiter plate에 다시 extraction buffer에 희석하여 각 sample 당 총 volume이 100μl가 되게 하였다. 실험에 사용 된 기질은 caspase-3의 경우에는 Asp-Glu-Val-Asp (DEVD)- p-nitroaniline (pNA)이었고 caspase-8의 경우에는 Ile-Glu- Thr-Asp (IETD)-pNA이었으며, caspase-9은 Leu-Glu-His-Asp (LEHD)-pNA였다. 준비된 plate를 37°C에서 2시간동안 incubation 시킨 후 ELISA reader를 이용하여 405 nm의 흡 광도를 이용하여 반응의 정도를 측정하였다.
9. 미토콘드리아 막 전위 변화 관찰
RG 또는 ABG 처리에 의한 AGS 세포 미토콘드리아 막 전위(mitochondrial membrane potential, MMP, Δψm) 변화 의 측정을 위하여 6 well plate에 1×105개/ml 정도로 분주 하여 24시간 동안 안정화시킨 다음 흑마늘 열수 추출물 을 처리하였다. 48시간 동안 배양한 후, 세포를 PBS로 씻 어 내고 0.05% trypsin-EDTA를 처리하여 부유시킨 다음 원심분리를 하여 세포를 모았다. 이렇게 모인 세포에서 상층액을 제거한 후, lipophilic cationic probe인 5,5',6,6'-te- trachloro-1,1',3,3'-tetraethylbenzimidazolcarbocyanine iodide (JC- 1, CalBiochem)가 10μM 첨가된 배양액을 1 ml씩 분주하여 37°C에서 20분 동안 염색하였다. 염색된 세포를 nylon mesh를 이용하여 하나씩으로 분리시킨 후 DNA flow cy-
tometry (Becton Dickinson)에 적용시켜 형광반응에 따른 histogram을 ModiFit LT 프로그램으로 분석하였다.
결과 및 고찰
1. AGS 위암세포 증식에 미치는 마늘 추출물의 영향
AGS 인체 위암세포의 증식에 미치는 RG 및 ABG의 영 향을 알아보기 위하여 이들을 48시간동안 농도별로 처 리한 후 MTT assay를 이용하여 조사하였다. Fig. 1에서 나타난 바와 같이 AGS 세포는 RG 및 ABG의 처리 농도 의존적으로 증식이 억제되었으나, 특히 ABG 3 mg/ml 처 리군에서 세포의 생존율이 30% 가까이 떨어졌으며, 3.5 mg/ml이상에서는 10% 미만으로 떨어지는 것을 확인하 였다. 그러나 동일 조건에서 RG 처리군에서는 상대적으 로 AGS 위암세포에 대한 항암효과가 낮게 나타나 흑마 늘 추출물에 함유된 다양한 성분들이 강력한 항암효능 을 나타내었음을 알 수 있었다.
2. AGS 위암세포의 형태에 미치는 마늘 추출물의 영향
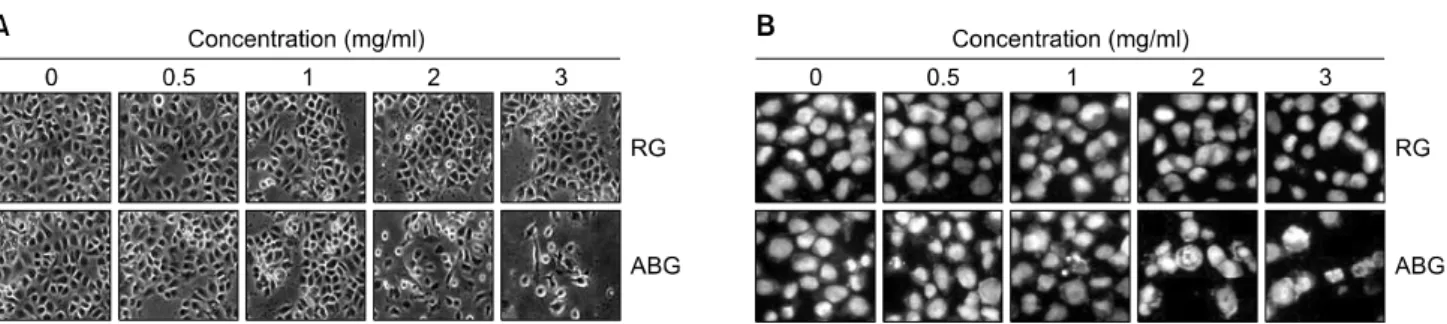
RG 및 ABG 처리에 의한 AGS 위암세포의 증식 억제 효과와 연관된 apoptosis 유발 여부 확인을 위한 암세포의 전체적인 형태 및 핵의 형태적 변화를 알아보기 위하여 RG와 ABG를 동일한 농도로 각각 48시간 동안 처리를 한 후 도립 현미경을 이용하여 처리 농도에 따른 전체적 인 세포의 형태적 변화를 관찰한 결과는 Fig. 2A에 나타 내었다. Fig. 2A에서 알 수 있듯이 RG가 처리된 AGS 세
Fig. 2. Morphological changes and nuclear chromatin condensation of AGS cells by ABG treatment. (A) AGS cells were seeded at 1×105/ml in a 6-well plate and incubated for 24 h. The cells were treated with variable concentrations of RG and ABG for 48 h. Cell morphology was visualized by an inverted microscopy. Magnification, ×200. (B) Cells were treated with RG and ABG for 48 h and then stained with DAPI solution. After 15 min incubation at room temperature, the cells were washed with PBS and nuclear morphology was photographed with a fluorescent microscope using blue filter. Magnification, ×400.
Fig. 3. Apoptosis induction of AGS cells by ABG treatment.
AGS cells were treated with variable concentrations of ABG for 48 h, then collected and stained with PI for flow cytometry analysis. The percentages of cells with hypodiploid DNA (sub- G1 phase) contents represent the fractions undergoing apop- totic DNA degradation. Data are expressed as mean of two independent experiments.
포에 비하여 ABG 처리군에서 처리 농도 증가에 따른 AGS 세포의 심한 형태적 변화가 관찰되었다. 또한, RG 및 ABG 처리에 의한 apoptosis 유발 여부 확인을 위한 핵 의 형태적 변화를 관찰하기 위하여 DAPI 염색법을 이용 하여 전형적인 apoptosis가 유발된 세포에서 관찰할 수 있 는 염색질 응축(chromatin condensation) 현상도13) ABG 처 리군에서만 관찰되었다. 이는 MTT assay의 결과와도 잘 일치되며, ABG에 의한 AGS 위암세포의 증식억제는 apoptosis 유발에 의한 것임을 알 수 있었다.
3. Apoptosis 유발에 미치는 RG 및 ABG의 영향 비교
이상의 결과에서 RG 보다는 ABG가 AGS 위암세포의 증식억제 및 apoptosis 유발 효과가 매우 높은 것으로 나
타났으므로, RG 및 ABG 처리에 따른 apoptosis 유발의 정 도를 정량적으로 비교 평가하기 위하여 동일한 조건으 로 배양된 세포들을 대상으로 PI 염색액을 이용하여 핵 을 염색한 후 DNA flow cytometry 분석을 이용하여 apoptosis 유발군에 해당되는 세포주기의 sub-G1기에 해 당되는 세포들의 빈도를 조사하였다.13) Fig. 3의 결과에 서 알 수 있듯이 RG 처리군보다 ABG 처리군에서 처리 농도의 증가에 따라 sub-G1기에 해당하는 세포의 빈도 가 매우 증가하여 상기 결과들을 잘 지지하여 주었다.
이상의 결과는 흑마늘 추출물이 생마늘 추출물에 비하 여 강력한 항암 효능을 지니며 이는 apoptosis에 의한 것 임을 재확인할 수 있었다.
4. MMP에 미치는 RG 및 ABG의 영향 비교
RG 및 ABG 처리에 따른 apoptosis 유발의 기전 해석을 위하여 apoptosis 조절에 중심이 되는 세포 내 소기관인 mitochondria의 연관성을 조사하였다. 이를 위하여 RG 및 ABG가 각각 48시간 처리된 AGS 위암세포를 대상으로 MMP (Δψm)의 소실 정도를 dual-emission fluorescent dye인 JC-1을 이용하여 염색 후 flow cytometry로 분석하였다.
RG 및 ABG가 처리되었을 경우 RG 처리군의 경우 3 mg/
ml에서 MMP의 소실 정도가 20% 정도 증가하였지만 ABG 처리군의 경우 MMP의 손실 정도가 45% 정도로 매 우 증가되었다(Fig. 4). 이러한 RG 및 ABG 처리군에서의 MMP 소실의 차이는 apoptosis 유발의 정도와 유사한 경 향성으로 나타나 특히 흑마늘 열수 추출물에 의한 apoptosis 유발에 mitochondria 기능 손상이 연관되어 있음 을 알 수 있었다.
5. Caspase 활성에 미치는 ABG의 영향
한편 cysteine-related protease인 caspase는 apoptosis 유발
Fig. 4. Loss of MMP by ABG treatment of AGS cells. (A) After being treated with ABG for 48 h, the cells were stained with JC-1 and then incubated at 37°C for 20 min, after which the mean JC-1 fluorescence intensity was detected using a flow cytometer.
(B) Data represent the mean±the SD of representative experiments performed at least three times.
에 중심적 작용을 하는 효소로서, 이들 family에 속하는 단백질들은 세포에서 핵과 mitochondria의 외막에 불활성 상태로 존재하며, Bcl-2/Bax family 발현의 변화에 따라 이 들의 활성도가 조절될 수 있다. 이들은 proenzyme 형태로 존재하다가 apoptosis 유도를 활성화 시키는 신호에 의해 활성화된 protease로 전환되어 직접 또는 간접적으로 세 포 내에 존재하는 많은 표적 단백질의 분해에 관여한다.
따라서 caspase의 활성화는 apoptosis의 유발에 대한 또다 른 증거가 될 수 있으며 많은 선행연구 등에서 검증되어
왔다.14,15) 지금까지 알려진 caspase 중 전형적인 두 가지
apoptosis 유도 기전에서 가장 중요한 역할을 할 수 있는 caspase-8, -9 및 -3의 발현 및 활성에 미치는 ABG의 영향 을 확인하기 위하여 Western blotting 및 in vitro caspase
activity assay를 통하여 caspases의 발현 변화 및 활성의 정 도를 직접 분석하였다.
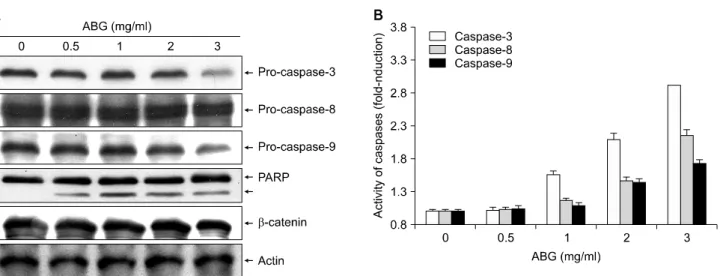
Fig. 5A에 나타낸 바와 같이 처리된 ABG의 농도가 증 가할수록 조사된 3가지 caspase의 비활성형인 pro-caspases 의 발현이 점차 감소되었음을 알 수 있었다. 따라서 이들 비활성형 발현의 감소가 활성형의 증가에 의한 것인지 를 조사한 결과 ABG의 처리 농도 의존적으로 조사된 3 가지 caspase의 활성이 모두 증가하였음을 확인할 수 있 었다(Fig. 5B). 또한 caspase-3의 표적 단백질이면서 apo- ptosis 표지 단백질의 일종인 poly (ADP-ribose) polymerase (PARP)의 발현14)을 조사한 결과, ABG 처리 농도의 증가 에 따라 단편화된 단백질의 발현이 증가되어 흑마늘 열 수 추출물 처리에 의한 apoptosis는 caspase 활성에 의한
Fig. 5. Effects of ABG on the expression and activity of caspases in AGS cells. (A) After 48 h incubation with ABG, the cells were lysed, and cellular proteins were separated by SDS-polyacrylamide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) The cells grown under same condition as (A) were collected and then lysed. Aliquots (100μg proteins) were incubated with DEVD-pNA, IETD-pNA and LEHD-pNA for caspase-3, -8 and -9 activity, respectively, at 37°C for 2 h. The released fluorescent products were measured. The data shown are means±SD of three independent experiments.
것임을 알 수 있었다.
6. Bcl-2 family의 인자들의 발현에 미치는 ABG의 영향
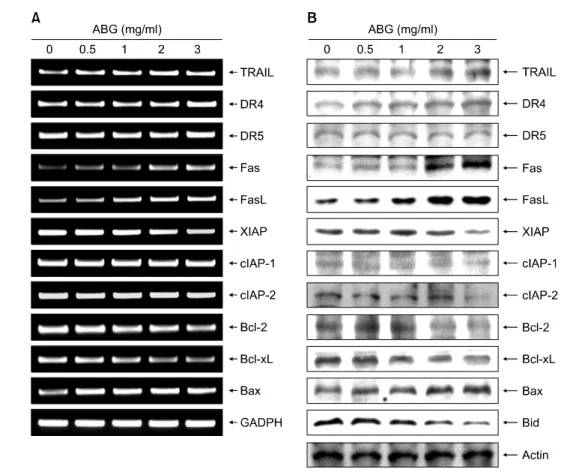
Apoptosis의 조절인자 중 Bcl-2 family에 속하는 몇 가지 중요한 인자들은 apoptosis 유발 조절에 가장 대표적인 유 전자로 알려져 있는데, 그 중 Bcl-2 및 Bcl-xL은 anti- apoptotic 분자로서 apoptosis의 유발을 억제하는 기능을 가지며, Bax는 pro-apoptotic 분자로 Bax 단백질 발현의 증 가는 apoptosis의 유발과 관계가 있다.15) 이 단백질들은 dimer의 형태로 존재하며 상대적 발현의 차이에 의하여 mitochondria로부터의 cytochrome c의 유리에 의한 caspase 및 DNA의 단편화와 연관된 endonuclease 등의 활성을 조 절하는 것으로 알려져 있다.14,15) 따라서 AGS 위암세포에 서 ABG에 의한 apoptosis 유발에 이들 유전자가 관련되어 있는지의 여부를 RT-PCR 및 Western blot 분석으로 전사 및 번역 수준에서 조사하였다.
Fig. 6에 나타낸 바와 같이 Bcl-2 family에 속하는 다양 한 유전자들의 발현 변화에서 대표적인 anti-apoptotic 인 자인 Bcl-2 및 Bcl-xL의 발현은 mRNA 및 단백질 수준에 서 모두 ABG 처리 농도의 증가에 따라 발현의 정도가 감소되었으며, anti-apoptotic Bax의 발현은 상대적으로 매 우 증가되었다. 아울러 Bid의 단편화 현상도 관찰되어 ABG 처리에 의한 apoptosis에서 mitochondria가 관여하고 있음을 알 수 있었다. 따라서 ABG 처리에 의한 AGS 위 암세포의 apoptosis 유발에 있어서 pro-apoptotic 인자의 발
현 증가 및 anti-apoptotic 인자의 발현 감소가 중요한 요 인으로 작용하는 것으로 추정된다.
7. IAP family의 인자들의 발현에 미치는 ABG의 영향
Apoptosis 조절에 관여하는 또 다른 인자 중 IAP family 에 속하는 여러 유전자 산물은 caspase와의 직접적인 결 합을 통하여 그들의 활성을 억제할 수 있을 것으로 밝혀 져 있다.12,13) Fig. 5의 결과에서 ABG 처리에 의한 AGS 위 암세포의 apoptosis 유도에 다양한 종류의 caspase 활성이 유도되었으므로 ABG 처리에 의한 AGS 위암세포의 apo- ptosis 유발에 이들 IAP family 인자들이 관여하는지의 여 부를 조사하였다.
Fig. 6에 나타낸 바와 같이 ABG 처리에 따라서 조사된 IAP family의 3가지 인자들(XIAP, cIAP-1 및 cIAP-2)은 전 사 및 번역 수준에서 모두 ABG 처리 농도 증가에 따라 발현이 모두 감소되었음을 알 수 있었다. 따라서 ABG 처리에 의한 이들 IAP family 인자들의 발현 감소는 caspase의 활성화에 어느 정도 기인하여 apoptosis를 촉진 하는 요인으로 작용하였을 것으로 추정된다.
8. Death receptor 관련 인자들의 발현에 미치는 ABG의 영향
Apoptosis 유발은 mitochondrial pathway라고 알려진 in- trinsic pathway 및 death receptor pathway라고 알려진 ex- trinsic pathway로 크게 구분할 수 있는데, extrinsic pathway
Fig. 6. Effects of ABG on the death-receptor related gene products, Bcl-2 and IAP family members expression in AGS cells.
(A) After 48 h incubation with AGS, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers, and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by 8∼10% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
의 경우는 death receptor들의 상호작용에 의하여 조절되 며 caspase-8은 이 과정의 중요한 initiator caspase로 작용한 다.17,18) Apo-2 ligand로 알려진 tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)은 두 가지 death domain- containing agonistic receptor인 DR4 및 DR5와 결합함으로 서 여러 종류의 암세포에서 apoptosis를 유발하는 것으로 알려져 있다. 하지만 osteoprotegerin, DcR1 (TRAIL-R3) 및 DcR2 (TRAIL-R4)와 같은 세 종류의 decoy receptor와 결합을 하게 되면 TRAIL signaling이 억제된다. 한편 TNF receptor superfamily에 속하는 Fas (CD95/Apo-1)에 FasL (CD95L) 또는 agonistic anti-Fas 항체가 결합하면 Fas clustering, Fas-associa- ted death domain protein (FADD) 및 pro-caspase-8이 death- inducing signaling complex (DISC)를 형성하게 되어 caspase- 8의 활성화에 의한 caspases cascade에 의하여 apoptosis를 유발하게 된다.17,18) 따라서 ABG 처리에 의한 apoptosis 유 발에 death receptor에 속하는 유전자들이 관여하는지를 확인하기 위하여 death receptor와 연관성이 매우 높은 몇
가지 유전자들의 발현 변화를 RT-PCR 및 Western blot analysis 방법으로 조사하였다.
Fig. 6에 나타낸 바와 같이 AGS 위암세포에서 조사된 5가지의 death receptor 연관 유전자들 중, DR-5를 제외한 나머지 유전자들의 발현이 ABG 처리 농도 증가에 따라 모두 전사 및 번역 수준에서 매우 증가되었다. 이는 ABG 의 처리에 의한 apoptosis 유발은 death receptor pathway의 경로를 거쳐 caspase-8을 활성화 시켰음을 의미하는 결과 이다. 아울러 caspase-8의 활성은 Bid의 단편화(tBid)를 유 도하여 tBid가 mitochondria 내로 이동하여 cytochrome c의 세포질 내 유출을 촉진함으로서 caspase-9의 활성이 증가 되었을 것으로 추정된다. 또한 caspase-9의 활성 증가는 caspase-3의 활성을 증가시켜 PARP와 같은 단백질들의 단편화를 유도함으로서 apoptosis가 유발되었을 것으로 생각되어진다.
결 론
본 연구에서는 AGS 인체위암세포 세포의 증식에 미치 는 흑마늘 열수 추출물(ABG)의 영향을 조사하였다. ABG 의 처리 농도의 증가에 따라 농도 의존적으로 AGS 위암 세포의 생존율은 감소되었으며, 이러한 현상이 apoptosis 유발과 연관성이 있었음을 염색질 응축에 따른 apoptotic body의 출현 증가 및 flow cytometry 분석에 따른 sub-G1기 세포의 빈도 증가로 확인을 하였다. ABG 처리에 의한 apoptosis 유발에 관여하는 유전자들의 발현 변화를 RT-PCR 및 Western blot 방법으로 조사한 결과, Bcl-2 family에 속하는 anti-apoptotic 인자인 Bcl-2 및 Bcl-xL의 발 현은 전사 및 번역 수준에서 감소되었고, pro-apoptotic Bax의 발현은 증가되었으며, IAP family 단백질들의 발현 도 감소되었다. 또한 caspase의 활성 증가와 더불어 PARP 와 같은 caspase-3의 기질 단백질들의 분해 등이 ABG에 의한 AGS 위암세포의 apoptosis 유도과정에서 관찰되었 다. 아울러 ABG에 의한 Bid의 cleavage 유도와 MMP 소실 및 death receptor 관련 유전자들의 발현 증가에 따른 caspase-8의 활성화와 caspase-9 및 -3 활성 증대 결과는 ABG에 의한 AGS 위암세포의 apoptosis 유도에는 extrinsic 및 intrinsic pathway가 둘 다 관여하고 있음을 의미하며, 특히 동일 조건에서 생마늘 열수 추출물보다 흑마늘 열 수 추출물의 높은 항암활성 결과는 흑마늘 열수 추출물 이 강력한 암예방 및 항암 효능을 지니고 있음을 암시하 여 준다.
감사의 글
본 연구는 2007년도 지식경제부 지원 경남도립남해대 학 지역연고산업육성사업인 보물섬 클러스터 구축을 통 한 남해마늘산업 핵심역량강화사업 및 농촌진흥청 공동 연구사업(과제번호: 7-19-42)의 지원에 의해 이루어진 것 임.
참 고 문 헌
1) Ariga T, Seki T. Antithrombotic and anticancer effects of garlic-derived sulfur compounds: a review. Biofactors 26, 93- 103, 2006.
2) Shimon LJ, Rabinkov A, Shin I, Miron T, Mirelman D, Wilchek M, Frolow F. Two structures of alliinase from Alliium sativum L.: apo form and ternary complex with aminoacrylate reaction intermediate covalently bound to the
PLP cofactor. J Mol Biol 366, 611-625, 2007.
3) Rybak ME, Calvey EM, Harnly JM. Quantitative deter- mination of allicin in garlic: supercritical fluid extraction and standard addition of alliin. J Agric Food Chem 52, 682-687, 2004.
4) Miron T, Mironchik M, Mirelman D, Wilchek M, Rabinkov A. Inhibition of tumor growth by a novel approach: in situ allicin generation using targeted alliinase delivery. Mol Cancer Ther 2, 1295-1301, 2003.
5) Kodai S, Takemura S, Minamiyama Y, Hai S, Yamamoto S, Kubo S, Yoshida Y, Niki E, Okada S, Hirohashi K, Suehiro S. S-allyl cysteine prevents CCl(4)-induced acute liver injury in rats. Free Radic Res 41, 489-497, 2007.
6) Ahmad MS, Pischetsrieder M, Ahmed N. Aged garlic extract and S-allyl cysteine prevent formation of advanced glycation endproducts. Eur J Pharmacol 561, 32-38, 2007.
7) Chauhan NB. Effect of aged garlic extract on APP processing and tau phosphorylation in Alzheimer's transgenic model Tg2576. J Ethnopharmacol 108, 385-394, 2006.
8) Allison GL, Lowe GM, Rahman K. Aged garlic extract and its constituents inhibit platelet aggregation through multiple mechanisms. J Nutr 136, 782S-788S, 2006.
9) Khanum F, Anilakumar KR, Viswanathan KR. Anticarcinoge- nic properties of garlic: a review. Crit Rev Food Sci Nutr 44, 479-488, 2004.
10) Searle J, Kerr JF, Bishop CJ. Necrosis and apoptosis: distinct modes of cell death with fundamentally different significance.
Pathol Annu 17, 229-259, 1982.
11) Lieberthal W, Koh JS, Levine JS. Necrosis and apoptosis in acute renal failure. Semin Nephrol 18, 505-518, 1998.
12) Zimmermann KC, Bonzon C, Green DR. The machinery of programmed cell death. Pharmacol Ther 92, 57-70, 2001.
13) Schulze-Osthoff K, Ferrari D, Los M, Wesselborg S, Peter ME. Apoptosis signaling by death receptors. Eur J Biochem 254, 439-459, 1998.
14) Jurgensmeier JM, Xie Z, Deveraux Q, Ellerby L, Bredesen D, Reed JC. Bax directly induces release of cytochrome c from isolated mitochondria. Proc Natl Acad Sci USA 95, 4997-5002, 1998.
15) Antonsson B, Martinou JC. The Bcl-2 protein family. Exp Cell Res 256, 50-57, 2000.
16) Park C, Gweon OC, Choi YH, Kim JI. Aged black garlic inhibits cyclooxygenase-2 expression and prostaglandin E2 production by phorbol 12-myristate-13-acetate through In- activation of nuclear factor-κB. Cancer Prev Res 14, 161-170, 2009.
17) Burz C, Berindan-Neagoe I, Balacescu O, Irimie A. Apoptosis in cancer: key molecular signaling pathways and therapy targets. Acta Oncol 48, 811-821, 2009.
18) Henshall DC. Apoptosis signalling pathways in seizure- in- duced neuronal death and epilepsy. Biochem Soc Trans 35, 421-423, 2007.