INTRODUCTION
Organisms live in environments that undergo constant physical and chemical changes, such as changes of tempera-ture, pH, nutrients, and oxidation. In order to survive these changes in the environment, as well as physiological changes, various global regulatory networks have evolved. When sub-jected to environmental stress, organisms undergo funda-mental changes in their cellular physiology and/or morpho-logy, as reflected and directed by changes in the global gene and protein expression patterns, thereby adapting and protec-ting themselves from harmful stresses. These days, various omic analyses such as transcriptome and proteome, which use a DNA microarray, two-dimensional gel electrophoresis (2-DE) coupled with mass spectrometry (MS), and
bioin-formatics, allow researchers to identify up- and down-regu-lated genes in response to a number of chemical and physi-cal stresses, which are physi-called stimulons (Lockhart and Win-zeler 2000; Padney and Mann 2000).
When a cell is subjected to ionizing radiation (IR), differ-ent types of reactive oxygen species (ROS) including supero-xide (O2∙-) and hydroxyl (OH∙) radicals are generated, which are believed to induce oxidative damage in DNA (Riley 1994). This means that exposure of living systems to IR triggers not only an oxidative stress response but also DNA repair systems leading to the elimination of free radi-cals and repair of DNA damage such as SOS response (Symons 1994; Diehl 1995). However, because responses to IR can vary in different cell types and at different radiation doses, or points in time following radiation (Short et al. 2007), it is very hard to identify genuine radiation-inducible genes from many of the genes induced after IR exposure, which seem to be unlinked and individually controlled.
─ ─ 1 ──
Proteome Analysis of Escherichia coli after High-dose Radiation
Sangyong Lim*, Misong Lee1, Minho Joe, Hyunpa Song and Dongho KimRadiation Research Center for Biotechnology, Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
1Department of Biological Science, Seoul National University, Seoul 151-921, Korea
Abstract-- Since proteomics can be employed to compare changes in the expression levels of many proteins under particular genetic and environmental conditions, using mass spectrometry to estab-lish radiation stimulon, we performed two-dimensional gel electrophoresis and identified E. coli proteins whose expressions are affected by high dose of ionizing radiation. After exposure to 3 kGy, it was found that 6 proteins involved in carbon and energy metabolism were reduced. Although 4 of 7 protein spots showing a significant increase in expression level were neither identified nor classified, uridine phosphorylase (Udp), superoxide dismutase (SodB), and thioredoxin-dependent thiol peroxidase (Bcp) were proven to be up-regulated after irradiation. This suggests that E. coli subjected to high doses of radiation (3 kGy) may operate a defense system that is able to detoxify reactive oxygen species and stimulate the salvage pathway of nucleotide synthesis to replenish da-maged DNA.
Key words : Two-dimensional electrophoresis, Ionizing radiation, Escherichia coli
* Corresponding author: Sangyong Lim, Tel. +82-63-570-3141, Fax. +82-63-570-3149, E-mail. [email protected]
Escherichia coli, one of the best-characterized prokaryotes,
has served as a model organism for countless biological studies. Since the genetic information of E. coli is much less complex than that of eukaryotes, several main cellular res-ponses under different conditions including oxidative stress have been well defined (Farr and Kogoma 1991; Han and Lee 2006). Therefore, the use of E. coli as a model system is helpful for comparing an IR response with either an oxi-dative or SOS response. In this study, we have analyzed changes in the protein profile of E. coli after an extremely high dose of radiation at 3 kGy.
MATERIALS AND METHODS
1. Bacterial strains and growth conditionsA wild-type Escherichia coli strain, MC4100 (araD139 ΔargF-lacU169rpsL150 thiA relA1 flb5301 deoC1 ptsF25 rbsR), was used in this study (Shin et al. 2001). Bacteria were routinely cultivated using a shaking period at 37�C in a Luria-Bertani (LB) broth containing 1% tryptone, 0.5% yeast extract, and 1% NaCl.
2. Irradiation
Cultures were irradiated at room temperature using a cobalt-60 gamma irradiator (point source, AECL, IR-79, Ottawa, Canada). The source strength was approximately
400 kCi at a dose rate of 80 Gy min-1, and the actual doses were within ±2% of the target dose. The applied dose of irradiation was 3 kGy.
3. Protein preparation and two-dimensional gel electrophoresis (2-DE)
Cells were harvested and washed with PBS, then resus-pended in a lysis buffer (8 M urea, 2 M thiourea, 2% CHAPS, 1% Pharmalyte, 1% DTT, protease inhibitor). After a 5 min incubation at room temperature, cell debris was removed by centrifugation (10,000×g for 3 min at 4�C), and the clear supernatant was precipitated with 100% acetone for 3 h at -20�C. Protein pellets after centrifugation (12,000×g for 10 min at 4�C) were resuspended in a solubilization buffer (9 M urea, 2% β-mercaptoethanol, 2% Pharmalyte (pH 4~ 7), and 8 mM PMSF) and applied to electrophoresis. Equal amounts of protein from the samples were separated by iso-electric focusing on a pH gradient ranging from 4 to 7. In the second dimension, the proteins were separated according to their molecular mass by sodium dodecyl sulfate-polyacryla-mide gel (SDS-PAGE, 10%) electrophoresis and visualized through colloidal Coomassie brilliant blue (G-250) staining.
4. Protein visualization and image analysis
The protein profiles were compared visually to assign clear differences in intensity using a PDQuest (Bio-Rad) analysis of the gel images. An estimate of the relative
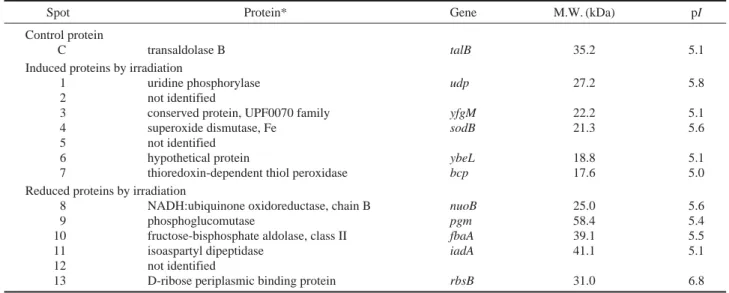
quan-Table 1. Identification of tentative induced and repressed proteins by irradiation
Spot Protein* Gene M.W. (kDa) pI
Control protein
C transaldolase B talB 35.2 5.1
Induced proteins by irradiation
1 uridine phosphorylase udp 27.2 5.8
2 not identified
3 conserved protein, UPF0070 family yfgM 22.2 5.1
4 superoxide dismutase, Fe sodB 21.3 5.6
5 not identified
6 hypothetical protein ybeL 18.8 5.1
7 thioredoxin-dependent thiol peroxidase bcp 17.6 5.0
Reduced proteins by irradiation
8 NADH:ubiquinone oxidoreductase, chain B nuoB 25.0 5.6
9 phosphoglucomutase pgm 58.4 5.4
10 fructose-bisphosphate aldolase, class II fbaA 39.1 5.5
11 isoaspartyl dipeptidase iadA 41.1 5.1
12 not identified
13 D-ribose periplasmic binding protein rbsB 31.0 6.8
*Mass finger prints of numbered protein obtained by MALDI-TOF were analyzed with MS-Fit. The theoretical MW and pI were calculated in ExPASY Proteo-mics Server.
titative changes was determined after normalization against the level of control protein, TalB (Table 1). Protein spots showing more than two-fold differences in the intensity between gel images were digested with trypsin for mass spectrometry analysis.
5. Identification of proteins by mass spectrometry Protein spots showing different levels were excised from stained gels, followed by in-gel digestion. Gel slices were washed twice with water and an acetone-water (1 : 1) solu-tion. To shrink the gel pieces, acetonitrile was added and washed with 0.1 M NH4HCO3. After removal of all liquid by a Speedvac concentrator, the gel pieces were reduced and alkylated with 10 mM dithiothreitol and 55 mM iodoaceta-mide respectively. The gel pieces were dried again, and the protein was digested using trypsin at 37�C overnight in a digestion buffer (50 mM NH4HCO3, 5 mM NaCl, 12.5 mg ml-1trypsin). The volume of the supernatant containing the
generated peptides was reduced by the Speedvac concentra-tor. Peptide mass fingerprinting was done using a Voyager-DETM STR Biospectrometry Workstation (Per-Septive Bio-systems, Framingham, MA, USA), and proteins were identi-fied thorugh peptide mass fingerprinting using the search pro-grams MS-FIT (UCSF Mass Spectrometry Facility at http:// prospector.usuf.edu) and Mascot (Matrix Science, London, UK, at http://www.matrixscience.com) in the NCBInr and Swiss-Prot database.
RESULTS AND DISCUSSION
We compared the protein profiles of E. coli (MC4100) following a dose of 3 kGy, to screen up- and down-regulated genes after IR. Cells were grown up to stationary phase and then subjected to IR. Whole-cell proteins were prepared from cultures immediately before (Fig. 1A) and after IR (Fig. 1B). The expression levels of 13 protein spots showed significant changes: seven spots showed increased expression, whereas six spots showed decreased expression (Table 1). First, the reduced proteins were mainly involved in carbon (pgm, fbaA, and rbsB) and energy metabolism (nuoB). It is evident that 3-kGy radiation causes massive DNA damage in the E. coli genome (Von Sonntag 1987; Symons 1994). Therefore, the surviving fraction of E. coli shows a growth lag, and DNA replication ceases during the lag period to repair DNA da-mage (Khil and Camerini-Otero 2002). In this respect, the down-regulation of genes involved in carbon and energy metabolism can be a way to save energy before the reinitia-tion of replicareinitia-tion and/or cell division. On the other hand, the duration of the lag period of irradiated cells is likely to be more extended due to the reduced expression of IadA, which catalyzes the hydrolysis of dipeptides containing a peptide bond to the β-carboxylate group of aspartic acid. The apparent physiological role of IadA is to prevent the accu-mulation of isoaspartyl dipeptides that could be toxic to the
Fig. 1. Two-dimensional electrophoresis protein profiles of Escherichia coli before (A) and after (B) irradiation. Bacteria were grown
aerobi-cally to stationary phase and then exposed to gamma radiation (3 kGy). Protein spots whose expression was up-regulated or down-regulated are circled. The identities of the proteins indicated by numbered are shown in Table 1.
p/4 7 MW(kDa) MW(kDa) p/4 7 75 50 37 25 20 75 50 37 25 20
A
B
cell or that could partially deplete the pool of utilizable amino acids required for viability (Gary and Clarke 1995).
Among the seven spots induced through 3-kGy radia-tion (Table 1), we could not funcradia-tionally classify four pro-teins because two (spot 2 and 5) were not identified, and others were a hypothetical protein (ybeL) and a protein with unknown function (yfgM). When exposed to IR, bacterial choromosomal DNA is degraded by a process that is pre-sumably initiated by cellular exonucleases at the sites of the DNA-strand breaks (Battista et al. 1999). The degradation of chromosomal DNA is necessarily accompanied by de-novo and salvage syntheses of nucleotides. Uridine phos-phorylase (Udp), which is up-regulated by 3 kGy radiation (Table 1), catalyzes the reversible phosphorylytic cleavage of uridine to uracil and ribose-1-phosphate. The produced uracil is utilized in the rescue of a pyrimidine base, which is involved in the salvage pathway of nucleotide synthesis (Leer et al. 1977; Mushegian and Koonin 1994).
Immediately after IR, ROS such as peroxides, superoxide anion, and hydroxyl radicals, are produced (Riley 1994; Spitz et al. 2004). To prevent the harmful effects of such ROS, cells are equipped with various antioxidant enzymes, including catalases, peroxidases (Prxs), and superoxide dis-mutases (SODs) (Farr and Kogoma 1991). It was found that Fe-containing SOD, which catalyzes the dismutation of su-peroxide into oxygen and hydrogen su-peroxide (H2O2), was induced by 3-kGy radiation (Table 1). Because H2O2is itself a reactive species, cells maintain a variety of antioxidants to remove H2O2. These include catalases, which catalyze the conversion of H2O2to H2O and O2, although it was not ele-vated in this study. A new type of Prx also contributes to the reduction of H2O2and alkyl hydroperoxides with the use of thioredoxin (Trx) and other thiol-containing reducing agents (Cha et al. 1996; Jeong et al. 2000). Escherichia coli has three Prx family members of peroxidases; periplasmic thiol peroxidase (Tpx, p20, scavengase), bacterioferritin-comigratory protein (BCP), and alkyl hydroperoxide reduc-tase peroxidase component (AhpC). Both Tpx and BCP specifically require Trx as a reductant, while AhpC requires AhpF, a type of flavoprotein (Cha et al. 1996; Jeong et al. 2000). We found that the BCP of three Prxs was induced by 3-kGy radiation (Table 1). However, since stress proteins are induced at different times after the onset of stress, we cannot rule out the possibility that catalases encoded by katG or katE are up-regulated following 3-kGy radiation.
A cellular response to extensive DNA damage, which is called SOS response, is concomitantly activated with oxi-dative stress in order to repair the damaged DNA. About 20 genes have been identified in E. coli, including lexA, whose product normally represses SOS genes. The remainder in-cludes genes such as uvrA, uvrB, recA, sulA, and umuC. Among the functions assigned to these genes are recombi-national repair, nucleotide excision repair, inhibition of cell division, and error-prone repair (Sutton et al. 2000). How-ever, no genes involved in SOS response were found in this study (Table 1). This is consistent with the previous reports that treatment with low, but not high, concentrations of H2O2 induces recA, an SOS gene (Farr and Kogoma 1991). In addition, we also were unable to find any typical SOS genes in Salmonella Typhimurium treated with a relatively high dose of 1 kGy radiation (Lim et al. 2008). Taken together, these results strongly suggest that a sudden large increase in ROS generated by a high dose of radiation (3 kGy) may cause massive DNA damage and subsequently invoke a DNA repair system, which is different from a traditional SOS response.
CONCLUSION
In E. coli, we identified up- and down-regulated proteins after a high-dose radiation (3 kGy) through two-dimensional gel electrophoresis. Most down-regulated genes, which are involved in carbon ( pgm, fbaA, and rbsB) and energy meta-bolism (nuoB), are attributed to the growth lag of cells fol-lowing irradiation. In contrast, up-regulated genes including uridine phosphorylase (udp), superoxide dismutase (sodB) and peroxidase (bcp) show that not only detoxification of ROS generated by IR but also nucleotide synthesis occurs actively during the lag period. These pathways appear to act synergistically with SOS response to ensure survival of cells faced with DNA damaging stress such as IR.
ACKNOWLEDGMENTS
This work was supported by the Nuclear R&D program of the Ministry of Education, Science and Technology (MEST), Republic of Korea.
REFERENCES
Battista JR, Earl AM and Park M-J. 1999. Why is Deinococcus radiodurans so resistant to ionizing radiation? TRENDS Microbiol. 7:362-365.
Cha M-K, Kim H-A and Kim I-H. 1996. Mutation and muta-genesis of thiol peroxidase of Escherichia coli and a new type of thiol peroxidase family. J. Bacteriol. 178:5610-5614. Diehl JF. 1995. Biological effects of ionizing radiation. pp. 89-132. In: Safety of Irradiated Foods. Marcel Dekker Inc., New York, NY.
Farr SB and Kogoma T. 1991. Oxidative stress responses in Escherichia coli and Salmonella typhimurium. Microbiol. Rev. 55:561-585.
Gary JD and Clarke S. 1995. Purification and characterization of an isoaspartyl dipeptidase from Escherichia coli. J. Bac-teriol. 270:4076-4087.
Han M-J and Lee SY. 2006. The Escherichia coli proteome: past, present, and future prospects. Microbiol. Mol. Biol. Rev. 70:362-439.
Jeong W, Cha M-K and Kim I-H. 2000. Thioredoxin-dependent hydroperoxide peroxidase activity of bacterioferritin comi-gratory protein (BCP) as a new member of the thiol-specific antioxidant protein (TSA)/alkyl hydroperoxide peroxidase C (AhpC) family. J. Biol. Chem. 275:2924-2930.
Khil PP and Camerini-Otero RD. 2002. Over 1000 genes are involved in the DNA damage response of Escherichia coli. Mol. Microbiol. 44:89-105.
Leer JC, Hammer-Jespersen K and Schwartz M. 1977. Uridine phosphorylase from Escherichia coli: physical and chemical characterization. Eur. J. Biochem. 75:217-224.
Lim S, Jung S, Joe M and Kim D. 2008. Gene expression pro-files following high-dose exposure to gamma radiation in Salmonella enterica serovar Typhimurium. J. Radiat. Ind. 3:111-119.
Lockhart DJ and Winzeler EA. 2000. Genomics, gene expres-sion and DNA arrays. Nature 405:827-836.
Mushegian AR and Koonin EV. 1994. Unexpected sequence similarity between nucleosidases and phosphoribosyltrans-ferases of different specificity. Protein Sci. 3:1081-1088. Padney A and Mann M. 2000. Proteomics to study genes and
genomes. Nature 405:837-846.
Riley PA. 1994. Free radicals in biology: oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 65:27-33. Short SC, Buffa FM, Bourne S, Koritzinsky M, Wouters BG
and Bentzen SM. 2007. Dose- and time-dependent changes in gene expression in human glioma cells after low radiation doses. Radiat. Res. 168:199-208.
Spitz DR, Azzam EI, Li JJ and Gius D. 2004. Metabolic oxida-tion/reduction reactions and cellular responses to ionizing radiation: A unifying concept in stress response biology. Cancer Metast. Rev. 23:311-322.
Sutton MD, Smith BT, Godoy VG and Walker GC. 2000. The SOS response: Recent insights into umuDC-dependent mu-tagenesis and DNA damage tolerance. Annu. Rev. Genet. 34:479-497.
Symons MCR. 1994. Direct and indirect damage to DNA by ionizing radiation. Radiat. Phys. Chem. 43:403-405. Von Sonntag C. 1987. Targets in radiation biology. pp. 94-109.
In: The Chemical Basis of Radiation Biology. Taylor and Francis Ltd., Philadelphia, PA.
Manuscript Received: October 18, 2010 Revision Accepted: November 16, 2010