저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Differential gene expression
in Behçet’s disease and
differential anti-tumor effects of
Tim3 pathway blocking molecules
by
Min-yeong Woo
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
Differential gene expression
in Behçet’s disease and
differential anti-tumor effects of
Tim3 pathway blocking molecules
by
Min-yeong Woo
A Dissertation Submitted to The Graduate School of Ajou
University in Partial Fulfillment of the Requirements
for the Degree of Doctor of Philosophy of Biomedical Sciences
Supervised by
Sun Park, M.D.,Ph.D.
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
ofMin-yeong Woo is approved.
SUPERVISORY COMMITTEE
Ho-Joon Shin In-Hong Choi Seonghyang Sohn Yong-Joon Chwae Sun ParkThe Graduate School, Ajou University
June, 21nd, 2013
i
-ABSTRACT-
PART I
Differential expressionof inflammation-related genes
according to colchicine responsivenessin Behçet’s disease
Behçet disease (BD) is a chronic relapsing, systemic inflammatory disorder which is treated with colchicine. However, some patients are not responsive to it. In this study, I wanted to explore the biomarkers to distinguish colchicine responsiveness. I stimulated peripheral blood mononuclear cells (PBMCs) and CD11b+ cells of BD patients with lipopolysaccharide (LPS) and analyzed the expression of inflammatory cytokines, transcription factors and microRNAs. Compared to colchicine-responsive BD patients (BDR), the expression of IL-6 and TNF-α were significantly increased in colchicine-non-responsive BD patients(BDNR) in both the basal and LPS-stimulated states. Concordantly, the expression of C/EBPδ, C/EBPβ transactiption factor of IL-6, was significantly upregulated in BDNR compared to BDR. Also, the expression of ATF-3 transcript was significantly increased in BDR compared to BDNR. The link of dyregulation in these transcription factors to abnormal cytokine production in BD was also demonstrated. Finally, I found the differential expression of miR-638, miR-4488, miR-3591-3pin BD according to colchicine responsiveness. My results suggest that these molecules might be useful to predict colchicine responsiveness in BD.
ii
Key words: Behçet’s disease,colchicine, LPS, IL-6, TNF-α, C/EBP-β, C/EBP-δ, ATF3, microRNA
iii
-ABSTRACT-
PART II
Differential anti-tumor effects of three forms
of Tim-3 pathway blocking molecules
Tim3 is a molecule containing T cell immunoglobulin variable region (IgV)-like domain and mucin-domain, and known to suppress Th-1 immune response. In a previous study, blocking Tim3 pathway increases anti-tumor immunity leading to tumor growth suppression. In this study, I evaluated the tumor suppressive effect of three different molecules blocking Tim3 pathway: Tim3 IgV like domain alone (Tim3V), fusion protein of Tim3V with mouse immunoglobulin CH2CH3 (Tim3VmIg) and its dimer (Tim3VdIg). Both Tim3VmIg and Tim3VdIg suppress tumor growth but T3V did not. Compared to control Tim3VdIg but not Tim3V reduced frequency of myeloid derived suppressor cells (MDSCs) in splenocytes of mice on 21 day after tumor challenge. These results indicate that Tim3V domain alone is not sufficient for tumor growth suppression but is in the presence of its fusion partner of immunoglobulin CH2CH3.
iv
TABLE OF CONTENTS
ABSTRACT ... i
TABLE OF CONTENTS ...iv
LIST OF TABLES ...ix
LIST OF FIGURES ... x
PART I
I. INTRODUCTION ... 1 A. Behçet’s disease ... 1 1. Clinical features ... 1 2. Diagnostic criteria ... 3 3. Epidemiology ... 4 4. Etiopathogenesis ... 5 5. Therapy ... 10B. Transcription factors and microRNAs regulating proinflammatory cytokines expression ... 11
1. Transcription factors ... 11
2. MicroRNA ... 12
v
II. MATERIALS AND METHODS ... 15
A. MATERIALS ... 15 1. Subjects ... 15 2. Culture media ... 18 3. Reagents ... 18 4. Antibodies ... 18 5. Oligonucleotide ... 19 B. METHODS ... 19
1. Human peripheral blood mononuclear cells (PBMCs) isolation ... 19
2. Cell cultures ... 20
3. CD11b+ cell isolation ... 20
4. Western blotting ... 20
5. Lactate dehydrogenase (LDH) release assay ... 20
6. Enzyme-linked immunosorbent assay (ELISA) ... 21
7. siRNA transfection ... 21
8. RNA isolation and cDNA synthesis ... 21
9. Real-time reverse transcription–polymerase chain reaction (Real-time RT PCR) ... 21
10.Construction of luciferase reporter vectors ... 21
11. Luciferase reporter assay ... 22
vi
III. RESULTS ... 26
A. Colchicine inhibits caspase-1 activation ... 26
B. Effect of colchicine on the release of interleukin-1β (IL-1β) is dependent on theinflammatory stimulus and responsiveness to colchicine ... 28
C. Colchicine does not inhibit lactate dehydrogenase (LDH) release in PBMCs ... 31
D.Colchicine differentially regulates IL-1β transcription in BD PBMCs according to their colchicines responsiveness ... 33
E.Transcription factors regulating proinflammatory cytokine production are differentially expressed according to colchicines responsiveness... 38
F.The principal cells abnormally expressing cytokines in BDNR are CD11b+ cells ... 42
G. The knocked-down effect of various transcription factors on expression of TNF-α and IL-6 in THP-1 cells ... 45
H. The differential effect of siRNA targeting C/EBPβ, C/EBPδ or ATF3 on the cytokine production in CD11b+ cells ... 48
I.The differential expression of micro RNAs in BD according to colchicine responsiveness ... 50
IV. DISCUSSION ... 56
V. CONCLUSION ... 63
REFERENCES ... 64
vii
PART II
I. INTRODUCTION ... 82
A.The TIM gene family ... 82
B. Tim3 ... 83
1. Expression ... 83
2. Structure ... 83
3. Ligand ... 84
4. Function in T cell responses ... 85
5. Roles in tumor immunity ... 86
C. Roles of myeloid-derived suppressor cells (MDSCs) ... 87
D. Tumor-associated macrophages (TAMs) ... 89
E. Purpose ... 90
II. MATERIALS AND METHODS ... 91
A. MATERIALS ... 91
1. Mouse ... 91
2. Culture media ... 91
3. Antibodies... 91
B. METHODS ... 92
1.Construction of expression vectors ... 92
2.Prodcution of recombinant lentiviruses ... 92
viii
4. Estabilishment of stable cells expressing Tim3 pathway blocking molecules ... 93
5. Measurement of cell growth rate... 93
6.Evaluation of tumor growth ... 94
7. Tumor vaccination ... 94
8.Flow cytometric analysis of leukocyte subpopulation ... 94
III. RESULTS ... 96
A. The characterization of Tim3 pathway blocking molecules ... 96
B. The tumor growth of 3LL cells stably expressing Tim3 pathway blocking molecules ... 99
C. The therapeutic effects of 3LL cells stably expressing Tim3 pathway blocking molecules ... 101
D.The effect of Tim3 pathway blocking molecules on the frequency of MDSCs ... 103
IV. DISCUSSION ... 108
V. CONCLUSION ... 111
REFERENCES ... 112
ix
LIST OF TABLES
PART I
Table1.Smmary of miRNA involvement in the immune response ... 14
Table 2.The clinical features of BD patients ... 16
Table3.Drugs taken by the patients at the time point of bleeding ... 17
Table4.Primers for real-time RT PCR ... 24
Table5.Sequence of siRNAs... 25
Table6.Primers for CRTC1 3’UTR amplication... 25
Table 7.Differential expression of genes related to inflammation according to colchicine responseiveness in Behçet’s disease ... 55
PART II
Table.1. Primers for cloning of Tim3 pathway blocking molecules ... 95x
LIST OF FIGURES
PART I
Fig. 1.Diagram of luciferase reporter vectors containing CRTC1 3’UTR proximal and distal
region, respectively ... 23
Fig. 2.Cleavage of caspase-1 is inhibited by colchicine ... 27
Fig. 3.Effect of colchicine on interleukin(IL)-1β release ... 30
Fig. 4.Lactate dehydrogenase (LDH) release is not inhibited by colchicine ... 32
Fig. 5.Effects of colchicine on the transcript levels of interleukin (IL)-1β, tumour necrosis factor (TNF)-a and IL-6 differ in Behc¸et disease peripheral blood mononuclear cells (PBMCs) according to their colchicine responsiveness ... 35
Fig. 6.The protein levels of TNF-αand IL-6 differ in BDPBMCs according to their colchicine responsiveness. ... 37
Fig.7.Transcript levels of various transcription factors regulatingthe proinflammatory cytokine expression ... 40
Fig. 8.The protein levels of various transcription factors regulating the proinflammatory cytokine expression ... 41
Fig. 9.The expression of proinflammatory cytokines and their regutory transcriptionfactors in CD11b+ and CD11b- PBMCs ... 43
xi
Fig. 10.The knock-down effect of various transcription factors on the expression of TNF-α and IL-6 in THP-1 cells ... 46 Fig. 11.Knockdown of transcription factors differentially affects production of TNF-α and IL-6 in CD11b+ cells according to colchicines responsiveness ... 49 Fig. 12.The differential expression of micro RNAs in BD PBMCs according to colchicine responsiveness ... 52 Fig. 13.The miR-4488 mimic oligonucleotide targets distal region of CRTC1 3’UTR... 53 Fig. 14.The expression of miR-4488 and its putative target CRTC1 mRNA ... 54
PART II
Fig. 1.The characterization of Tim3 pathway blocking molecules ... 98 Fig. 2.The tumor growth in mice challenged with stable cell lines expressing Tim3 pathway
blocking molecules. ... 100 Fig. 3.The therapeutic vaccine effects of T3V and T3VdIg in a tumor model ... 102 Fig. 4.The effects of Tim3 pathway blocking molecules on MDSCs frequency ... 105
Differential gene expression
in Behçet’s disease and
differential anti-tumor effects of
Tim3 pathway blocking molecules
PART I
Differential expression of
inflammation-related genes according to colchicines
1
I.
INTRODUCTION
A. Behçet’s disease (BD)
Behçet’s disease (BD) is a multisystem, chronic relapsing, systemic inflammatory disorder characterized by four major symptoms consisting of oral aphthous ulcers, genital ulcers,uveitis, and skin lesions, and sometimes by inflammation in tissues and organs throughout the body including the vascular system, central nervous system, gastrointestinal tract, lungs, kidneys and joints (Sakane et al., 1999). It was first reported in 1937 by Hulusi Behçet who is the Turkish dermatologist as a symptom complex of recurrent oral ulcers, genital ulcers, and uveitis (Behçet, 1937). A complicated genetic background leading to a pro-inflammatory, innate-immune-system-derived activation, adaptive immune response against environmental and auto-antigens are accepted to be the immunopathogenic aspects of BD (Direskeneli, 2001).
1. Clinical features
Symptoms of Behçet’s disease are recurrent, which may last a few days to several weeks and sometimes may cause peripheral tissue damage or even death.Mucocutaneous findings are the most frequently observed hallmarks of BD. These symptoms are oral and genital ulcers, papulopustular lesion, erythema nodosum-like vasculitic nodular lesions, acneiform lesions and superficial thrombophlebitis (Gül, 2005). Notably ulcerations akin to recurrent aphthous stomatitis appear in all BD patients (Al-Otaibi et al., 2005). The
2
oralulceration is the earliest symptoms of up to 86.5 % of adults and children diagnosed with BD (Ekmekçi et al., 2003). Although the oral ulcers of BD may be more common on the soft palate and pharynx than in recurrent aphthous stomatitis, they have the equal periodicity as those of recurrent aphthous stomatitis and can influence any oral mucosal surface (Main et al., 1992). The genital ulcers of BD initially appeared as papules or pustules that later ulcerate. The ulcers seem to be indurated and can develop into secondary infection. In males, the ulcers are mainly localized on the inguinal area, pubis, scrotum, penis and perineum. In females, major and minor labia, the vulvae, cervix, and vagina are the most frequently affected sites. The ulcers are usually cured in a few weeks, with scar (Nazzaro, 1964; Al-Otaibi et al., 2005). Genital ulcers are less common than oral ulcers and less frequent in children than adults (Krause et al., 1999). The most common cutaneous symptoms are papulo-pustular lesions, erythema nodosum, and erythema nodosumlike disease. Papulo-pustular lesions are cutaneous folliculitis or pimple-like lesions on an erythematous base which are initially manifested as papules and developinto pustules within 24 to 48 h. These lesions generally appear concurrently, most frequently on the skin of the face, back and chest. The papulo-pustular lesions are considered histopathologically as a lymphocytic vasculitis (Lakhanpal et al., 1985; Al-Otaibi et al., 2005). Similarly, the histopathology of the erythema nodosum shows focal vasculitis of small-vessels and perivascular lymphocytic infiltrate, notably involving the venules, with panniculitis (Chun et al., 1989). Other cutaneous lesions, such as necrotic folliculitis and aphthosis, may be seen in BD, but to a lesser frequency (Al-Otaibi et al., 2005).
3
uveitis, panuveitis, retinal vasculitis, bilateral swelling of the optic nerve head and bilateral lamellar macular hole. Ocular menifestations change from a gritty sensation and blurred sign to acute pain and blindness (al Aboosi et al., 1996). BD patients with predominant anterior uveitis have a comparatively good visual prognosis. Usually, a poorer prognosis is more common in male patients than females (Yazici et al., 1984). The presence of HLA-B51 enhance susceptibility to ocular disease (Nishiyama et al., 2001), and Factor V (FV Leiden) mutation seems to be an additional factor for the progress of ocular disease, particularly retinal vaso-occlusion (Verity et al., 1999).
The neurological complications of BD mainly involve the central nervous system (CNS), despite of rare involvement of muscle and peripheral nerves. CNS symptoms include pseudotumor cerebri, brain-stem involvement, neuropsychiatric symptoms, and meningo-encephalitis (Martini,1995).
The effects of the disease may be cumulative, especially with neurologic, vascular, and ocular involvement. Particularly, ocular manifestations are the most common cause of important morbidity in BD, since it may lead to blindness in about five years if not treated (Shimizu et al., 1979). The mortality rate is low, but death can occur from neurologic involvement, vascular disease, bowel perforation, cardiopulmonary disease, or as a complication of immunosuppressive therapy (Boe et al., 1958).
2. Diagnostic criteria
There are no pathognomonic laboratory findings or histological features for BD diagnoses. Therefore diagnoses are made according to clinical criteria proposed by the
4
International Study Group for Behçet’s Disease (Lancet, 1990) or the Behçet’s Disease Research Committee of Japan (Kurokawa et al, 2004).
According to the International Study Group guidelines, the assented diagnostic criteria need the presence of recurrent oral ulceration (three times in one year) plus two of the following in the absence of other systemic disease: recurrent genital ulceration, eye lesions (uveitis or retinal vasculitis), skin lesions (erythema nodosum, pseudofolliculitis, papulo-pustular lesions, or acneiform nodules), or a positive pathergy test (Lancet, 1990). More recently, Kurokawa MS et al described that BD symptoms are defined in more expanded range including minor symptoms such as involvement of vessels, gastrointestinal tract, and CNS. Therefore the salientclinical and histopathological features of BD are considered as clinical criteria (Kurokawa et al, 2004).
3. Epidemiology
BD has been described all over the worlds, but its prevalence is certainly higher in eastern Mediterranean areas and Asia, particularly Japan and China (Shimizu et al., 1979;Ohno, 1982). The number of BD patients has been increasing ever since the first report on BD in Korea was published in 1961 (Joo, 1961). According to 2008 report, there are 29816 patients in Korea.
The mean age at the time of diagnosis is most commonly in the 30s, even though the age at the time of final diagnosis mostly is in the 40s (Bang et al., 1997). Notably, about 2~3% of all BD patients have childhood-onset of BD (generally described as definitive BD before the age of 16 years) (Saylan et al., 1999). Childhood-onset of BD patients who have the age
5
of 2 years have been reported (Kari et al., 2001), but generally children develop symptoms between 7 and 13 years of age (Koné-Paut et al., 1998, 2002; Gület al., 2000). As a family history of BD is more common among patients before the age of 10 years, family examination for BD is regarded critical for early and precise diagnosis of BD, to provide effective method for control its complications (Khairallah et al., 2012).
Inconsistent results have been reported regarding gender preponderance of BD (Al-Otaibi et al., 2005). The female preponderance was commonly reported in studies with Korean patients, Singapore subjects, Israeli Jews, and subjects in Camas village of Tureky (Bang et al., 1997; Tan et al., 1999;Krause et al., 1999, 2001;Tüzün et al., 1996). While male preponderance has been noted in the West Indies (Lannuzel et al., 2002), Spain (Gonzalez-Gay et al., 2000;Baixauli et al., 2001), Israeli Arabs (Krause et al., 2001), London (Muhaya et al., 2000), Japan (Muhaya et al., 2000), Jordan (al Aboosi et al., 1996), Iran and Turkey (Gurler et al., 1997; Kaya et al., 2002), India (Pande et al., 1995), Lebanon (Ghayad and Tohme, 1995), Saudi Arabia (al Dalaan et al., 1994), and Kuwait (Mousa et al., 1986). It may suggest the relevance of environmental factors for the pathogenesis of BD (Bang et al., 1997).
4. Etiopathogenesis
The cause of BD remains unknown, but an autoimmune reactions triggered by an infectious or environment agent or genetic factors are strongly associated with pathogenesis (Direskenli 2001, M. Khairallah et al., 2012). Immunopathogenic aspects of BD include neutrophil activation, cellular and humoral immunity to antigenic stimuli such as herpes
6
simplex virus(HSV), streptococci, superantigens, heat shock proteins (65 kDa, αβ-crystallin), and as autoantigens such as α-enolase (Direskenli 2001). Archetypal BD lesions such as pustular folliculitis, pathergy reactions and hypopyon show neutrophil infiltrations and neutrophil activation. In a recent report, fMLP (a strong chemoattractant)-induced superoxide production was decreased in BD(Direskenli, 2001). When prestimulated BD neutrophils were successively stimulated, they produced a lower increase in superoxide production than in healthy controls, signifying a state of in vivo neutrophil preactivations (Ekşioglu-Demiralp et al., 2001; Direskenli, 2001). In a previous study, in vivo “primed” state of neutrophils has been suggested that neutrophil oxidase is activated with dual signalling system (Hallett, 1995). According to this report, tyrosine phosphorylation of various intracellular proteins seems to be increased by agents such as fMLP or the cytokines including tumor necrosis factor-α (TNF-α) and granulocyte monocyte colony stimulating factor. BD neutrophils seem to be primed in vivo with proinflammatory cytokines without full activation (Hallett,1995;Direskenli, 2001).
T cell activation, both in tissue specimens and peripheral blood, is demonstrated in BD (Direskenli, 2001). As in most other autoimmune disease and vasculitides, the Th1-type cytokine expression is noticeable in BD. CD4+ and CD8+ T cells which produce the Th1-type proinflammatory cytokines are increased in peripheral blood and related with disease activity (Sugi-Ikai et al., 1998; Direskenli, 2001). Moreover, expression of costimulatory molecules such as PD-L1, CTLA-4 and CD86 which regulate T cell response is more decreseaed in active BD patients than healthy controls. Therefore these results suggestthat PD-L1, CTLA-4 and CD86 are involed in BD pathogenesis (Sim et al., 2011). Although BD
7
does not have aspect of a classical autoimmune disease, it is reported that the spontaneous production of Ig was increased (Suzukiet al.,1886). Autoantibodies such as antiendothelial cell (AECA), antilymphocyte, and anticardiolipin antibodies, especially of IgA isotype, are also present in BD patients (Direskeneli et al., 1995; Matsui et al., 1999; Tokay et al., 2001). B cells express high levels of activation markers such as CD13, CD33, CD80, and the memory marker CD45RO despite of normal levels of total B cells number (Ekşioglu-Demiralp et al., 1999). Given that the role of B cells in the immune system is not limited to antibody production, but also includes antigen presentation, the activated and memory type B cell profile might modulate T cell activation in BD (Direskenli, 2001).
Abnormal production of proinflammatory cytokines such as IL1, IL-6 and TNF-α in BD has been reported. IL-1 family cytokines such as IL-1α and IL-β have been increased in sera of BD patients(Zhou, 2012). IL-6 levels are associated with disease activity of BD. Furthermore, IL-6 plays a peculiar role in BD with central neuronal system. Circulating TNF-α levels are increased in the peripheral blood of BD patients (Sfikakis, 2007). Also increased numbers of monocyte and the γδ+ T cells highly producing TNF-α are reported in BD patients. Underlying mechanisms for these finding remain elusive, however, Single Nucleotide Polymorphisms (SNPs) of IL1 leading to the increased expression of IL1 and variant number tandem repeats (VNTR) in 3’untranslated region of IL-6 were observed in BD.
Besides these proinflammatory cytokines such as Th1 cytokines also play important roles in BD. Notably, IL-2 andIFN-γ are increased in BD patients with uveitis. Moreover, SNPs of Th1 type cytokines such as IL-12 and IFN-γ are closely linked to BD pathogenesis.
8
Genome Wide Association Studies (GWAS) from Turkey and Japan independently indicated that IL-12R-IL23RB2 region also associated with BD (Zhou, 2012). Moreover, proinflammatory cytokine IL-17 which is expressed in Th17 cells is considered to be associated with inflammatory process of BD. In line with previous reports, the high IL-17 levels in BD were reported. Although the functional relevance has not been studied, IL17F A126G is more frequently found in BD patients(Zhou,2012).
The relevance of microbial factors with BD pathogenesis has been proposed by detection of HSV-1 DNA in peripheral blood lymphocyte and monocytes from BD patients (Eglin et al., 1982; Bonass et al., 1986), and circulating antibodies to HSV-1 with increased frequency compared to healthy and disease controls (Al-Otaibi et al., 2005). The BD-like symptoms develop in HSV-infected mouse. Similar to BD patients, these mice show differential expression of CCR7 in peripheral blood mononuclear cells (Choi et al., 2013). HSV has been currently known only virus possibly associated with BD (Yazici et al., 1998; Sakane T et al., 1999).
The role of oral microbial flora has long been suggested in BD pathogenesis considering that oral aphthae are the first symptom over 70% of BD patients (Direskenli, 2001). Particularly, Streptococcus sanguinis seems to have some etiological play in BD. The patients with BD patients may have higher numbersof S. sanguinis in the mouth than healthy controls (Isogai et al., 1990). Moreover enhanced antibody and T cell responses to S.
sanguinis Ags were detected in BD patients compared to normal controls (Lehner et al.,
1991; Hirohata et al., 1992; Yokota et al., 1992). In addition, Bes-1 Ag of S. sanguinis is homologous to human retinal protein, suggesting the implication of S. sanguinis in BD
9
uveitis. Oral streptococcal heat shock protein (HSP) has been suggested as a pathogenic Ag cross reactive with oral mucosal homogenated in BD (Lehner et al., 1991). Both antistreptococcal and antiretinal HSP60 antibodies are increased in the serum of patients with BD and uveitis (Tanaka et al., 1999).
BD has a strong genetic association with familial aggregation. The association of the HLA-B locus in BD has been recently reported (Gület al., 2001). HLA-B51, one of the split antigens of HLA-B5, has been increased in frequency in BD patients from Turkey (Kaya et al., 2002), Japan (Mizuki et al., 1997), Greece (Mizuki et al., 2002), Israeli Jews and Israeli Arab (Krause et al., 2002). Particularly, HLA-B*501 is the critical subtype of HLA-B51 associated with BD. Although the function of HLA class I antigen is the presentation of endogeneous antigens which synthesized within the cell to CD8+ cytotoxic T cells, whether HLA-B51 restricted CD8+ T cells participate in the pathogenesis of BD is currently unknown. Notably, neutrophils from HLA-B51 transgenic mice but not those from control mice produce significant amounts of superoxide when stimulated with fMLP(Direskenli, 2001).The exact mechanism that the HLA-B51 molecules affect neutrophil function should be dissected.
Recent genome-wide association studies (GWAS) have revealed genetic risk factors in BD. Elanine F Remmers and colleagues conducted a GWAS with 311,459 SNPs in BD patients from Turkey. They identified common variants in IL10 and IL23R-IL12RB2 locusare associated with BD(Remmers et al., 2010). In another report the association of chromosomes 1p31.3 (IL23R-IL12RB2, re12119179) and 1q32.1 (IL10, rs1554286) with BD was suggested(Mizuki et al., 2010). Recently, Yohei Kirino and colleagues performed a
10
GWAS of 779,467 SNPs with BD patients from Turkey. They identified the new susceptibility loci for BD at CCR1-CCR3 (encoding C-C chemokine receptors1-3), STAT4 (encoding signal transducer and activator of transcript 4) and KLRC1-KLRC4 (encoding killer cell lectin-like receptor subfamily C, member 1-4) and ERAP1 (encoding ERAP1). Moreover they found the interaction of HLA-B*51 and ERAP1 (Kirino et al., 2013).
5. Therapy
The treatment of BD patients remains largely symptomatic and empirical, but is generally specific to the clinical trait of each patient. The treatment of BD patients is guided by organ involvement. As topical remedy, steroids are usually effective for mucocutaneous involvement. Systemic administration of anti-inflammatory drug as a primary drug such as colchine is used. Combinations of corticosteroids and non-selective immuno-suppressive drugs such as azathioprine, methotrexate, cyclosporine A, tacrolimus, chlorambucil, or cyclophosphamide are used when critical organs are involved (Al-Otaibi et al., 2005). Some patients insufficiently respond to remedy and need additional treatments such as IFN-γ and TNF-α blockers(Sfikakis PP, 2002).
Colchicine is an alkaloid derived from the plant of the Lily family Colchicum
autumnale (Niel et al., 2006). Colchicine has been used to treat gout, familial Mediterranean
fever (FMF), BD, Sweet’s syndrome, scleroderma, amyloidosis and liver cirrhosis for several decades (Ben-Chetrit et al., 2006). The primary mechanism of colchicine is suggested to inhibit microtubule polymerization(Niel et al., 2006). Microtubules are an element of the cytokelaton which is important for regulation of many functions including
11
cell migration, division, and polarization. Each colchicine molecule binds to a tubulin molecule and then prevents its polymerization. Consequently, microtubule elongation is interrupted, the miotic spindle is disturbed, and cell division cannot proceed. As a result, signaling pathways, intracellular vesicle motility and secretion of cytokines or chemokines are inhibited. Colchicine affects adhesion, mobility, phagocytosis and cytokine production of leukocytes leading to suppression of inflammation(Ben-Chetrit et al., 2006). These mechanisms account for most of the anti-inflammatory effects of colchicine, which give rise to many therapeutic uses (Niel et al., 2006).
B. Transcription factors and microRNAs regulating proinflammatory cytokines expression
1. Transcription factors
Transcriptional regulation of proinflammatory cytokine production has been studied in cells stimulated with LPS. Induction of the LPS-dependent TNF-α and IL-6 transcription is modulated by various transcription factors such asnuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB),activating transcription factor 3 (ATF3), CCAAT/enhancer-binding proteinβ (C/EBPβ), and C/EBPδ (Medzhitov et al., 2009). NF-κB functions as the transcription activator. NF-κB is constitutively expressed and retained in cytoplasm in basal state, but stimulation translocated into the nucleus after stimulation. ATF3 is inducible repressor of IL-6. ATF3 is a member of the cAMP response element-binding
12
protein (CREB) family and has been shown to form a regulatory circuit withNF-κB and C/EBPδ for IL-6 expression. In the absence of ATF3, over-expression of IL-6 and the increased susceptibility to endotoxin shock have been demonstrated (Hoetzenecker et al., 2011).ATF3 induced by NF-kB activation prevents cells from endotoxic shock (Maitra et al. 2011). C/EBPδ is rapidly induced by LPS stimulation and amplifies IL-6 expression (Medzhitov et al., 2009). In C/EBPδ-/-mice, expression of TNF-α, IL-6 and IL-1β is attenuated by LPS stimulation therefore C/EBPδ is important in induction of LPS-induced gene (Valente et al., 2013).C/EBP also plays an important role in inflammatory response. C/EBPβ binds to TNF-α gene promoter and activates it (Pope et al., 1994). Further, C/EBPβ enhances NF-kB activation by down-regulation ofIκB (Cappello et al., 2009).
2. MicroRNA
MicroRNA (miRNA) is involved in the posttranscriptional regulation of gene expression. It was first identified in Caenorhabditis elegans in 1998 (Fire et al., 1998).miRNAs are small, evolutionary conserved noncoding RNAs. These are derived from larger primary transcript encoded in the genome. Precursor form of miRNA most commonly transcribed by RNA polymeraseII is processed by Drosha and DGCR8 in the nucleus resulting in pre-miRNA which is exported to the cytoplasm and converted to duplex miRNA through the action of Dicer. One of the miRNA strands is loaded on the RNA-induced silencing complex (RISC) which guides miRNA to the 3’UTR of target messenger RNAs leading to repression of target protein expression (O'Connellet al.,2012).
13
(Table 1).MiR-155 suppresses the expression of negative regulator such as SOCS1 and SHIP1, thus leading to increased activation of AKT and IFN response gene (Androulidaki et al., 2009; O'Connell et al., 2012).On the other hands, miR-146a functions as a negative regulator during inflammatory response (O'Connell et al., 2012). Mir-146a suppresses the expression of TRAF6 and IRAK1 which involved in transduction of TLR signaling and NF-κB activation (O'Connell et al.,2012).Other miRNAs suppressing production of TNF-α or IL-6 are miR-92a, miR-203 and miR-187.Mir-92atargets mitogen-activated protein kinase kinase 4 (MKK4) and inhibits production of TNF-α and IL-6 by LPS stimulation (Lai. et al., 2013).MiR-203 targets MyD88 and results in repression of NF-κBactivation and subsequent production of TNF-αand IL-6 in macrophage RAW264.7 cells with LPS stimulation (Wei et al., 2013).MiR-187 directly targets TNF-α and indirectly reduceses IL-6 and IL-12p40 expression in human monocyte with LPS stimulation (Rossato et al., 2012).
C. Purpose
Colchicine is one of the first line drugs for treatment of Behcet’s disease (BD) and known to inhibit caspase-1 activation and inflammatory cytokine production. However, the therapeutic and preventive effect is not observed in some BD patients. In this study, I wanted to explore whether the effects of colchicine on proinflammatory cytokine expression and cell death in peripheral blood mononuclear cells (PBMC) are associated with colchicine responsiveness of BD patients. Also, I wanted to investigate whether the expression of transcription factors and microRNA which regulate inflammatory cytokine transcription is correlated with colchicine responsiveness of BD patients.
14
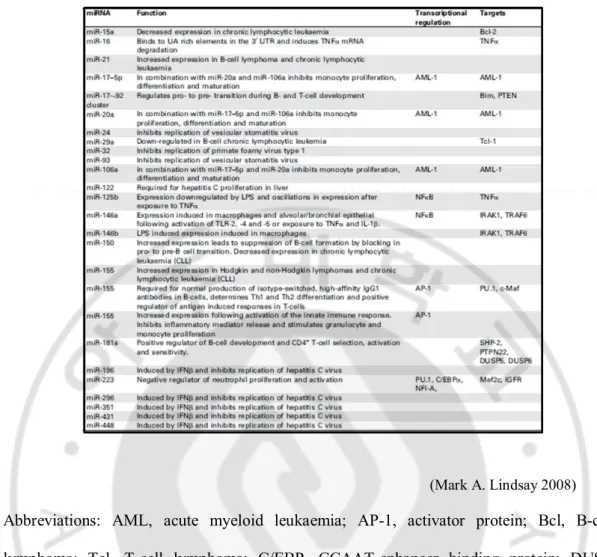
Table 1.Summary of miRNA involvement in the immune response
(Mark A. Lindsay 2008)
Abbreviations: AML, acute myeloid leukaemia; AP-1, activator protein; Bcl, B-cell lymphoma; Tcl, T-cell lymphoma; C/EBP, CCAAT-enhancer binding protein; DUSP, dualspecificity phosphatase; IGFR, insulin-like growth factor receptor; IRAK, IL-1 receptor activated kinase; JNK, c-jun N-terminal kinase; Maf, musculoaponeurotic fibrosarcoma:Mef, myeloid ELF-1 like factor; miRNA, microRNA; NF, nuclear factor; PTEN, phosphatase and tensin homolog; PTP, protein tyrosine phosphatase; SHP, Src homology 2domain– containing protein-tyrosine phosphatase; TLR, Toll-like receptor; TRAF, TNF receptor-associated factor.
15
II.
MATERIALS AND METHODS
A. MATERIALS
1. Subjects
Patients with BD who presented for the first time or who werebeing monitored at the Department of Dermatology, AjouUniversity Hospital from February to December 2011 wereincluded. The diagnosis was determined using the BD criteriaset by the International Study Group and the Japanesecriteria. The patients were divided into two groups, colchicineresponders (BDR) and colchicine nonresponders (BDNR),based on their response to colchicine treatment. Response tocolchicine (BDR group) was defined as an improvement in preexistingsymptoms, especially erythema nodosum and oral andgenital ulcers, without recurrence during a 3-month treatmentwith colchicine (1.2 mg daily). Nonresponse to colchicine(BDNR group) was defined by recurrence of symptoms or thedevelopment of a new symptom without amelioration of preexistingsymptoms after 3 months of colchicine treatment. Themean age was 48.1 ± 7.6 years for the BDR group and43.9 ± 9.9 years for the BDNR group (male to female ratio,5 : 11 for BDR and 6 : 11 for BDNR). The clinical features ofthe patients and medications taken are summarized inTables 2and 3. Blood from 12 healthyvolunteers with no inflammatory disorders was used as a control.Informed consent was obtained from all subjects prior toenrolment in the study. This study was approved by the InstitutionalReview Board(AJIRB-GEN-GEN-10-119).
16
Table 2.The clinical features of BD patients Clinical characteristics BDR (n = 16) % BDNR (n = 17) % Oral ulcer 15 93 17 100 Genital ulcer 11 68 12 70.6 Erythema nodosum 11 68 11 64.7 Ocular symptoms 8 50 8 47.1 Arthralgia 7 43 5 29.4 Gastrointestinal involvement 3 18 2 11.8 Neurological involvement 1 6 1 5.9 Thrombophlebitis 0 0 0 0 The skin pathergy test 3 18 3 17.6 HLA-B51 9a 56 4b 23.5
*The numbers of of subjects with non-HLA-B51 are 5 in BDR and 9 in BDNR,respectively. HLA type of 6 subjects is undetermined.
17
Table 3.Drugs taken by the patients at the time point of bleeding
BDR (n = 16) % BDNR (n = 17) % Colchicine 15 93 15 88.2 Prednisolone 3 18 5 29.4 methyl-prednisolone 3 18 4 23.5 Azathioprine 1 6 2 11.8 Salazopyrine 1 6 3 17.6 Pentoxyphylline 8 50 10 58.8 Cilostazol 0 0 1 5.9 Minocycline 3 18 2 11.8 Aspirin 1 6 1 5.9 Cyclosporine 1 6 0 0 Hydantoin 0 0 1 5.9 Acetazolamide 0 0 1 5.9 Pentasa 2 12 0 0 MTX 0 0 1 5.9 Dexamethasone 0 0 1 5.9 hydroxy chloroquine 0 0 1 5.9
18
2. Culture media
Roswell Park Memorial Institute (RPMI) 1640 (GIBCO, 31800-022, Invitrogen, Carlsbad, CA, U.S.A) and Fetal bovine serum(FBS, GIBCO, 16000-044), and 100 U/ml penicillin and 100 μg/ml streptomycin (GIBCO BRL, 15140-122) were used for cell cultures.
3. Reagents
Ficoll–Paque PLUS was purchased from GE Healthcare(17-1440-02, U.S.A.). Adenosine 5′-triphosphate disodium salt (ATP; 51963-61-2), colchicine (C3915) and lipopolysaccharide (LPS) were purchased from Sigma-Aldrich(St Louis, MO, U.S.A.). pCDNA3 (Invitrogen, Gaithersburg,MD, U.S.A.) was used as double-stranded (ds)DNA. Monosodium urate (MSU) crystals (1198-77-2) were purchased from InvivoGen (San Diego, CA, U.S.A.). Trizol was purchased from Invitrogen (Gaithersburg,MD, U.S.A.) and enhanced chemiluminiscence (LR01-01) was purchased from Animal Genetics (U.S.A.). Lipofectamin 2000 reagent (11668-019) was purchased fromInvitrogen
4. Antibodies
Anti-caspase-1 (#3866S) was purchased from Anderson ling (DanversMA, U.S.A). Anti-CD11b microbeads (130-049-601) were purchased from Miltenyi Biotec (Germany). Anti-C/EBP-β (ab32358), anti-C/EBP-δ (ab65081) were purchased from Abcam(Cambridge UK). Anti- ATF3 (SAB2103491-50UG) was purchased from Sigma-Aldrich (St Louis, MO, U.S.A.). Anti-mouse-IgG-HRP was purchased from cell signaling. Anti-rabbit-IgG-HRP (G21234) was purchased from Invitrogen. Human Fc receptor binding inhibitor (16-9161-73)
19
was purchased from eBioscience (San Diego, CA,U.S.A.) Anti-α-tubulin antibody was purchased from Sigma-Aldrich(St Louis, MO, U.S.A.) and anti-β-actin antibody was purchased from Bethyl(A300-491A, Montgomery, TX, U.S.A.).
5. Oligonucleotide
All siRNAs were purchased from INTEGRATED DNA TECHNOLOGIES (IDT;Coralville, Iowa,U.S.A.). FITC-labeled negative control oligonucleotide was purchased from BIONEER (SN1021,Daejeon,Korea). MiR-4488 mimic (4464066), anti-miR-4488(antagomir;4464) or corresponding control oligonucleotide (AM17121, AM17012) were purchased from Ambion (Carlsbad, California, U.S.A.).
B. METHODS
1. Human peripheral blood mononuclear cells (PBMCs) isolation
PBMCs were isolated by using Ficoll –Paque PLUS. Ten ml of blood from healthy donors or Behcet's disease patients was diluted with 25 ml of PBS (containing 2mM EDTA, pH 7.2) and then was carefully layered on Ficoll –Paque PLUS. After centrifugation at 400 xg for 40 minutes at 20 °C with no break, PBMCs layer was isolated and washed 2 times with PBS buffer.
20
2. Cell cultures
Isolated PBMCs, THP-1 (human monicytic cell line) and Jurkat T cells (human T lymphocytic cell line) were cultured in RPMI1640 supplemented with10 %FBS and100 U/ml penicillin and 100 μg/ml streptomycin. Cells were incubated in the 5% CO2 incubator at 37 °C.
3. CD11b+ cell isolation
CD 11b+ cells isolated from total PBMCs using a CD11b+ microbeads according to the manufacturer’s instructions. Purity of isolated CD11b+ cells was measured by flowcytometry and was above 80% (data not shown).
4. Western blotting
PBMCs (3x106) were lysed in 20μl of lysis buffer (50 mM HEPES, 250 mM NaCal, 1 mM EDTA, 1 mM DTT, 1% Triton X-100) containing protease inhibitors (Roche, Basel,Swiss Confederation). The lysate was subjected to Western blotting using Abs to C/EBPβ, C/EBPδ, ATF3 and the active form of caspase-1.
5. Lactate dehydrogenase (LDH) release assay
LDH release was measured using an LDH Cytotoxicity Detection Kit (TaKaRa Bio) according to the manufacturer’s instructions. The percentage of LDH release was calculated as: 100 x (LDH release of sample – spontaneous release of LDH) ⁄(maximum release of LDH – spontaneous release of LDH).
21
6. Enzyme-linked immunosorbent assay (ELISA)
The release of IL-1β, IL-6 or TNF-α was analyzed using ELISA Kits (IL-1β: Invitrogen, KHC0012; IL-6:eBioscience, BMS/213; TNF-α: eBioscience, BMS2034) according to the manufacturers’ instructions.
7. siRNA Transfection
5x 105 cells were transfected with 20 nM of siRNAs using Lipofectamine 2000.
8. RNA isolation and cDNA synthesis
Total RNA was isolated from the cells using Trizol Reagent.cDNAwas synthesizedusing the SuperScript™ II Reverse Transcriptase(Invitrogen, 18064-014).
9. Real-time reverse transcription–polymerase chain reaction (Real-time RT PCR) Real-time RT PCR was performed using SYBR Premix Ex Taq (TaKaRa Bio) and primers (Table4). The transcript levels were normalized to that of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) or actin. PCR was performed in an ABI PRISM 7000 Sequence Detection System (Applied Biosystems, Foster City, CA, U.S.A.). For real-time quantification of microRNAs, standard TaqMan Small RNA assays kit was used.
10. Construction of luciferase reporter vectors
The proximal and distal regions of 3’UTR of CRTC1 (Fig. 1) were amplified with PCR using appropriate primers (Table 6). The amplified products were sequenced and then
22
inserted into psiCHECK2 vector (Promega, C8021) and named pCRTC1-P containing the proximal region of 3’UTR and pD containing the distal region of CRTC1-3’UTR respectively. The putative target sequence (GCCCCC) in the distal region of CRTC1-3’UTR of CRTC1 was mutated to TAAACC using specific primers andpCRTC1-D.
11. Luciferase reporter assay
Jurkat cells were resuspended with 100 μl of electroporesis buffer (Invitrogen, MPK10096E) containing 10 ng of luciferase reporter vector and miR-4488 mimic, anti-miR-4488 or corresponding control oligonucleotide. Electroporation was performed using microporator system (Invitrogen Neon, MPK 10096) with pulse voltage of 1200; and pulse width of 40. After 24 h, luciferase activity was measured using the Dual-Luciferase® Reporter 1000 (Promega, E1980, Fitchburg, Wisconsin USA). Briefly, 10 μl of Luciferase Assay Reagent II was added to 20 μl of lysate. The Firefly luciferase activity was measured and then the Renilla luciferase activity by addition of 100 μl of Stop & Glo Reagent. Renilla luciferase activity was normalized to the Firefly luciferase activity.
12. Statistics
23
Fig. 1.Diagram of luciferase reporter vectors containing CRTC-1 3’UTR proximal and distal region, respectively.The location of 4 predicted miR-4488 target sequences was
depicted in 3’UTR of CRTC-1 (vertical Black bar). The sequences of human miR-4488 is aligned with the first predicted target sites in 3’UTR of CRTC1 mRNA. The 4th predicted target sequences were changed into the indicated mutant sequences. The proximal and distal regions which are amplified and inserted into a luciferase reporter vector were denoted as lines with numbers indicating the start and end nucleotide. The transcription start site was set as 1.
24
Table 4.Primers for real-time RT PCR
Primer Neucleotide sequnce
IL-1β (F) TGATGGCTTATTACAGTGGCAATG (R) GTAGTGGTGGTCGGAGATTCG IL-6 (F) AAGCCAGAGCTGTGCAGATGAGTA (R) TGTCCTGCAGCCACTGGTTC TNF-α (F) GACAAGCCTGTAGCCCATGTTGTA (R) CAGCCTTGGCCCTTGAAGA C/EBPα (F) TCGTGGGTCAGCTCTGAGGA (R) GCAATGCTGAAGGCATACAGTACAA C/EBPβ (F) GGCCGGTTTCGAAGTTGATG (R) GCAATGCTGAAGGCATACAGTACAA C/EBPδ (F) CATCGACTTCAGCGCCTACATC (R) AAGAGCTCGTCGTGGCACAG ATF3 (F) AAGAGGCGACGAGAAAGAAA (R) TGGAGTCCTCCCATTCTGAG CRTC1 (F) CTCTTGCGGCATGTGATTTG (R) AGGACACTTGGCTCAGGTGGA CRTC2 (F) AAGAGGCGACGAGAAAGAAA (R) TGGAGTCCTCCCATTCTGAG GAPDH (F) ACAATGTCACCCACCTTGTCTTC (R) GGTCAGTGGACAGTTTACTGGTATC Actin (F) AAGAGGCGACGAGAAAGAAA (R) TGGAGTCCTCCCATTCTGAG
25
Table 5. Sequence of siRNAs
SiRNA Neucleotide sequnce
siC/EBPβ 5’- AGAUGUUCCUACGGGCUUGUUGCTG 5’- CCGUUUCAAGCAUUAAGAACACUTT 5’- AGCACAGCGACGAGUACAAGAUCCG siC/EBPδ 5’- CCUUUGUAUUGUAGAUAAGAGGAAA 5’- CCACUAAACUGCGAGAGAAGCUAAA 5’- GGAGCGCAAAGAAGCUACAGCCUGG siATF3 5’- GCAAUGUACUCUUCCGAUGUUUGTG 5’- ACAGCAUUUAGUGAAGUUGUGCAAC 5’- GCAUUAUUGGAUGUCAAUAGCAUTG
Table 6. Primers for CRTC1 3’UTRamplification
Primer Neucleotide sequnce
CRTC1 3'UTR-P (F) GCCTGTGAGCGGGCACGC
(R) CGGACTCACTTCTTACACTCAGG
CRTC1 3'UTR-D (F) TCGCCCTGAAAACAGCAGCTCCCATCAC (R) AAAAGCAGCCCTGCCAGAGGATGG
26
III. RESULTS
A. Colchicine inhibits caspase-1 activation
A previous study showed that caspase-1 activation mediated by MSU crystals was inhibited by pretreatment of THP1 cells (a monocytic cell line) with colchicine (Martinon F et al., 2006). I examined the inhibitory effect of colchicine on caspase-1 activation induced by ATP treatment or dsDNA transfection, both of which stimulate caspase-1 activation through NLRP3 and AIM2, respectively (Bryant et al., 2009; Fernandes-Alnemri et al., 2009). Colchicine was added to PBMCs isolated from healthy controls 30 min before priming with a low dose [1 ng/ ml] of LPS for 3 h. PBMCs were then stimulated with dsDNA or ATP, and caspase-1 activation was analysed by immunoblotting (Fig. 2). LPS priming alone (albeit weakly), dsDNA transfection and ATP stimulation all induced caspase-1 activation to yield cleaved p20 caspase-caspase-1. This caspase-caspase-1 activation was inhibited by colchicine treatment in all cases.
27
Fig. 2. Cleavage of caspase-1 is inhibited by colchicine. Human peripheral blood
mononuclear cells (PBMCs) were cultured in the presence or absence of colchicine [10 ng/ml]. After 30 minutes Lipopolysaccharide [LPS; 1 ng/ mL] was then added to the cells. Three hours later, PBMCs were stimulated with ATP [5 mM] or transfected with ds DNA [1 μg/mL]. Cells were harvested 1 h after ATP addition or 6 h after transfection for immunoblot analysis of caspase-1 activation (A). Representative immunoblot of three independent experiments with PBMCs from three healthy subjects (B). The relative level of the cleaved caspase-1 (p20) was shown by a chart. The p20 level was normalized to that of α-tubulin. H1, H2, H3, healthy subjects
28
B. Effect of colchicine on the release of interleukin (IL)-1β is dependent on the inflammatory stimulus and responsiveness to colchicine
Caspase-1 is involved in the conversion of pro- IL-1β to its mature form and its subsequent release (Bryant C et al., 2009). To ascertain whether the response to colchicine of patients with BD is associated with suppression of IL-1β release through colchicine mediated inhibition of caspase-1 activation, we assessed the release of IL-1β by PBMCs isolated from patients in the BDR and BDNR groups (Fig. 3). IL-1β release triggered by all stimuli, except dsDNA, was significantly less in the BDR and BDNR groups than in the healthy controls (P<0.005). Unexpectedly, IL-1β release by ‘healthy’ PBMCs stimulated with LPS alone was significantly increased by colchicine treatment; however, no significant alteration in IL-1β release was observed in PBMCs from the BDR and BDNR groups stimulated with LPS alone in the presence of colchicine. When cells were primed with LPS and then stimulated with ATP or dsDNA, IL-1β release increased compared with that in PBMCs stimulated with LPS alone. Colchicine treatment of PBMCs stimulated with ATP or dsDNA did not affect IL-1β release in PBMCs from any of the subjects (controls or patients with BD). The form of IL-1β released in the presence of colchicine was similar to that released in the absence of colchicine, and was detected as a 17-kDa band when immunoblotting the supernatants (data not shown). This is in agreement with a previous study showing no effect of colchicine on IL-1β release by cells stimulated with ATP (Martinon F. et al., 2009). Notably, MSU-driven IL-1β release by healthy PBMCs and PBMCs from the BDR group was significantly suppressed by colchicine treatment in
29
contrast to PBMCs from the BDNR group. Taken together, although caspase-1 activation in PBMCs stimulated with LPS, ATP or dsDNA was suppressed by colchicine, colchicine did not inhibit IL-1β release by PBMCs from the BDR and BDNR groups. In contrast, the effect of colchicine on MSU-driven IL-1β release was associated with colchicine responsiveness in patients with BD. Thus, the effect of colchicine on the release of IL-1β is dependent on both the inflammatory stimulus and responsiveness to colchicine, implying a difference in host cell factor(s) or inflammatory stimuli between BDR and BDNR.
30
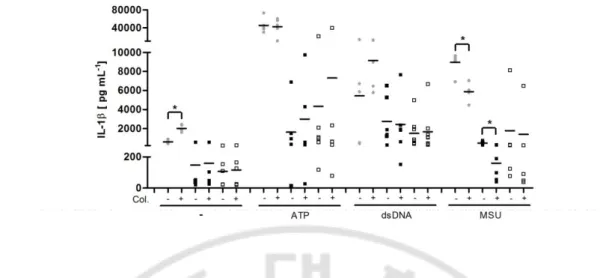
Fig. 3.Effect of colchicine oninterleukin (IL) -1β release.Peripheral blood mononuclear
cells (PBMCs) from healthy subjects (n=5), and Behc¸et disease colchicine responders (n = 7) or nonresponders (n = 8) were cultured in the presence or absence of colchicine [10 ng/ml] for 30 minutes. Lipopolysacchride (LPS) [1 ng/ml] was added to the culture. Three hours later, PBMCs were stimulated with ATP [5 μM] or monosodium urate crystals (MSU) [250 μg/ml] or transfected with ds DNA [1 μg/ml]. Culture supernatants were harvested 3 h after stimulation with LPS alone, 1 h after ATP addition or 6 h after MSU addition or dsDNA transfection. IL-1β was measured by enzyme-linked immunosorbent assay (ELISA). *P < 0.001
31
C. Colchicine does not inhibit lactate dehydrogenase (LDH) release in PBMCs
Caspase-1 activation leads to pyroptosis, which is characterized by formation of pores in the plasma membrane; the extracellular release of the cytoplasmic contents renders pyroptosis inherently proinflammatory (Lamkanfi,2012). Therefore, to ascertain whether inhibition of proinflammatory cell death by colchicine in BD PBMCs is dependent upon their colchicine responsiveness, I assessed the release of LDH (an indicator of pyroptosis) by stimulated PBMCs (Fig. 4). LDH release in response to LPS, ATP and dsDNA was significantly reduced in the BDR and BDNR groups compared with the healthy controls (P<0.005). The data in Figures 2 and 3 suggest the possibility that BD PBMCs may respond less well to LPS and ATP. Contrary to expectation, LDH release in response to LPS alone was significantly increased by colchicine treatment in both healthy controls and the BDR group (> 1.5 fold increase in all subjects); however, colchicine treatment of PBMCs from BDNR in the presence of LPS stimulation alone did not cause a significant change in LDH release (P=0.146). When the cells were primed with LPS and then stimulated with ATP, dsDNA or MSU, no significant alteration in LDH release by colchicine-treated PBMCs from any of the healthy and BD subjects was observed. These results agree with those in a recent report arguing that caspase-1 is capable of inducing necrotic cell death independently of its catalytic activity (Motaniet al., 2011). Taken together, despite inhibiting caspase-1 activation, colchicine may not suppress proinflammatory cell death and may consistently increase LDH release by LPS-stimulated PBMCs isolated from BDR.
32
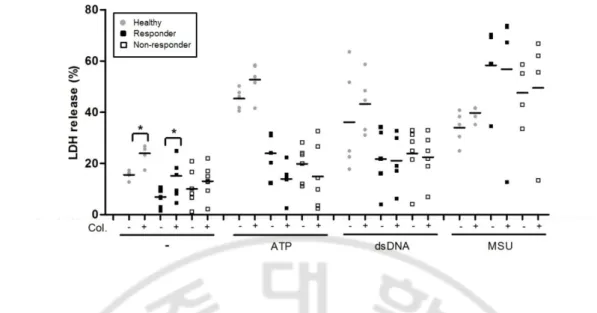
Fig. 4. Lactate dehydrogenase (LDH) release is not inhibited by colchicine. Peripheral
blood mononuclear cells (PBMCs) from healthy subjects (n = 5), and Behc¸et disease colchicine responders (n = 8) or nonresponders (n = 8) were cultured in the presence or absence of colchicine [10 ng/ml]. Lipopolysacchride (LPS) [1 ng/ml] was added to the culture 30 min later. Three hours later, PBMCs were stimulated with ATP [5 μM] or monosodium urate crystals (MSU) [250 μg/ml] or transfected with double-stranded (ds)DNA [1 μg/ml] ). LDH release was analyzed in culture supernatants harvested 3 h after stimulation with LPS alone, 1 h after ATP addition or 6 h after MSU addition or dsDNA transfection. *P< 0.001.
33
D. Colchicine differentially regulates IL-1β transcription in BD PBMCs according to their colchicine responsiveness
Colchicine causes an increase in LPS-stimulated IL-1β mRNA levels and a decrease in TNF-α mRNA levels (Allen et al., 1991). I next examined whether the effect of colchicine on the transcription of proinflammatory cytokines, such as IL-1β, IL-6 and TNF-α might be associated with colchicine responsiveness in patients with BD (Fig. 5). PBMCs were stimulated with LPS in the presence or absence of colchicine and the relative transcript levels of each cytokine were compared. Firstly, the relative IL-1β transcript levels were analyzed against the average IL-1β transcript level in healthy PBMCs not treated with LPS or colchicine (Fig. 5A). IL-1β transcript levels were similar in all subjects in the absence of LPS and colchicine. Upon stimulation with LPS [10 ng/ml], IL-1β transcription was significantly upregulated in BDNR compared with that in BDR (P<0.001). Using similar methods, I next analyzed the transcript levels of TNF-α and IL-6 (Fig. 5C, E). Basal transcript levels of TNF-α and IL-6 were significantly increased in BDNR compared with those in healthy controls (P<0.001). Upon LPS stimulation, transcript levels of TNF-α and IL-6 were significantly higher in BDNR than those in BDR and controls (P<0.001). Also, basal and LPS-induced protein levels were significantly higher in the BDNR group than in the control and BDR groups (Fig. 6 A, C). Interestingly, basal and LPS induced IL-6 production was higher in the BDR group than in the controls. The data in figures 5C and 5E suggest differences in the post-transcriptional regulation of IL-6 expression between the BDR group and healthy controls. These findings were not related to medication or disease
34
activity and lower the possibility that BDNR PBMCs may be less responsive to LPS stimulation. Taken together, the results suggest that expression of proinflammatory cytokines, particularly IL-6 and TNF-α is abnormally increased in PBMCs from BDNR, in contrast to that in BDR.
I next analysed the effect of colchicine on IL-1β, TNF-α and IL-6 transcription (Fig. 5B, D, F). Compared with IL-1β transcript levels in the absence of colchicine, those in the presence of colchicine were similar in controls, significantly decreased in BDNR regardless of LPS stimulation, but significantly increased in BDR stimulated with 10 ng/ mL of LPS (P<0.005). Basal transcription of TNF-α and IL-6 in PBMCs from controls and the BDR group was significantly induced by colchicine treatment, but not in PBMCs from the BDNR group (P<0.005). LPS-stimulated [10 ng/mL] transcription of TNF-α and IL-6 in PBMCs from healthy controls was significantly suppressed by colchicine treatment, but not that in PBMCs from patients with BD (P<0.001). Further, colchicine treatment significantly increased the secretion of these cytokines in BDR, but not in BDNR (Fig. 6 B,D). Although these results are contrary to expectation, they demonstrate that the effect of colchicine treatment on the transcription of IL-1β, TNF-α and IL-6 in BD PBMCs is associated with their responsiveness to colchicine; they also suggest that alterations in IL-1β transcript levels in BD PBMCs stimulated with 10 ng/ml of LPS after colchicine treatment may be a useful indicator for predicting colchicine responsiveness in patients with BD. Also, increasement of transcription of IL-6 by colchicine was demonstrated that colchicine increase IL-6 transcript level with other stimulation in a previous study (Lacroix-Lamandé et al., 2012).
35
Fig. 5.Effects of colchicine on the transcript levels of IL-1β, TNF-αand IL-6 differ in BD PBMCs according to their colchicine responsiveness. PBMCs from healthy subjects
(n = 9), and Behc¸et disease colchicine responders (n = 9) or nonresponders (n=9) were cultured in the presence or absence of colchicine [10 ng /mL]. Lipopolysaccharide (LPS) was added to the culture 30 min later. Three hours later, mRNA levels for the indicated cytokines were analyzed by real-time reverse transcription–polymerase chain reaction and
36
normalized to GAPDH mRNA levels. Fold increase over healthy control [HC(-)]: relative mRNA level in PBMCs under each of the culture conditions vs. the average mRNA level in healthy control PBMCs without LPS stimulation(A,C,E). Fold increase over colchicine [Col.(-)]: relative mRNA levels with colchicine treatment against mRNA levels without colchicine treatment(B,D,F). Each symbol represents each subject and the bars represent the mean of each group. *P< 0.001; **P< 0.005.
37
Fig. 6. The protein levels of TNF-α and IL-6 differ in BDPBMCs according to their colchicine responsiveness. PBMCs from healthy subjects (n = 3), and BDR(n = 5) or
BDNR (n = 5) were cultured in the presence or absence of colchicine [10 ng/mL]. Lipopolysaccharide (LPS) was added to the cultures 30 min later. After a further 3 h, cytokine levels in the culture supernatants were analyzed by enzyme-linked immunosorbent assay. Healthy control samples were assayed in duplicate. Fold increase over healthy control [HC(-)]: relative cytokine levels in PBMCs under each of the culture conditions vs. the average cytokine level in healthy control PBMCs without LPS stimulation(A, C). Fold increase over colchicine [Col.(-)]: relative cytokine levels with colchicine treatment against cytokine levels without colchicine treatment(B, D). Each symbol represents a single subject, and the bars represent the mean for each group. *P< 0.005; **P < 0.05.
38
E. Transcription factors regulating proinflammatory cytokine production are differentially expressed according to colchicine responsiveness
To examine the mechanisms of differential cytokine expression in BD patients according to colchicine responsiveness, I examined the expression of transcription factors such as C/EBPα, C/EBPβ, C/EBPδ and ATF3 which are involved in the regulation of inflammatory cytokines. C/EBPβ and C/EBPδ positively regulate the expression of pro-inflammatory cytokines such as IL-6 and TNF-α but ATF3 negatively regulates. Transcript levels of C/EBPα C/EBPβ, C/EBPδ were significantly up-regulated in BDNR without LPS stimulation (Fig. 7). After LPS stimulation, C/EBPα and C/EBPβ transcript levels were significantly reduced in BDNR (P<0.001) whereas, C/EBPδ transcript levels were significantly further increased in BDNR (P<0.005). ATF3 transcript levels were significantly up-regulated in BDR compared to BDNR and healthy controls in the presence and absence of LPS stimulation. Notably,ATF3 transcript levels were further increased by LPS.
I next compared the protein levels of transcription factors in BD patients and healthy controls by immunoblotting. C/EBPβ transactivation ability is determined by the relative amount of actually present isoforms (LAP/LIP ratio), and the suitable dimerization/interaction partners. The isoforms of C/EBPβ are generated by usage of the differential initiation codon in the single form of C/EBPβ transcript. The C/EBPβ mRNA half-life is roughly 40-60 minutes whereas the half-life of C/EBPβ isoforms about 2 hours for both LAP* and LAP and 8.5 h for LIP. C/EBPβ LAP*(38 kDa) and LAP(35 kDa) induce proinflammatory cytokine expression but LIP(20 kDa) suppress it (Huber R. et al., 2012).
39
As previously reported, LIP isoform was predominantly detected in all subjects by immunoblotting (Fig. 8). Total amount of LAP* and LAP isoforms was relatively increased in BDNR compared to BDR and healthy controls. C/EBPδ expression was increased in BDNR in basal state and slightly increased by LPS stimulation. ATF3 protein levels were similar in BDR and BDNR and not affected by LPS stimulation. These results indicated that expression of proinflammatory cytokine-regulating transcription factors was differentially modulated in BDR and BDNR
40
Fig.7.Transcript levels of various transcription factors regulating the proinflammatory cytokine expression. PBMCs from healthy subjects (n = 5), BDR (n = 5) or BDNR (n = 5)
were cultured in the presence or absence of LPS [10 ng/ml]. After 3 hours, total RNA is isolated and then transcript levels were analyzed by real-time reverse transcription– polymerase chain reaction (RT-PCR) and normalized to actin mRNA. Fold increase over healthy control [HC(-)]: relative mRNA level in PBMCs under each of the culture conditions vs. the average mRNA level in healthy control PBMCs without LPS stimulation. Each symbol represents each subject and the bars represent the mean of each group. *P< 0.001; **P< 0.005.
41
Fig. 8. The protein levels of various transcription factors regulatingthe proinflammatory cytokine expression. PBMCs from healthy subjects (n = 3), BDR (n = 3)
or BDNR (n = 3) were cultured in the presence or absence of LPS [10 ng/ml]. After 3 h, total cell lysates were subjected to western blotting to detect the indicated transcription factors.
42
F. The principal cells abnormally expressing cytokines in BDNR are CD11b+ cells
To find subset population in PBMCs responsible for abnormal cytokine expression in BD, I isolated CD11b- and CD11b+ cells from PBMCs and investigated the expression levels of TNF-α and IL-6 (Fig. 9). TNF-α transcript levels were higher in both CD11b- and CD11b+ cells of BDNR than BDR and healthy controls in the absence of LPS stimulation but further increased in BDNR by LPS stimulation only in CD11b+ cells (Fig. 9A). IL-6 transcript levels of BDNR were higher only in CD11b+ cells stimulated with LPS than that of BDR and healthy controls (Fig. 9C). Moreover, similar to results obtained using PBMCs, the protein levels of TNF-α and IL-6 was increased in CD11b+ cells of BDNR compared to BDR andhealthy controls (Fig. 9 B, D).
Also, the transcript levels of C/EBPα, C/EBPβ, C/EBPδ and ATF3 showed similar patterns to those obtained in PBMCs (Fig. 9 E-H). Transcript levels C/EBPα, C/EBPβ and C/EBPδ were higher in BDNR than BDR and healthy controls both CD11b- and CD11b+ cells regardless of LPS stimulation. In BDNR CD11b+ cells, the reduction of C/EBPα and C/EBPβ transcript levels by LPS stimulation was observed. mRNA levels of ATF3 were increased in BDR compared to BDNR and healthy controls. These results indicate that the main producer of abnormally high proinflammatory cytokines in BDNR might be CD11b+ cells and the abnormal expression of C/EBP might be related with it.
43
Fig. 9. The expression of proinflammatory cytokines and their regulatory transcription factors in CD11b+ and CD11b- PBMCs. CD11b+ and CD11b- cells from healthy subjects (n = 3), BDR (n = 3) or BDNR (n = 3) were isolated and cultured in the presence or absence of LPS [10 ng/ml]. After 3 h, mRNA levels were analyzed by real-time
44
RT-PCR and normalized to actin mRNA levels. Fold increase over healthy control [HC(-)]: relative mRNA level in CD11b+ or CD11b- cells under each of the culture conditions vs. the average mRNA level in healthy control CD11b+ or CD11b-cells without LPS stimulation(A,C,E-H). Protein levels of IL-6 and TNF-α in the culture supernatant were analyzed by ELISA (B,D). Each symbol represents each subject and the bars represent the mean of each group.
45
G. The knocked-down effect of various transcription factors on expression of TNF-α and IL-6 in THP-1 cells
In advance to investigate the role of C/EBPβ and ATF3 on expression of TNF-α and IL-6;In BD CD11b+ cells, the knock-down effect of C/EBPβ, C/EBPδ and ATF3 respectively on the expression of these cytokines was examined using the siRNA in THP-1 cells a human moncytic cell line. The similar transfection efficiency of siRNA was verified by flowcytometric analysis (Fig. 10 A). The reduced expression of transcription factors was confirmed by western blotting (Fig 10 B). THP-1 cells differentiated by PMA [5 ng/ml] stimulation for 24 hwere transfected with the siRNA targeting C/EBPβ(siC/EBPβ), si C/EBPδ and siRNA targeting ATF3 (siATF3), respectively or with the mixture of equal amount of siC/EBPβ and siC/EBPδ. After 24 h, the culture was changed to new media containing LPS or not. After 3 h, transcript levels of TNF-α and IL-6 were analyzed by real-time PCR (Fig. 10 C,E). When siATF3 was transfected, transcript levels of TNF-α and IL-6 were more induced by LPS stimulation in PMA-treated THP-1 cells compared to control oligonuclotide transfection regardless. On the other hand, when siC/EBPβ or siC/EBPδ was transfected, transcript levels of TNF-α and IL-6 were significantly suppressed in THP-1 cells compare to regardless of LPS stimulation. LPS induced protein levels of TNF-α and IL-6 were down-regulated by transfection (Fig. 10 D and F)of siC/EBPβ or siC/EBPδ compared to control oligonucleotide transfection. However, siATF3 tranfection did not induce more production of TNF-α and IL-6 compared to control oligonucleotide transfection.