저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Studies on the Efficient Intracellular
Delivery Vehicle Using Variable Region
Domain of a Cell-penetrating Anti-DNA
Monoclonal Antibody
by
Sun-Woo Im
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
Studies on the Efficient Intracellular
Delivery Vehicle Using Variable Region
Domain of a Cell-penetrating Anti-DNA
Monoclonal Antibody
by
Sun-Woo Im
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements for the
Degree of Master of Biomedical Sciences
Supervised by
Young-Ju Jang, Ph.D.
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
This certifies that the dissertation
of Sun-Woo Im is approved.
SUPERVISORY COMMITTEE
The Graduate School, Ajou University
July, 5th, 2016
Yong-Ju Jang
Sun-Park
-ABSTRACT-
Studies on the Efficient Intracellular Delivery Vehicle Using
Variable Region Domain of a Cell-penetrating Anti-DNA
Monoclonal Antibody
The cell-penetrating antibodies and antibody fragments can be utilized as a tool for the medical diagnosis and treatment. In this study, we have produced the protein of recombinant VH domain of a monoclonal anti-DNA antibody 2C10, IgG of which has been previously shown to have cell-penetrating activities for studying the applicability of the domain as a cellular delivery vehicle. To evaluate the cell-penetrating property of 2C10 VH domain in various mammalian cell lines, the flow cytometric and confocal microscopic analysis and cell proliferation assay were used. The VH domain penetrated into all cell lines we tested in time- and dose-dependent manner, although the internalization efficiency was varied. We also found that the VH domain was localized in nuclei as well as cytoplasm of living cells and it was mainly internalized by the clathrin-mediated endocytosis pathway. The VH domain did not change the cell cycle and significantly affect the cell proliferation in various mammalian cell lines at the concentration that we used for functional assays. We tested further the delivering activity of a valuable molecule by the VH domain using the conjugates of VH and siRNA for testicular Nuclear Auto-antigenic Sperm Protein (tNASP). It was found that the siRNA was successfully transferred by 2C10 VH domain into Caki and HeLa cancer cell lines, and the knockdown effects of tNASP-siRNA were observed in Caki cells, in where
the level of tNASP expression is known to be high; the levels of RNA transcripts and proteins of tNASP were decreased by the delivered tNASP-siRNA; the down-regulated tNASP effectively inhibited cell proliferation and caused G1 phase arrest of cell cycle. These results indicate that the recombinant 2C10 VH domain could be applied as a valuable vehicle which can deliver specific biomolecule(s) into the cytoplasm or cell nuclei for therapy and diagnosis of diseases.
Keywords : Cell-penetrating antibody, Single domain antibody, Cellular delivery vehicle
TABLE OF CONTENTS
ABSTRACT ……….………. i
TABLE OF CONTENTS ……….…... iii
LIST OF FIGURES ……….…... v
I. INTORDUCTION ……….………... 1
II. MATERIALS AND METHODS ……….……....… 3
1. Cell lines …...……….... 3
2. Purification of recombinant 2C10 VH domain .……….……... 3
3. Conjugation of 2C10 VH domain with FITC or si-RNA……….. 4
4. Western blotting ……….………... 4
5. Analysis of cell penetration by flow cytometry .……….……….. 5
6. Analysis of cell penetration by confocal microscopy .……….………. 5
7. Inhibition of cell penetration by endocytosis inhibitors ….………..… 6
8. Gene transcription analysis ………..………. 6
9. Cell proliferation assay ………….……… 6
10. Cell cycle assay ……….……… 7
III. RESULTS ………... 8
1. Expression and purification of recombinant 2C10 VH domain ...……. 8
2. The purified 2C10 VH domain penetrated into various cell lines in a dose-dependent manner ………...………. 8
3. The penetrated 2C10 VH domain was localized in cytoplasm and nucleus of living cells ……...………...………... 13
4. Internalization of 2C10 VH domain into living HeLa cells occurred
mainly via clathrin-mediated endocytosis ……….………. 13
5. 2C10 VH domain did not affect cell proliferation in the various mammalian cell lines and change the cell cycle ………. 19
6. 2C10 VH domain transferred siRNA into mammalian cell lines …... 19
7. Expression of tNASP was decreased in Caki cells by the tNASP-siRNA delivered by 2C10 VH domain …..……….……….... 23
8. The tNASP-siRNA delivered by 2C10 VH domain effectively inhibited cell proliferation and caused G1 phase arrest of cell cycle in Caki cells ………..….………….. 24
IV. DISCUSSION ……….. 28
V. REFERENCES ……….. 31
LIST OF FIGURES
Fig. 1. Purification of 2C10 VH domain ……….…... 9 Fig. 2. Analysis of the penetration of 2C10 VH domain into the various mammalian
cell lines by flow cytometry ………... 10
Fig. 3. Analysis of the penetration of various concentrations of 2C10 VH domain
into HeLa and Caki cells by flow cytometry .………….………... 11
Fig. 4. Analysis of the penetration properties of 2C10 VH domain in Renca cells by
confocal laser microscopy ……….……….…… 12
Fig. 5. Analysis of the kinetics of cell penetration of 2C10 VH domain in Renca
cells by confocal laser microscopy ……….………...………… 15
Fig. 6. Localization of 2C10 VH domain in cell nuclei …………...…...………... 16 Fig. 7. Inhibition of cell penetration of 2C10 VH domain by temperature .…….. 17 Fig. 8. Effect of endocytosis inhibitors on 2C10 VH domain penetration .…....… 18 Fig. 9. Effect of 2C10 VH domain on cell proliferation and cell cycle ……….… 21 Fig. 10. Delivery of siRNA into HeLa cells by 2C10 VH domain ……...….... 22 Fig. 11. Decrease of tNASP expression by the tNASP-siRNA delivered by 2C10
VH domain in Caki cells ...……..………...……… 25
Fig. 12. Decrease of the mRNA level of tNASP by tNASP-siRNA delivered by
2C10 VH daomin in Caki cells ………..……… 26
Fig. 13. Inhibition of cell proliferation and arrest of cell cycle by tNASP-siRNA
I. INTRODUCTION
Antibody (Ab), also known as an immunoglobulin G (IgG), recognizes a unique target molecule on cell surface or extracellular molecule with high affinity (Tansey et al., 1994). In addition, living cell membranes are impermeable to most macromolecules including Ab (Muller et al., 2005; Choi et al., 2014; Weisbart et al., 2012). However, since the first cell-penetrating antibody was reported in 1982 (Alarcon-Segovia et al., 1982), several other penetrating anti-DNA antibodies have been published additionally. These antibodies have been also known that the binding and penetration properties of anti-DNA Abs into living cells could induce the subsequent stimulation of cytokines, cell signals, cell proliferation, pro-inflammatory reaction, cell death and catalytic reaction (Koren et al., 1995; Madaio et al., 1996; Schmidt-Acevedo et al., 2000; Jang et al,. 2009; Lee et al., 2007). Interestingly, the several studies have been focused on characteristics of cell-penetrating antibodies for various researches as well as therapeutic and diagnostic of diseases (Muller et al., 2005; Weisbart et al., 2012; Palanca-Wessels et al., 2016; Frigerio et al., 2013; Thueng-in et al., 2012; Li et al., 2012).
The monoclonal antibodies (mAbs) are monospecific antibodies that specifically bind to one epitope of antigen. Thus, the mAbs are widely used in various fields for mAb-based immunotherapy (Waldmann, 2003; Andrew et al., 2012; Lim et al., 2013; Frigerio et al., 2013). In addition, The mAbs can be engineered to small fragments such as variable region fragment (Fv), fragment antigen-binding (Fab) fragment, and single domain antibody (sdAb) (Nelson, 2010; Kijanka et al., 2015). Antibody fragments are of varied structure, but typically smaller than native antibodies, and lack an Fc domain. Their smaller size offers more binding events per mass of protein purified and administered, less viscous formulations and possibly improved tissue delivery (Rodrigo et al., 2015). The lack of an Fc domain is one reason Ab-fragments exhibit reduced circulation half-lives
as well as immune system activation, which is advantageous in some applications. Especially, the sdAb, which is able to maintain the binding selectivity of a monoclonal antibody IgG to an antigen can be conjugated with a specific molecule and produced in bacterial cells easily (Rodrigo et al., 2015; Ghannam et al., 2015; Nelson, 2010). Consequently, the sdAb could be suitable to use for the delivery vehicle of functional molecules into living cells. Other forms of fragments such as Fab and scFv originated from cell-penetrating Abs have been shown to maintain penetrating activity in mammalian cells (Im et al., 2015; Weisbart et al., 2012; Choi et al., 2014).
In previous studies, we have been reported that anti-double stranded DNA (dsDNA) mAb 2C10 effectively penetrates various mammalian cell lines in vitro and is localized in cytosol as well as nuclei (Im et al., 2015). Therefore, anti-dsDNA mAb 2C10 is a candidate molecule that is able to deliver a specific molecule into living cells. In this study, we developed the recombinant 2C10 VH domain and have explored the possibility of the produced 2C10 VH domain as an intracellular delivery vehicle in mammalian cells. We have used the siRNA of testicular Nuclear Auto-antigenic Sperm Protein (tNASP), which is expressed highly in cancer, germ, transformed, and embryonic cells, for confirming delivery molecules by VH vehicle.
II. MATERIALS AND METHODS
1. Cell lines
Cell lines obtained from the American Type Culture Collection (ATCC, Manassas, VA) include: COS-7 monkey kidney cells; HCT116 human colon carcinoma; Caki human kidney carcinoma; L929 mouse fibrosarcoma; Jurkat human T cell leukemia; HeLa human cervix adenocarcinoma; Renca mouse kidney adenocarcinoma; HaCaT human keratinocyte cells; NIH/3T3 Human embryonic kidney cells; SV40 MES 13 (MES) mouse mesangial cells. COS-7, HCT116, Caki, L929, HeLa, HaCaT and NIH/3T3 cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM, Gibco-BRL, Life Jechnologies, Grand Island, NY, USA), and Jurkat and Renca cells were maintained in Roswell Park Memorial Institute medium (RPMI, Gibco-BRL) and MES cells were maintained in 3:1 mixture of Dulbecco’s Modified Eagle’s Medium and Ham’s F12 medium (Gibco-BRL) with 14mM HEPES. All cells were cultured in growth media that was supplemented with 5% or 10% FBS. Cells were cultured in a 37°C humidified incubator supplied with a mixture of 95% air and 5% CO2.
2. Purification of recombinant 2C10 VH domain
2C10 VH domain was previously constructed as recombinant fusion proteins tagged with five histidines (H5) and a domain of staphylococcal protein A (SPA) (Jang et al., 1996). The protein of 2C10 VH domain was expressed and induced in the bacterial expression host, Escherichia coli BL21 Star (DE3) (Invitrogen, Eugene, OR, USA). Soluble extract from the periplasmic space was
isolated following the methods previously described (Alarcon-Segovia et al., 1982). The expressed soluble VH domain protein was purified using a His-Trap column installed in an AKTA system (GE Healthcare, Little Chalfont, U.K.).
3. Conjugation of 2C10 VH domain with FITC or si-RNA
Purified 2C10 VH domain was conjugated with FITC by Alexa Fluor 488 labeling kit (Invitrogen). 3’-biotin labeled small interference RNA sense strasnd (5’-3’) sequences, ordered form Daramcon (Lafayette, CO, USA), was as follows : GCACAGUUCAGCAAAUCUAdTdT. 3’-biotin labeled tNASP-siRNA was dissolved in RNase-free ddH2O and then mixed with 2C10 VH domain which was labeled streptavidin by Streptavidin Conjugation kit (Abcam, Cambridge, MA, USA) at the molar ratio of 1:2 (2C10 VH:siRNA) and incubated for 40 min at 37◦C.
4. Western blotting
To visualize the purified recombinant 2C10 VH domain, Western blotting with rabbit IgG (Sigma–Aldrich, St. Louis, MO, USA) as the primary detecting antibody for tagged SPA and horseradish peroxidase (HRP)-conjugated anti-rabbit mouse IgG as the secondary antibody (Cell Signaling Technologies, Massachusetts, USA) was used. Specific primary antibodies used for detecting signaling proteins were mouse anti-p-ERK (Cell Signaling Technologies), anti-ERK (Cell Signaling Technologies) and anti-tNASP (Santa Cruz, California, USA). HRP-conjugated anti-mouse goat IgG (Sigma–Aldrich) was used as a secondary antibody. Visualization of protein bands was accomplished using ECL reagents (Amersham Life Science, Amersham, UK).
5. Analysis of cell penetration by flow cytometry
Confluent cells were treated with 0.05% trypsin and 0.02% EDTA for 2 min at 37◦C. The detachment of cells was stopped by adding culture medium. Cells were pelleted by a 3 min centrifugation at 800×g, re-suspended in culture medium and incubated with recombinant 2C10 VH domian or FITC conjugated recombinant 2C10 VH domain at 37◦C for 1 h. Unbound VH domain was removed by washing with phosphate buffered saline (PBS) and the cells were fixed with 4% paraformaldehyde (PFA). To detect VH domain which were penetrated into cells, cells were permeabilized with 0.1% Triton X-100 in PBS for 3 min followed by washing with PBS and incubation with rabbit IgG for 1 h and fluorescein isothiocynate (FITC)-conjugated anti-rabbit IgG (Sigma–Aldrich) for 1 h. Finally, the cells were washed with ice-cold PBS. FITC signal of penetrated VH domain was analyzed by flow cytometry, with counting 100,000 cells for each sample using a FACS Canto II apparatus (Beckton Dickinson) and FlowJo-7 software.
6. Analysis of cell penetration by confocal microscopy
To analyze images of the penetrated 2C10 VH domain, confocal microscopic experiments were performed. Cells were grown to confluence in 12-well plates on which 18 mm cover slips were seated. Cells were incubated with VH domain at 37°C for 1 h or indicated periods, washed in ice-cold PBS and fixed with 4% PFA. Cells were then permeabilized with 0.1% Triton-X 100 in PBS for 3 min followed by washing with PBS. The permeabilized cells were treated with rabbit IgG, followed by treatment with FITC-anti-rabbit IgG. Finally, cells were washed with ice-cold PBS. Nuclei were stained with Hoechst 33342 (Invitrogen) and the specimens were mounted using mounting medium. Images were acquired using a model LSM 510 confocal microscope (Carl Zeiss).
7. Inhibition of cell penetration by endocytosis inhibitors
Confluent cells were pretreated with dansylcadaverine (200 μM) (Sigma– Aldrich), nystatin (50 μg/ml) (Sigma–Aldrich) or Nocodazole (10 μM) (Sigma– Aldrich) under serum-free conditions for 30 min before incubation with FITC-conjugated 2C10 VH domain in the DMEM with 10% FBS for 1 h. Penetration was analyzed by flow cytometry or LSM 510 confocal microscope.
8. Gene transcription analysis
Total RNA was isolated from cultured cells using the RNA-spin Total RNA Extraction Kit (iNtRON, Sungnam, KOREA) and cDNA was synthesized from 200 ng total RNA using the PrimeScript RT reagent kit (TaKaRa, Otsu, Japan). Quantitative PCR (qPCR) was performed using SYBR Premix Ex Taq (TaKaRa) and was analyzed using an ABI 7500 system (Applied Biosystems). The primers for tNASP were GGAGGCTCCAGTTCTCCCTA-3′ (forward), and 5′-AGCTTTGCCCTGACCTTCTC-3′ (reverse). The primers for Beta-glucuronidase
(GUSB) 5′-TCCGTATGTGGATGTGATCTGT-3′ (forward) and
5′-ATCAGAGGTGGATCCTGGTG-3′ (revers) were used as internal normalization controls. Analysis was performed using the 2ΔΔT method (Livak et al., 2001). All reactions were run in triplicate
.
9. Cell proliferation assay
For proliferation assays, cells were seeded into 96-well plates and cultured for 24, 36, 48 and 36 h with VH domain alone or tNASP-siRNA conjugated VH domain. Cell proliferation was determined using MTT reagents (Beyotime,
Shanghai, China) according to the manufacturer’s protocol. Absorbance was detected at the wavelength of 595 nm. Cell viability was measured using at least three wells in each treatment group.
10. Cell cycle assay
Cell were treated with tNASP-siRNA (0.2 μM) alone or VH domain (25 or 50 μg/ml) alone or tNASP-siRNA conjugated 2C10 VH domain (1 μM) for 24, 48 and 60 h. Cells were harvested, washed twice with ice-cold PBS and fixed with 70% ethanol at 4°C overnight. Subsequently, cells were incubated in 50 mg/ml of propidium iodide and 0.5 mg/ml of RNase for 50 min at 37°C. The treated cells were analyzed by flow cytometry (Becton Dickinson, Franklin Lakes, USA).
III. RESULTS
1. Expression and purification of recombinant 2C10 VH domain.
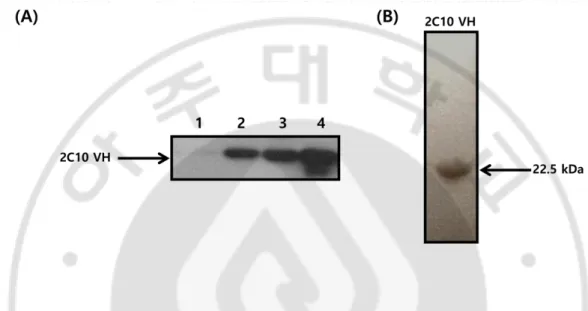
The construct of 2C10 VH domain cloned into the prokaryotic expression vector pIg16 with protein A domain and (His)5 tags in C-terminal was transformed in E. coli BL21(DE) cells for the induction of protein. The soluble VH domain was extracted from periplasmic space and purified by IgG affinity chromatography. Purified VH domain was visualized by Western blotting (Fig. 1A) and Coomassie Brilliant Blue R250 staining (Fig. 1B).
2. The purified 2C10 VH domain penetrated into various cell lines
in a dose-dependent manner.
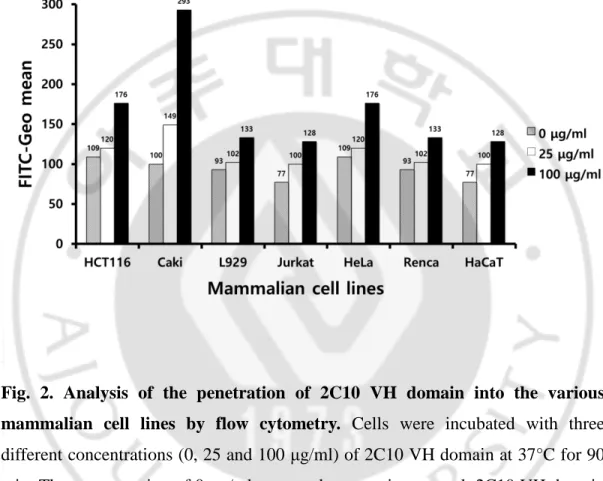
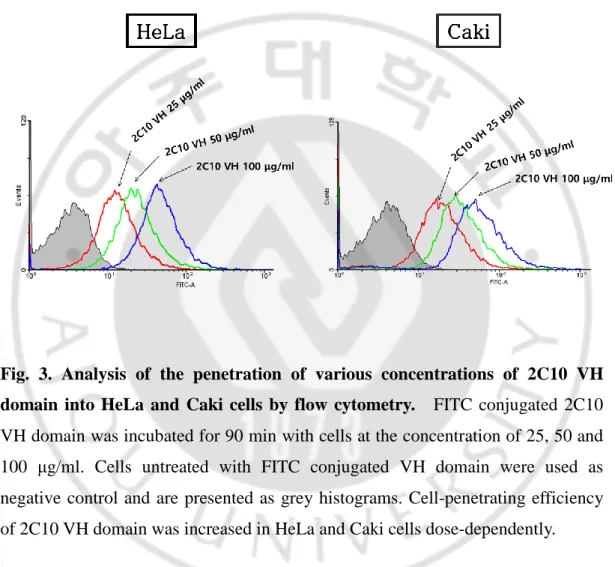
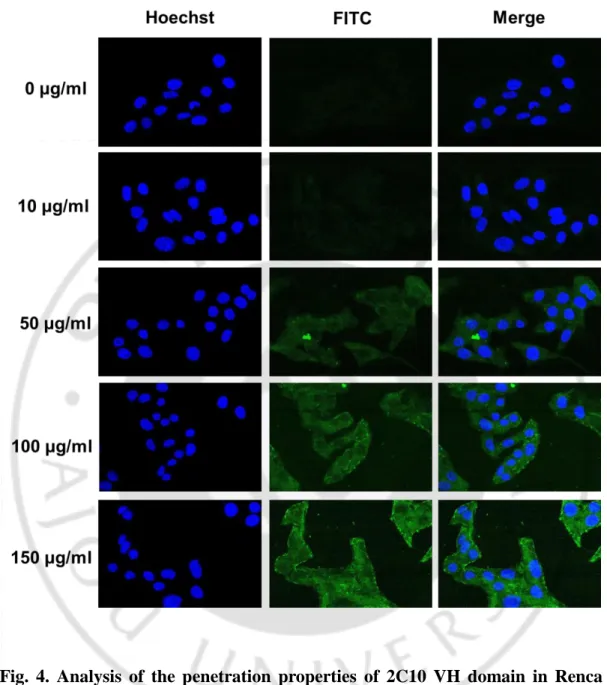
The properties of cell penetration of the 2C10 VH domain were analyzed by flow cytometry in various mammalian cell lines, HCT116, Caki, L929, Jurkat, HeLa, Renca and HaCaT cells, at 25 or 100 μg/ml of VH domain (Fig. 2). After treatment of cells with the VH domain for 1 h at 37°C, cells were fixed and permeabilized for detecting the penetrated VH domain using IgG (rabbit) and α-rabbit IgG-FITC. The VH domain was penetrated into all cell lines, although penetration efficiency of VH domain was varied. To investigate the characteristics of cell-penetrated VH domain, HeLa, Caki, or Renca cells treated with the various concentrations of VH domain conjugated with FITC were analyzed by flow cytometry and/or confocal microscope. The VH domain-FITC was penetrated dose-dependently into HeLa and Caki cells (Fig. 2). Fluorescence images also represented that the VH domain was penetrated concentration-dependently in Renca cells (Fig. 3). These results are consistent with that 2C10 IgG penetrated in a dose-dependent manner into MES cells (Im et al., 2015).
Fig. 1. Purification of 2C10 VH domain. (A) Western blot analysis of purified
2C10 VH domain. Whole cell lysates and purified VH domain (∼1 μg) were separated on 12% SDS-PAGE gels and analyzed by Western blotting using rabbit IgG for the reaction with SPA and HRP-conjugated anti-rabbit IgG. Lane 1 represents; cells untreated with IPTG; lane2; cells induced the expression of VH domain by IPTG; land 3; extracted soluble whole protein in periplasmic space; lane 4; purified VH domain by affinity chromatography. (B) SDS-PAGE and staining of the purified VH domain. Purified VH domain was subjected to 12% SDS-polyacrylamide gel electrophoresis. The protein in the gel was stained with Coomassie brilliant R250. Recombinant 2C10 VH domain was shown at 22.5 kDa, under the non-reducing SDS-PAGE.
Fig. 2. Analysis of the penetration of 2C10 VH domain into the various mammalian cell lines by flow cytometry. Cells were incubated with three
different concentrations (0, 25 and 100 μg/ml) of 2C10 VH domain at 37°C for 90 min. The concentration of 0 μg/ml was used as negative control. 2C10 VH domain was penetrated into the various mammalian cell lines (HCT116, Caki, L929, Jurkat, HeLa, Renca, and HaCaT).
Fig. 3. Analysis of the penetration of various concentrations of 2C10 VH domain into HeLa and Caki cells by flow cytometry. FITC conjugated 2C10
VH domain was incubated for 90 min with cells at the concentration of 25, 50 and 100 μg/ml. Cells untreated with FITC conjugated VH domain were used as negative control and are presented as grey histograms. Cell-penetrating efficiency of 2C10 VH domain was increased in HeLa and Caki cells dose-dependently.
Fig. 4. Analysis of the penetration properties of 2C10 VH domain in Renca cells by confocal laser microscopy. Cells grown on 18-mm cover slips in 12-well
plates were incubated with different concentrations (0, 10, 50, 100 and 150 μg/ml) at 37°C for 1 h. Nucleus was stained with Hoechst (0.5 μg/ml) for 5 min. The fluorescence images were observed using confocal laser microscopy at a magnification of 400×. The concentration of 0 μg/ml was used as a negative control.
3. The penetrated 2C10 VH domain was localized in cytoplasm and
nucleus of living cells.
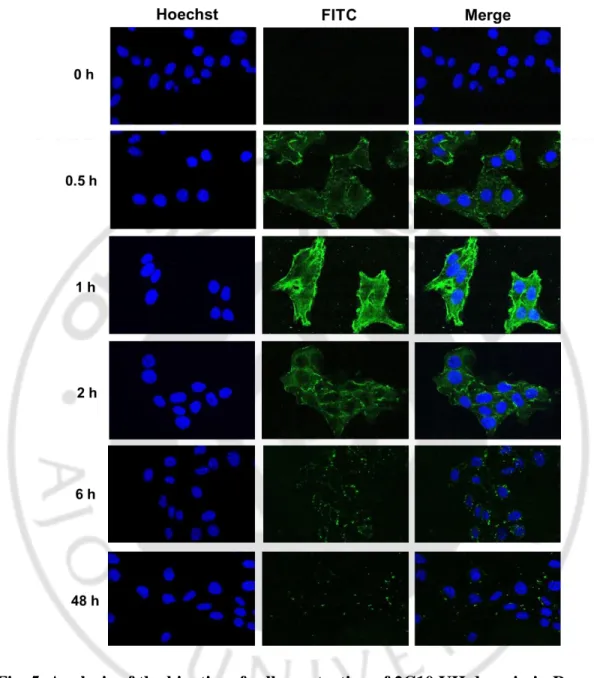
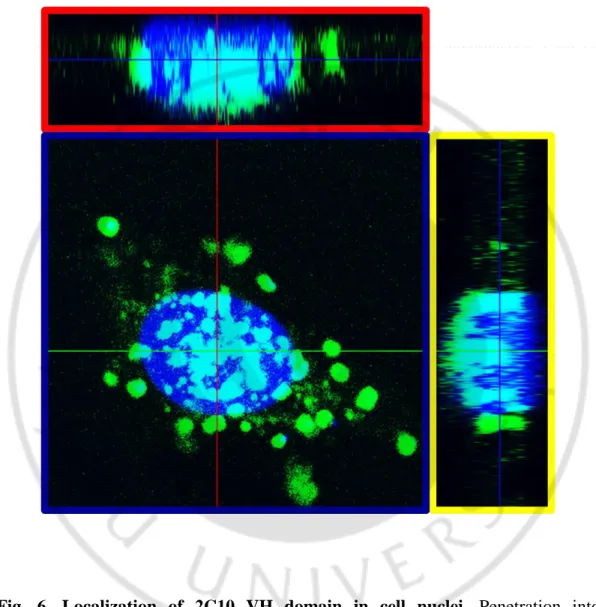
To determine kinetics of cell-penetration of 2C10 VH domain in living cells, the Renca cells were incubated with VH domain at 100 μg/ml for different periods (0, 0.5, 1, 2, 6 and 48 h). Intracellular VH domain was detected with rabbit IgG and FITC-conjugated anti-rabbit IgG and analyzed by confocal microscope. The VH domain was began to be detected from 0.5 h, and the penetration level was increased at 1 h. FITC signal was gradually decreased from 2 h and disappeared at 48 h (Fig. 5). In addition, we analyzed Z-stack of the image of penetration to confirm the intracellular localization of the VH domain. HeLa cells treated with FITC-conjugated VH domain and was stained DAPI for the visualization of cell nuclei. It was shown that the internalized 2C10 VH domain could be localized in cellular nuclei (Fig. 6).
4. Internalization of 2C10 VH domain into living HeLa cells
occurred mainly via clathrin-mediated endocytosis.
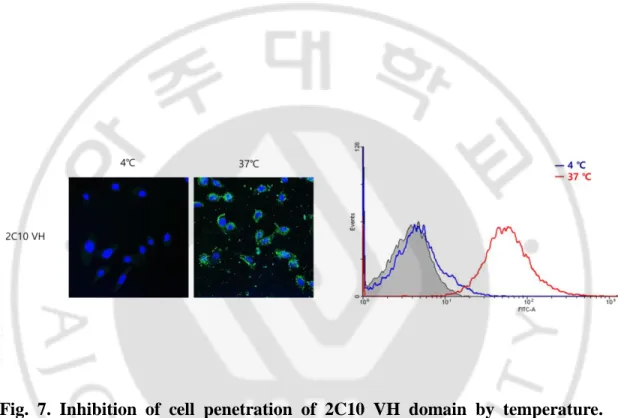
To elucidate the penetration mechanism(s) of 2C10 VH domain, at first we tested temperature dependence of VH domain. Endocytosis is dependent on physical properties of the membrane. In mammalian cells, internalization ceases at 10°C (Silverstein et al., 1977; Dunn et al., 1980). HeLa cells were incubated with FITC-conjugated VH domain at different temperature (37°C or 4°C) and analyzed by confocal microscope and flow cytometry. Penetration of the VH domain into HeLa cells at 37°C was blocked at 4°C (Fig. 7). To confirm that endocytosis is a possible mechanism for cell-penetration of the VH domain, secondly we tested effects of specific inhibitors of endocytosis pathway on the penetration of VH domain. HeLa cells were treated with endocytosis inhibitors (dansylcadaverine,
nystatin, and nocodazole) before addition of FITC-conjugated VH domain. Result of flow cytometry (Fig. 8A) and confocal microscope (Fig. 8B) showed that dansylcadaverine, which is a specific inhibitor of clathrin-mediated endocytosis, significantly (36%) inhibited cellular penetration of the VH domain. Nystatin, which is a specific of caveolae-mediated endocytosis, showed the lower level of inhibition effect (15%) than dansylcadaverine on the cell-penetration. Nocodazole (inhibitor of micropinocytosis) negligibly affected penetration of the VH domain. These results show that cell-penetration of the 2C10 VH domain might occur mainly via clathrin-mediated endocytosis.
Fig. 5. Analysis of the kinetics of cell penetration of 2C10 VH domain in Renca cells by confocal laser microscopy. Cells grown on 18-mm cover slips in 12-well
plates were incubated with 100 μg/ml of 2C10 VH domain at 37◦C for different incubation times. Nucleus was stained with Hoechst (0.5 μg/ml) for 5 min. The fluorescence images were observed using confocal lager microscope at a magnification of 400×. Incubation for 0 h was used as a negative control.
Fig. 6. Localization of 2C10 VH domain in cell nuclei. Penetration into
cytoplasm and nucleus of FITC conjugated 2C10 VH domain was analyzed by confocal microscopy z-stack image. HeLa cells were washed with cold PBS and then treated with 100 μg/ml FITC conjugated VH domain for 1 h. Each Box outlined with red, yellow and blue is section of X-, Y- and Z-axis. Nuclear penetration was observed in three sections.
Fig. 7. Inhibition of cell penetration of 2C10 VH domain by temperature.
Effect of temperature on the penetration of 2C10 VH domain into HeLa cells that incubated with 100 μg/ml FITC conjugated VH domain for 1 h at 4°C or 37°C was examined by confocal microscope and flow cytometry. Each figure shown is a representative of temperature dependence.
Fig. 8. Effect of endocytosis inhibitors on 2C10 VH domain penetration. HeLa
cells were pre-treated with the indicated inhibitors (endocytosis inhibitors: dancycadaverine, nystatin, nocodazole) for 30 min, and then incubated with FITC conjugated 2C10 VH domain for additional 1 h. (A) Fluorescence images were observed using confocal microscope. Untreated and VH domain alone were used as a negative and positive control. Compared to controls, penetration level of VH domain was different in the cells pretreated endocytosis inhibitors. (B) The penetration level of FITC conjugated VH domain in the presence of inhibitors was also measured by flow cytometry. Penetration levels of VH domain are represented as mean percentage (%) by comparing the fluorescence intensity with untreated ‘control’.
(A)
5. 2C10 VH domain did not affect cell proliferation in the various
mammalian cell lines and change the cell cycle.
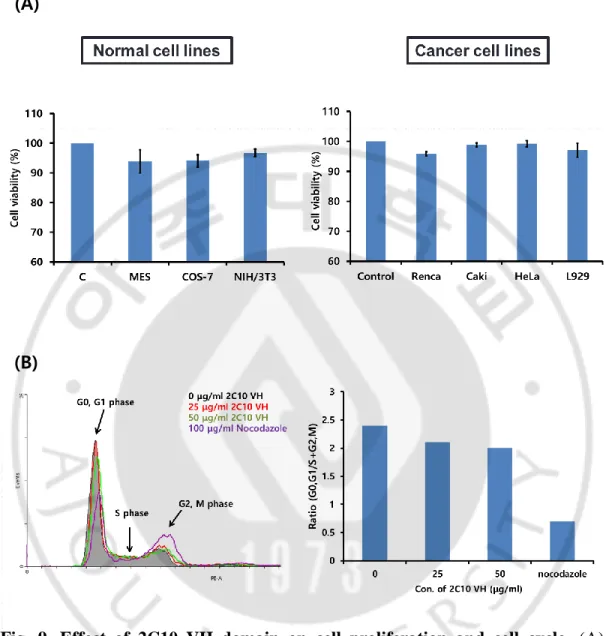
We assessed the effect of the 2C10 VH domain on cell proliferation in various cell lines. Cell proliferation assay was performed in cancer cell lines (Renca, Caki, HeLa and L929) and normal cell lines (MES, COS-7 and NIH/3T3) (Fig. 9A). Treatment of 25 μg/ml VH domain for 24 h did not change cell survivals significantly in all cell lines we tested. Compared to untreated control cells, cell survivals were decreased less than 10% (0.1% ~ 6.1%) in cancer and normal cell lines. To examine whether VH domain affects the phase of cell cycle, Cell cycle of HeLa cells were treated with 25 or 50 μg/ml VH domain for 24 h was also analyzed by flow cytometry (Fig 9B). Ratio of (G0G1/S+G2M) was calculated from percentages of each phase of cell cycle. Nocodazole was used for a control reagent which arrests cell cycle at G2M phase. The ratios of (G0G1/S+G2M) in HeLa cells which were treated with either 25 or 50 μg/ml were negligibly changed, compared to untreated control cells. Contrarily, in cells treated with nocodazole, the ratio was decreased remarkably. These results indicate that 2C10 VH domain does not significantly affect cell proliferation after penetrating target cells.
6. 2C10 VH domain transferred siRNA into mammalian cell lines.
The ability of transferring biomolecules inside living cells by 2C10 VH domain was tested using siRNA for testicular Nuclear Auto-antigenic Sperm protein (tNASP) conjugated with the VH domain. The VH domain was conjugated with streptavidin, and siRNA was labeled with biotin. Streptavidin-conjugated VH domain was mixed with biotin-labeled tNASP-siRNA at a molar ratio of 1:2, and the mixture was incubated at 37°C for 40 min to form a VH-siRNA complex. The resulting mixture was added to HeLa cells in serum-free medium for 2 h. Cells
were stained with FITC labeled anti-rabbit antibody and streptavidin conjugated to PE for image analysis (Fig. 10). It was shown that biotin-labeled tNASP-siRNA alone could not enter the cells. Streptavidin-conjugated VH domain, however, entered the cells as previously reported (Im et al., 2015). Importantly, VH-tNASP-siRNA was localized and visible inside cells. These results indicated that VH-tNASP-siRNA for tNASP was successfully delivered via 2C10 VH domain.
Fig. 9. Effect of 2C10 VH domain on cell proliferation and cell cycle. (A)
Percentage of cell viability was determined by MTT assay in cancer or normal cell lines incubated with 25 μg/ml 2C10 VH domain after 24 h. Untreated was used as a negative control. Cell survivals were decreased less than 10% in cells treated VH domain. (B) Cell cycle distribution was measured by flow cytometry in HeLa cells. Cells were treated with 25 or 50 μg/ml VH domain. Percentages of cell cycle phase are represented as ratio (G0G1/S+G2M). Nocodazole and 0 h were used as a control.
(A)
Fig. 10. Delivery of siRNA into HeLa cells by 2C10 VH domain. Cells grown on
18-mm cover slips in 12-well plates were incubated with 4 μM of 2C10 VH domain conjugated with siRNA at 37◦C for 2 h. Nucleus was stained with Hoechst (0.5 μg/ml) for 5 min. The penetrated VH domain was detected with FITC (green) and transferred siRNA detected with PE (red). The fluorescence images were observed using confocal lager microscope at a magnification of 400×. Untreated was used as a negative control. siRNA conjugated VH domain was detected with red and green.
7. Expression of tNASP was decreased in Caki cells by the
tNASP-siRNA delivered by 2C10 VH domain.
It was well known that NASP expression is a characteristic of all dividing cells, but the splice variant tNASP is presented only in cancer, germ, embryonic and transformed cells at a high level of expression (Richardson et al., 2000). We examined protein level of tNASP in cancer (HeLa, Caki and L929) and normal cell lines (MES and COS-7). As shown in Fig. 11A, protein levels of tNASP in HeLa and Caki cells were higher than normal cells, although the expression level of tNASP was not detectable in L929 cells. To evaluate the knockdown effect of tNASP-siRNA transferred by VH domain in Caki cells, after cells were treated with VH domain alone or tNASP-siRNA transferred by either the VH domain or transfection reagent for the described period, western blotting was performed (Fig. 11B). Decrement of tNASP expression was observed in cells treated with si-tNASP transferred by transfection reagent, which was used for a positive control. Protein level of tNASP was increased at 24 h after treatment with VH-tNASP-siRNA by unknown reason and decreased at 48 h. Expression level of tNASP was not changed in cells treated with the VH domain alone. In addition, phosphorylated ERK, which is known to be a signal molecule and increased in proliferated cells, was observed as decreased at 48 h in cells treated with VH-tNASP-siRNA as well as cells transfected with tNASP-siRNA. The level of tNASP transcript was analyzed by qRT-PCR (Fig. 12). It was significantly decreased in cells treated with VH-tNASP-siRNA, compared to untreated cells and 2C10 VH alone group.
8. The tNASP-siRNA delivered by 2C10 VH domain effectively
inhibited cell proliferation and caused G1 phase arrest of cell cycle
in Caki cells.
To investigate further the effects of tNASP-siRNA transferred inside cells by the 2C10 VH domain. Recent studies were shown down-regulation of tNASP effectively inhibits the proliferation and causes G1 phase arrest in cancer cells (Fang et al., 2015). Therefore, we tested the cell cycle arrest in the G0/G1 phase and inhibition of cell proliferation. The cell cycle distribution was measured by flow cytometry to determine the effect of transferred tNASP-siRNA in Caki and COS-7 cells (Fig. 13A). Compared to untreated control cells, the ratio of (G0,G1)/(S,G2,M) in Caki cells was remarkably increased by VH-tNASP-siRNA. In COS-7, on the other hand, which are normal cells and was shown to have low expression level of tNASP, the transferred si-tNASP did not affect the cell cycle.
In MTT assay, VH-tNASP-siRNA significantly caused the inhibition of the growth of Caki cells at 48h (Fig. 6E). However, inhibition effect of VH-tNASP-siRNA in COS-7 cells was not significant. Meanwhile, the inhibition of cell survival was not high at 24h treated with the VH domain alone, and only 10-20% at 48h in Caki and COS-7 cells. These results indicate that 2C10 VH domain can deliver siRNA to living cells and the transferred siRNA can cause changes of function of cells.
Fig. 11. Decrease of tNASP expression by the tNASP-siRNA delivered by 2C10 VH domain in Caki cells. (A) tNASP protein expression level in various
mammalian cell lines was observed by western blot assay. Cancer and normal cell lines were grown under the same condition for 24 h. Whole cell lysates were separated on 10% SDS-PAGE gels and analyzed by western blotting using anti-tNASP IgG for detecting anti-tNASP inside cells. (B) Knockdown effect of anti- tNASP-siRNA transferred by VH domain was examined in Caki cells. Cells were treated with 0.2 μM tNASP-siRNA or 1 μM VH-tNASP-siRNA or 1 μM VH domain alone and then incubated for the indicted time points. Transfection with tNASP-siRNA and incubation with VH alone were used as a control. Tubulin was examined to verify equal loading.
(A)
Fig. 12. Decrease of the mRNA level of tNASP by tNASP-siRNA delivered by 2C10 VH daomin in Caki cells. Cells were transfected with 0.2 μM
tNASP-siRNA or were treated with 1 μM 2C10 VH-si-tNASP or 1 μM VH alone and then incubated for 36 h. Quantitative real-time RT-PCR was performed with the gene specific primers for tNASP gene. By normalizing to GAPDH genes, results are expressed as fold change untreated control. Asterisks indicate significant differences from respective control groups: *, P<0.05, **, P<0.01, ***, P<0.001.
Fig. 13. Inhibition of cell proliferation and arrest of cell cycle by tNASP-siRNA delivered by 2C10 VH domain. (A) Cell cycle was examined by flow cytometry
in Caki and COS-7 cells. Cells were treated with 0.2 μM tNASP-siRNA or 1 μM tNASP-siRNA conjugated 2C10 VH or 1 μM 2C10 VH. Percentages of cell cycle phase are represented as ratio (G0G1/S+G2M). Untreated was used as a control. (B) Percentage of cell survival was determined by MTT assay in Caki or COS-7 incubated with 1 μM VH domain or 1 μM VM-tNASP-siRNA for described period. Untreated was used as a negative control.
(A)
VI. DISCUSSION
The anti-dsDNA Abs are diagnostic of systemic lupus erythematosus (SLE) and are involved in the pathogenesis of lupus nephritis (Jang et al., 1996; Hahn, 1998; Van der Vlag and Berden, 2011). These Abs can enter living cells and cause inflammatory cascades, proliferation and alteration of cellular functions (Mortensen et al., 2008; Mok and Lau, 2003; Yung and Chan, 2008). The cell penetration property of anti-dsDNA Abs can be utilized as a tool for various purposes. We have previously reported the 2C10 monoclonal antibody effectively penetrated into the cytoplasm and cell nuclei, and many arginine residues in CDR3-VH of 2C10 are implicated in cell penetration ability (Im et al., 2015; Jang et al., 1996; Jang et al., 1990).
In present study, we produced the 2C10 VH domain, which was previously engineered, to use newly as an intracellular delivery vehicle. The recombinant VH domain could enter the various mammalian cells originated from colon, kidney, muscle, lymphocyte, ovary, as well as skin. Penetration of VH domain in HeLa, Caki and Renca cells was dose-dependent. In Renca cells, the VH domain was detected in early time and was shown to be penetrated in time-dependent manner up to 1 h. After 1 h, the FITC signal was significantly decreased and disappeared at 48 h. At 1 h incubated with VH domain, we observed that VH domain was localized in cytoplasm and cell nuclei. These results indicated cell penetration properties and pattern of 2C10 VH domain was similar to those of 2C10 IgG. Meanwhile, the cell penetrating Abs might enter the cells via non-specific charge– charge interactions of the positively-charged amino acid residues in the CDRs with the negatively-charged heparan sulfate proteoglycan (HSPG) on the cell membrane and can be internalized by several mechanisms such as endocytosis pathway (Jang et al., 2009; Lee et al., 2010; Kim et al., 2012; Studzian et al., 2015). Therefore, we tested whether endocytosis was the mechanism for cell-penetration of VH domain.
It was found that clathrin-mediated endocytosis is main mechanism of the cell-penetration of 2C10 VH domain. Detailed fate of the internalized VH should be studied further. The caveolae-mediated endocytosis was also shown to be involved in internalization of VH domain. We also investigated at the VH domain affected cell survival and cell cycle. In cells treated with 1 μM VH domain for 24 h, the VH domain did not affect cell survival rate. In addition, cells treated with the same amount of VH domain did not change the pattern of cell cycle during 24 h in HeLa cells, and similar results were obtained in Caki cells (data not show). In confocal analysis, The VH domain was clearly detected at 1 μM concentration. These findings suggest that less than 1 μM 2C10 VH domain is suitable for being used as a delivery vehicle.
The siRNA interferes with the expression of specific genes and inhibits the corresponding protein expression (Sontheimer, 2005). Recently, siRNA was widely used in the treatment of cells (Song et al., 2003; Urban-Klein et al., 2005; Bummelkamp et al., 2002). However, siRNA alone could not enter into cells due to both the size and the strong anionic charge of phosphate backbone (Reischl et al., 2009; Peixuan et al., 2010). Therefor efficient delivery systems is necessary for siRNA application in clinical. We examined that the VH domain could be used to deliver siRNA for down-regulation of a specific gene. We produced complexes of VH domain and siRNA via streptavidin-biotin interaction. Theses complexes are detected in HeLa cell after incubation for 2 h. Interestingly, activation of ERK, which is increased in proliferated cells, was also restrained in cells, in which tNASP was down-regulated. Recent research showed that knockdown of tNASP would act on ERK signaling pathways to inhibit cell proliferation (Fang et al., 2015). Therefore, our results indicated that the siRNA was successfully transferred by VH domain into Caki and HeLa cancer cell lines. To further investigate the effects of tNASP-siRNA delivered by VH domain, we treated tNASP-siRNA conjugated VH domain in cancer and normal cells. In western blot assay, the
expression levels of tNASP were shown to be different in cancer and normal cells. Recent studies were also showed that NASP is histone H1 binding protein that is involved in transporting histones into the nucleus of dividing cells and testicular form is expressed in embryonic tissues, tumor cells, and the testis (Richardson et al., 2000; Fang et al., 2015; Alekseev et al., 2003). Therefore, the effects of down-regulated tNASP were different in cancer and normal cells. In cancer Caki cells (high expression of tNASP), silence of tNASP caused inhibition of cell proliferation and induced cell cycle arrest in G0/G1 phase. In contrast, transferred tNASP-siRNA negligibly inhibited cell proliferation and did not affect cell cycle in normal COS-7 cells (low expression of tNASP). Meanwhile, recent studies have found that downregulation of tNASP by tNASP-siRNA inhibits cell proliferation via upregulation of P21 and downregulation of cyclineD1 in renal carcinoma cells (Fang et al., 2015; Alekseev et al., 2011). The tNASP-siRNA delivered by VH domain activated ERK signal molecule in Caki cells. These results suggest that recombinant 2C10 VH domain can efficiently deliver siRNA to living cells and the transferred siRNA causes the inhibition of cancer cell proliferation.
In summary, we have successfully developed the delivery system using 2C10 VH domain based on cell penetrating antibody. The VH domain penetrates various cell lines in time- and dose-dependent manner and is localized in nuclei as well as cytoplasm. In addition, the VH domain internalizes into living cells mainly by the clathrin-mediated endocytosis pathway and does not affect remarkably cell proliferation after internalizing target cells. The siRNA is successfully transferred by the VH domain into living cells and siRNA transferred by VH domain can perform the knockdown effect of target gene in cells. Recombinant 2C10 VH domain we established could be applied as a valuable vehicle which can deliver specific biomolecules into the cytoplasm or cell nuclei for therapy and diagnostic of diseases.
V. REFERENCES
1. Alarcon-Segovia D, Llorente L, Ruiz-Arguelles A: Antibody penetration into living cells. I. Intranuclear immunoglobulin in peripheral blood mononuclear cells in mixed connective tissue disease and systemic lupus erythematosus. Clin. Exp. Immunol 23: 22–33, 1982
2. Alekseev OM, Bencic DC, Richardson RT, Widgren EE, O'Rand MG: Overexpression of the Linker histone-binding protein tNASP affects progression through the cell cycle. J Biol Chem. 278(10): 8846-52, 2003 3. Alekseev OM, Richardson RT, Tsuruta JK, O'Rand MG: Depletion of the
histone chaperone tNASP inhibits proliferation and induces apoptosis in prostate cancer PC-3 cells. Reprod Biol Endocrinol., 2011
4. Andrew MS, Jedd DW, Lloyd JO: Antibody therapy of cancer. Nature Reviews Cancer 12: 278-287, 2012
5. Brummelkamp TR, Bernards R, Agami R: Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell. 2(3): 243-7, 2002
6. Choi DK, JeomilBae, Shin SM, Shin JY, Kim SH, Kim Y: A general strategy for generating intact, full-length IgG antibodies that penetrateinto the cytosol of living cells. mAbs 6:1402-1414, 2014
7. Dunn WA, Hubbard AL, Aronson NN: Low temperature selectively inhibits fusion between pinocytic vesicles and lysosomes during heterophagy of 125I-asialofetuin by the perfused rat liver. J. Biol. Chem 255: 5971–78, 1980
Xu A, Liu B, Cao J, Wang Z: Downregulation of tNASP inhibits proliferation through regulating cell cycle-related proteins and inactive ERK/MAPK signal pathway in renal cell carcinoma cells. Tumour Biol. 36(7): 5209-14, 2015
9. Frigerio B, Fracasso G, Luison E, Cingarlin S, Mortarino M, Coliva A, Seregni E, Bombardieri E, Zuccolotto G, Rosato A, Colombatti M, Canevari S, Figini M: A single-chain fragment against prostate specific membrane antigen as a tool to build theranostic reagents for prostate cancer. Eur J Cancer. 49(9): 2223-32, 2013
10. Ghannam A, Kumari S, Muyldermans S, Abbady AQ: Camelid nanobodies with high affinity for broad bean mottle virus: a possible promising tool to immunomodulate plant resistance against viruses. Plant Molecular Biology: 1-15, 2015
11. Hahn BH: Antibodies to DNA. New Engl. J. Med 338: 1359–68, 1998 12. Im SR, Im SW, Chung HY, Pravinsagar P, Jang YJ: Cell- and
nuclear-penetrating anti-dsDNA autoantibodies have multiple arginines in CDR3 of VH and increase cellular level of pERK and Bcl-2 in mesangial cells. Mol Immunol. 67(2 Pt B): 377-87, 2015
13. Jang YJ, Lecerf JM, Stollar BD: Heavy chain dominance in the binding of DNA by a lupus mouse monoclonal autoantibody. Mol. Immunol 33: 197– 210, 1996
14. Jang YJ, Stollar BD: Ultraviolet cross-linking of helical oligonucleotides to two monoclonal MRL-lpr/lpr anti-DNA autoantibodies. Variations in H and L chain binding to DNA. J. Immunol. 145: 3353–59, 1990
acid-hydrolyzing antibody penetrates into cells via caveolae-mediated endocytosis, localizes in the cytosol and exhibits cytotoxicity. Cell. Mol. Life Sci 66: 1985–97, 2009
16. Kijanka M, Dorresteijn B, Oliveira S, van Bergen en Henegouwen PM: Nanobody-based cancer therapy of solid tumors. Nanomedicine (Lond) 10(1):161-74, 2015
17. Kim A, Shin TH, Shin SM, Pham CD, Choi DK, Kwon MH, Kim YS: Cellular internalization mechanism and intracellular trafficking of filamentous M13 phages displaying a cell-penetrating transbody and TAT peptide. PLoS One 7(12): e51813, 2012
18. Koren E, Koscec M, Wolfson-Reichlin M, Ebling FM, Tsao B, Hahn BH, Reichlin M: Murine and human antibodies to native DNA that cross-react with the A and D SnRNP polypeptides cause direct injury of cultured kidney cells. J. Immunol 154: 4857–64, 1995
19. Lee EJ, Jang EJ, Lee E, Yu J, Chung HY, Jang YJ: Cell-penetrating autoantibody induces caspase-mediated apoptosis through catalytic hydrolysis of DNA. Bioorg. Med. Chem 15: 2016–23, 2007
20. Lee WR, Jang JY, Kim JS, Kwon MH, Kim YS: Gene silencing by cell-penetrating, sequence-selective and nucleic-acid hydrolyzing antibodies. Nucleic Acids Res 38: 1596-609, 2010
21. Li T, Bourgeois JP, Celli S, Glacial F, Le Sourd AM, Mecheri S, Weksler B, Romero I, Couraud PO, Rougeon F, Lafaye P: Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: application to brain imaging. FASEB J 26(10): 3969-79, 2012
22. Lim KJ, Sung BH, Shin JR, Lee YW, Kim DJ, Yang KS, Kim SC: A cancer specific cell-penetrating peptide, BR2, for the efficient delivery of an scFv into cancer cells. PLoS One 8(6): e66084, 2013
23. Livak KJ, Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Δ Δ C(T)) method. Methods 25(4): 402-408, 2001
24. Madaio MP, Fabbi M, Tiso M, Daga A, Puccetti A: Spontaneously produced anti-DNA/DNase I autoantibodies modulate nuclear apoptosis in living cells. Eur. J. Immunol 26: 3035–41, 1996
25. Mok CC, Lau CS: Pathogenesis of systemic lupus erythematosus. J. Clin. Pathol. 56 (7): 481–90, 2003
26. Mortensen ES, Fenton KA, Rekvig OP: Lupus nephritis: the central role of nucleosomes revealed. Am. J. Pathol. 172 (2): 275–83, 2008
27. Muller S, Zhao Y, Brown TL, Morgan AC, Kohler H: TransMabs: cell-penetrating antibodies, the next generation. Expert Opin Biol Ther 5: 237– 241, 2005
28. Nelson AL: Antibody fragments: hope and hype. MAbs 2(1): 77–83, 2010 29. Palanca-Wessels MC, Booth GC, Convertine AJ, Lundy BB, Berguig GY,
Press MF, Stayton PS, Press OW: Antibody targeting facilitates effective intratumoral siRNA nanoparticle delivery to HER2-overexpressing cancer cells. Oncotarget 7(8): 9561-75, 2016
30. Peixuan G, Oana C, Nick S, Joe T, Steve H, Songchuan G, Yi S: Engineering RNA for Targeted siRNA Delivery and Medical Application. Adv Drug Deliv Rev. 62(6): 650–666, 2010
31. Reischl D, Zimmer A: Drug delivery of siRNA therapeutics: potentials and limits of nanosystems. Nanomedicine. 5(1): 8-20, 2009
32. Richardson RT, Batova IN, Widgren EE, Zheng LX, Whitfield M, Marzluff WF, O'Rand MG: Characterization of the histone H1-binding protein, NASP, as a cell cycle-regulated somatic protein. J Biol Chem 275(39): 30378-86, 2000
33. Rodrigo G, Gruvegård M, James M., Van Alstine: Antibody Fragments and Their Purification by Protein L Affinity Chromatography. Antibodies 4(3): 259-277, 2015
34. Schmidt-Acevedo S, Pérez-Romano B, Ruiz-Argüelles A: LE cells result from phagocytosis of apoptotic bodies induced by antinuclear antibodies. J. Autoimmun 15: 15–20, 2000
35. Silverstein SC, Steinman RM, Cohn Annu ZA: Endocytosis Rev. Biochem 46: 669–722, 1977
36. Song E, Lee SK, Wang J, Ince N, Ouyang N, Min J, Chen J, Shankar P, Lieberman J: RNA interference targeting Fas protects mice from fulminant hepatitis. Nat Med. 9(3): 347-51, 2003
37. Sontheimer EJ: Assembly and function of RNA silencing complexes. Nat Rev Mol Cell Biol.6(2): 127-38, 2005
38. Studzian M, Bartosz G, Pulaski L: Endocytosis of ABCG2 drug transporter caused by binding of 5D3 antibody: trafficking mechanisms and intracellular fate. Biochim. Biophys. Acta 1853: 1759–71, 2015
39. Tansey EM, Catterall PP: Monoclonal antibodies: a witness seminar in contemporary medical history. Med Hist. Jul 38(3): 322–327, 1994
40. Thueng-in K, Thanongsaksrikul J, Srimanote P, Bangphoomi K, Poungpair O, Maneewatch S, Choowongkomon K, Chaicumpa W: Cell penetrable humanized-VH/V(H)H that inhibit RNA dependent RNA polymerase (NS5B) of HCV. PLoS One 7(11): e49254, 2012
41. Urban-Klein B, Werth S, Abuharbeid S, Czubayko F, Aigner A: RNAi-mediated gene-targeting through systemic application of polyethylenimine (PEI)-complexed siRNA in vivo. Gene Ther. 12(5):461-6, 2005
42. Van der Vlag J, Berden JH: Lupus nephritis: role of antinucleosome autoantibodies. Semin. Nephrol. 31: 376–389, 2011
43. Waldmann TA: Immunotherapy: past, present and future. Nature Medicine 9(3): 269–277, 2003
44. Weisbart RH, Gera JF, Chan G, Hansen JE, Li E, Cloninger C, Levine AJ, Nishimura RN: A Cell-Penetrating Bispecific Antibody for Therapeutic Regulation of Intracellular Targets. Mol Cancer Ther 11(10): 2169–73, 2012
45. Yung S, Chan TM: Anti-DNA antibodies in the pathogenesis of lupus nephritis--the emerging mechanisms. Autoimmun Rev 7 (4): 317–21, 2008
- 국문요약 -
세포투과성 항-DNA 단클론 항체의 가변부위 도메인을 이
용한 효율적인 세포내 물질 전달 도구에 관한 연구
아주대학교 대학원 의생명과학과 임 선 우 ( 지도교수 : 장 영 주 ) 세포투과 항체의 특성을 활용하면 병의 진단과 치료의 도구로서 활용될 수 있다. 따라서, 본 연구의 목적은 세포 내로 침투하는 특성을 가진 항-DNA 단클론 항체 (2C10)에서 유래한 재조합 2C10 VH domain 의 살아 있는 세포내 물질 운반체로의 기능과 유용성을 확인하는 것이다. 재조합 2C10 VH domain의 세포투과 특성을 다양한 포유동물 세포주에 서 flow cytometry, confocal microscopy, cell proliferation assay를 통하여 분석하였다. VH domian은 비록 세포주마다 세포를 투과하는 정도는 다 르지만 분석한 모든 세포주에 시간과 투여량에 의존적인 투과를 보였다.또한 2C10 VH domain이 살아있는 세포 속에서 세포질에 분포할 뿐만 아니라 일부는 핵까지 들어가는 것을 관찰하였다. 2C10 VH domain의 세 포투과기전을 밝히기 위하여 endocytosis inhibitor를 사용한 결과, 주로 clathrin-mediated endocytosis를 통하여 세포 내로 투과하는 것을 확인 하였다. 2C10 VH domain의 세포 침투는 25 μg/ml 농도에서도 유의성 있
게 관찰되었으며, 이 농도에서는 cell cycle과 cell proliferation에 큰 영향 을 주지 않는 것을 확인하였기 때문에 다른 실험의 기본 농도로 정하여 수행하였다. 2C10 VH domain이 중요한 생물학적 기능을 가진 분자를 세 포내로 운반할 수 있는지 확인하기 위하여, VH domain에 암세포의 증식 에 연관되어 있다고 알려진 tNASP(testicular Nuclear Auto-antigenic Sperm Protein)를 타겟으로 하는 소간섭 알앤에이(siRNA)에 biotin을 결 합시킨 후, streptavidin을 결합시킨 2C10 VH와 복합체를 형성시킨 후 사 용하였다. VH domain에 의하여 운반된 tNASP-siRNA에 의하여 세포 내 의 tNASP 단백질의 발현 정도와 전사체(transcripts)의 양이 낮아졌으며, cell proliferation이 감소하였고, cell cycle이 G0, G1 phase에서의 arrest가 유도 되는 것을 Western blot 및 정량적 RT-PCR 분석과 flow cytometry, MTT assay 그리고 confocal microscope 이미지 분석을 통하여 확인하였
다. 이러한 결과들을 통하여, 항체공학을 이용하여 생산된 항-DNA 단클론 항체 2C10 유래 VH domain이 주요한 기능을 지닌 특정 분자를 세포내로 전달할 수 있는 운반체로서의 기능을 할 수 있으며, 다양한 생 명과학 및 의학 분야에 활용될 수 있는 가능성을 지님을 제시할 수 있다. 핵심어 : 세포투과 항체, Single domain antibody, Cellular delivery vehicle