저작자표시-비영리-동일조건변경허락 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. l 이차적 저작물을 작성할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 동일조건변경허락. 귀하가 이 저작물을 개작, 변형 또는 가공했을 경우 에는, 이 저작물과 동일한 이용허락조건하에서만 배포할 수 있습니다.
i - ABSTRACT -
Association of the Interleukin-1 beta and Interleukin-1 Receptor
Accessory Protein Gene Polymorphisms with Persistent Hepatitis B
Virus Infection and Hepatocellular Carcinoma
Background & Aims: The reasons for persistent hepatitis B virus infection (HBV) are
unknown, but they are probably related to host immune factors. Interleukin-1β (IL-1β) plays significant roles in inflammation and immune defense via IL-1 receptor accessory protein (IL-1RAcP). We investigated whether genetic polymorphisms of IL-1B and IL-1RAcP genes are associated with persistent HBV infection and the presence of hepatocellular carcinoma (HCC).
Methods: We enrolled a total of 292 patients with chronic HBV infection (111 with chronic
hepatitis, 95 with liver cirrhosis and 86 with HCC) and 107 healthy individuals who recovered from HBV infection. We assessed 28 SNPs in IL-1B and IL-1RAcP genes by using Illumina’s Sentrix array matrix chip.
Results: Four SNPs of IL-1B (-2023G>C, -581T>C, 289T>C, 3340A>G) and six SNPs of
the IL-1RAcP (-51668T>A, -8261T>C, -8183A>G, -256C>T, 59264G>A, 65445A>G) showed biallelic polymorphism. IL-1B -2023 C allele, IL-1RAcP -8261 T allele and -8183 A allele were significantly associated with persistent HBV infection (OR, 1.63, P = 0.03, OR, 0.64, P <0.01 and OR, 0.20, P = 0.01, respectively). IL-1B 289 C allele was marginally
ii
associated with an increased risk for the presence of HCC (OR, 1.55, P = 0.04). On the haplotype analysis, IL-1B -2023C/-581C/2893C haplotype and IL-1RAcP -8261T/-8183A haplotype were associated with persistent HBV infection. There is no significant association between the haplotypes of IL-1B/IL-1RAcP and the presence of HCC.
Conclusions: The genetic polymorphisms of IL-1B -2023 C allele, IL-1RAcP -8261 T allele
and -8183 A allele are probable host factors for persistent HBV infection.
Keywords : Chronic hepatitis B, Hepatocellular carcinoma, Interleukin-1β, Interleukin-1 receptor accessory protein, Single nucleotide polymorphism
iii
TABLE OF CONTENTS
ABSTRACT ··· ⅰ TABLE OF CONTENS ··· ⅱ LIST OF FIGURES ··· ⅲ LIST OF TABLES ··· ⅳ . Ⅰ INTRODUCTION ··· 1 . Ⅱ MATERIALS AND METHODS··· 3A. MATERIALS··· 3
1. Patients ··· 3
2. DNA preparation ··· 5
B. METHODS ··· 5
1. Selection of the candidate genes (IL-1B and IL-1RAcP) and relevant single nucleotide polymorphisms ··· 5
2. Genotyping of IL-1B and IL-1RAcP SNPs in the Korean patients ··· 8
3. Statistical analyses··· 9 . Ⅲ RESULTS ··· 10 . Ⅳ DISCUSSION ··· 17 . Ⅴ CONCLUSION ··· 21 REFERENCES ··· 22 국문요약 ··· 27
iv
LIST OF FIGURES
v
LIST OF TABLES
Table 1. General characteristics of the study subjects ··· 4
Table 2. Frequencies of IL-1B and IL-1RAcP genotypes and Hardy-Weinberg
equilibrium ··· 11
Table 3. Logistic analysis of IL-1B and IL-1RAP polymorphisms with HBV
Persistence ··· 13
Table 4. Logistic analysis of IL-1B and IL-1RAcP polymorphisms with the presence of HCC ··· 14
Table 5. Logistic analysis of IL-1B and IL-1RAP Haplotype with HBV persistence ··· 16
- 1 -
I. INTRODUCTION
Hepatitis B virus (HBV) infection is one of the most important chronic liver diseases in the world, especially in several areas of Asia and Africa. More than 350 million people in the world are persistently infected with HBV and are at high risk of developing chronic hepatitis, cirrhosis and hepatocellular carcinoma (HCC) (Lee, 1997). The age at the time of infection is a well-known determinant of the progression to chronicity (Tassopoulos, et al., 1987). However, the underlying mechanisms for the persistence of HBV infection or the occurrence of HCC are not fully understood.
Single nucleotide polymorphisms (SNPs) are one of the most commonly occurring forms of genetic variation in the human genome. The SNPs of interleukin-10 and tumor necrosis factor (TNF)-α genes have been reported to be associated with persistence HBV infection (Hohler, et al., 1998; Cheong, et al., 2006). Several SNPs in various genes such as the osteoponin, insulin-like growth factor II and transforming growth factor ß1 genes have been associated with the risk of developing HCC (Kim, et al., 2003; Kim, et al., 2006; Shin,
et al., 2007). Thus, specific SNPs may promote the chronicity of HBV infections and the
subsequent development of HCC.
Interleukin-1 (IL-1) is one of the proinflammatory cytokines, and it has a central role in inflammation and destruction (Arend, et al., 2008). IL-1 family has three ligands and these are interleukin-1α (IL-1α), interleukin-1ß (IL-1ß), and interleukin-1 receptor antagonist 1ra). The receptors for the IL-1 family include type I, II IL-1 receptors (IL-1RI and II) and IL-1R accessory protein (IL-1RAcP). After IL-1 binds to IL-(IL-1RI, a second chain called the IL-1RAcP joins with IL-1/IL-1RI to form a complex, and leading to cell
- 2 -
activation mediated by the cytoplasmic domains of both receptor chains (Wesche, et al., 1997; Arend, et al., 2008). It has been recently reported that human HBV e antigen (HBeAg) can trigger the host IL-1 response by binding to IL-1RAcP and this interaction may play an important role in modulating the host immune response in patients with acute and chronic HBV infections (Yang, et al., 2006).
Several polymorphisms of IL-1B gene that are thought to affect IL-1ß production have been reported. In Caucasians who are infected with Helicobacter pylori, IL1B 31C/1L1B -511T haplotype and IL-1RN (encodes IL-1ra) allele 2 (IL-1RN*2) have been associated with an increased risk of gastric cancer (El-Omar, et al., 2000; 2001). IL-1B -31 T/T genotype or
IL-1B -31T/-511C haplotype was associated with the presence of HCC in Japanese patients
with chronic HCV infection (Wang, et al., 2003). The subjects with IL-1B -511 genotype C/C were closely related with HBV-DNA replication (Zhang, et al., 2004). However, the role of the IL-1RAcP polymorphisms has never been investigated.
In the present study, we investigated whether the genetic polymorphisms of 1B and
IL-1RAcP have any associations with persistent HBV infection and the presence of HCC in the
- 3 -
II. MATERIALS AND METHODS
A. MATERIALS
1. Patients
All the patients were unrelated ethnic Koreans who attended outpatient clinics of Gastroenterology Department or the Center for Health Promotion of Ajou University Hospital in Suwon, South Korea. Between June 2000 and February 2006, a total of 399 patients were enrolled. Informed consent was acquired from each patient; the sampling protocols we used and the informed consent form were approved by the Institutional Review Board of Ajou University Hospital. The patients who are positive for antibody to hepatitis C virus were excluded beforehand. The patients were then divided into four groups according to their HBV infection status, clinical data and serological profiles as 1) the spontaneously recovered group, 2) the chronic hepatitis group. 3) the liver cirrhosis group and 4) the HCC group. The spontaneously recovered group (acute self-limited HBV-infected patients or those with HBV eradication) included 107 subjects (M = 80, F = 27, mean age: 46 years) who were hepatitis B surface antigen (HBsAg) negative, antibody to hepatitis B core antigen (anti-HBc IgG) positive and antibody to hepatitis B surface antigen (anti-HBs) positive. These subjects who visited the Center for Health Promotion of Ajou University Hospital during the period of the study served as a control group. The chronic hepatitis group included 111 patients with chronic hepatitis B (M = 82, F = 29, mean age: 40 years), and they were diagnosed based on positivity for HBsAg and HBV DNA for more than 6 months, and they had an elevated alanine aminotransferase (ALT) level at least once during the
- 4 -
follow-up period. These patients had normal platelet counts and no evidence of cirrhosis by imaging studies. Ninety-five patients were diagnosed as having liver cirrhosis (M = 76, F = 19, mean age: 45 years) based on the typical morphological findings on computed tomography or ultrasound, and they had evidence of portal hypertension or the corresponding laboratory features, including low platelet counts and synthetic dysfunction. The HCC group included 86 HCC patients (M = 69, F = 17, mean age: 53 years) who were diagnosed on the basis of computed tomography, ultrasonography, magnetic resonance imaging and angiography combined with a level of alpha-fetoprotein (AFP) of ≥400 ng/ml. Until December 2004, HBV-DNA was measured by the Digene hybrid capture II assay and we have used the branched DNA (bDNA) assay (VersantTM 3.0, Bayer Healthcare LLC
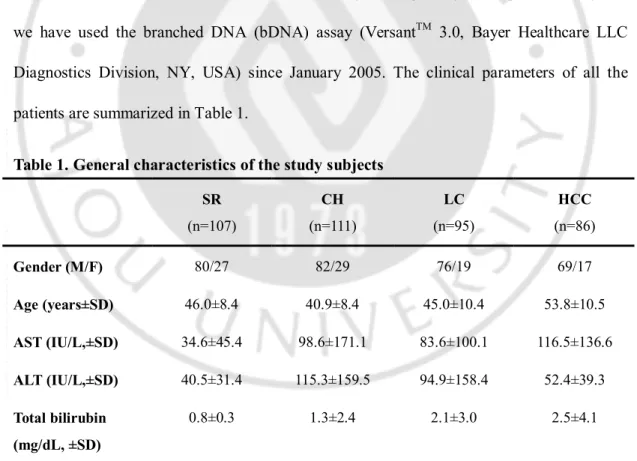
Diagnostics Division, NY, USA) since January 2005. The clinical parameters of all the patients are summarized in Table 1.
Table 1. General characteristics of the study subjects
SR (n=107) CH (n=111) LC (n=95) HCC (n=86) Gender (M/F) 80/27 82/29 76/19 69/17 Age (years±SD) 46.0±8.4 40.9±8.4 45.0±10.4 53.8±10.5 AST (IU/L,±SD) 34.6±45.4 98.6±171.1 83.6±100.1 116.5±136.6 ALT (IU/L,±SD) 40.5±31.4 115.3±159.5 94.9±158.4 52.4±39.3 Total bilirubin (mg/dL, ±SD) 0.8±0.3 1.3±2.4 2.1±3.0 2.5±4.1
SR Spontaneously recovered group, CH Chronic hepatitis B, LC Liver cirrhosis, HCC Hepatocellular carcinoma, SD standard deviation
- 5 -
2. DNA preparation
The blood samples were stored at -80℃ until use. The genomic DNA was purified using G-DEXTM blood genomic DNA purification kits (Intron Biotechnology Inc., Seongnam, South Korea) and the genomic DNA was quantified using the Picogreen double stranded DNA quantification reagent according to the following standard protocol (Molecular Probes, Eugene, OR, USA). A Picogreen working solution was prepared by making 1:200 dilutions in TE buffer (pH 8.0). The λ DNA standard and samples were mixed with the Picogreen working solution and then they were incubated for 5 min at room temperature. The plates were read using a VictorTM 3 multilabel counter (excitation 480 nm
and emission 520 nm; PerkinElmer Inc., Waltham, MA, USA). A standard DNA concentration curve was calculated by using the known concentrations of λ DNA.
B. METHODS
1. Selection of the candidate genes (IL-1B and IL-1RAcP) and
relevant single nucleotide polymorphisms
IL-1B and IL-1RAcP genes were selected for further analysis from the gene expression
data obtained by comparing the HCC tissues and adjacent non-tumor tissues with using the GenePlorer TwinChip Human-8K human cDNA microarray (Digital Genomics, Seoul, Korea). The IL-1B mRNA levels were more highly expressed in the chronic hepatitis and cirrhotic tissues than the control tissues. The IL-1RAcP mRNA levels were more highly expressed in the chronic hepatitis, liver cirrhosis and HCC tissues than that in the control tissues (data not shown). The validated SNPs in the IL-1B and IL-1RAcP genes were
- 6 -
selected from a public SNP database (http://www.ncbi.nlm.nih.gov/projects/SNP/). As shown in Figure 1, there were 15 IL-1B SNPs and 13 IL-1RAcP SNPs (Fig. 1A and 1C).
A.
- 7 -
C.
- 8 -
Fig 1. Gene map, SNPs and pairwise LD of the IL-1B and IL-1RAcP genes. A. Gene map
and SNPs of IL-1B gene. IL-1B showing the locations of the 15 known SNPs. Exons are shown as black boxes, the 5’ and 3’ UTRs are represented by light grey boxes and the introns and 5’ and 3’ flanking regions are marked by grey and black lines, respectively. †, polymorphic SNP site, ‡, monomorphic or low frequency (MAF <0.05) SNP site B. LD blocks and haplotypes of IL-1B gene. The LD blocks in the spontaneously recovered group (SR), the chronic hepatitis + liver cirrhosis (CLD; chronic liver disease) and hepatocellular carcinoma group (HCC) are displayed in dark gray with using |D’|=1 (Haploview software; cut off values MAF=0.05 and HWE=0.05). C. The gene map and SNPs of IL-1RAcP. IL-1B showing the locations of the 13 known SNPs. D. The LD blocks and haplotypes of the
IL-1RAcP gene.
2. Genotyping of IL-1B and IL-1RAcP SNPs in the Korean
patients
Genotyping was performed using a Golden gate genotyping assay kit according to a standard protocol (Illumina Inc., San Diego, CA, USA). Briefly, 250 ng of genomic DNA were mixed with oligomers, and allele-specific extension was carried out by decreasing the temperature from 70 ℃ to 30 ℃ over a 16 h period. The specific extension products were then used in the polymerase chain reactions (PCRs), which consisted of 34 cycles for 35 s at 95 ℃, 35 s at 56 ℃ and 2 min at 72 ℃. The PCR products were purified using 96-well filter plates (Millipore, Billerica, MA, USA). For hybridization, all the samples were
- 9 -
transferred to a 384-wellmicroplate. The Sentrix array matrix (SAM) chip and the purified PCR products were hybridized at 60 ℃ for 30 min and then at 45 ℃ for 16 h. The SAM chip was then washed and imaged at a resolution of 0.8mm by using a BeadArray Reader (Illumina Inc., San Diego, CA, USA). Genotyping analysis was performed using Illumina’s BEADSTUDIO software (Version 3.0.22). The other reagents were purchased from Sigma-Aldrich (St Louis, MO, USA).
3. Statistical analyses
We performed χ2 tests to check for deviations from the Hardy–Weinberg equilibrium
(HWE). The genetic models for the association tests were divided into the co-dominant, dominant and recessive models. A multiple logistic regression model was used to calculate the odds ratios (OR), the 95% confidence intervals (CI) and the P values, after controlling for age and gender as covariates, with using SAS statistical software (SAS 9.1 and SAS Enterprise Guide 4.1; SAS Institute, Cary, NC, USA). A linkage disequilibrium (LD) block of SNPs was confirmed using HAPLOVIEW software (version 4.0; http://www.broad.mit.edu/mpg/haploview) (Fig 1B. and 1D). The individual haplotypes were inferred by the EM algorithm and with using the SAS haplotype procedure. Haplotype analyses were done by performing multiple logistic regression analyses. For all the statistical tests, significance was set at P <0.05. For more precise estimates, statistical computation was performed to determine the P values of Fisher’s exact test by Monte Carlo simulation (number of iterations = 100,000).
- 10 -
III. RESULTS
A. The genotypes and allele frequencies
IL-1B gene is located on chromosome 2q14 and we selected 15 known IL-1B SNPs
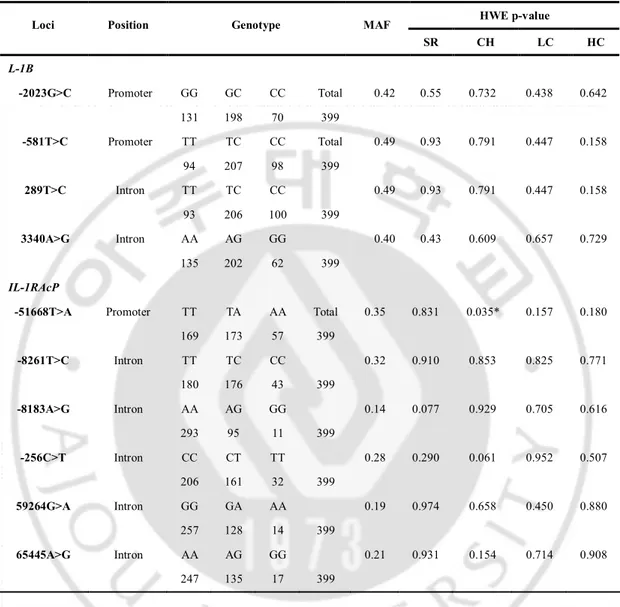
from dbSNP database. Of theses, seven were located in the flanking 5’UTR, five were in introns, two were in the 3’UTR and one was in the flanking 3’UTR (Fig. 1A). IL-1RAcP gene is located on chromosome 3q28 and we selected 13 known 1L-1RAcP SNPs. Of these, eight were located in introns, four were in the flanking 5’UTR and one was in the 3’UTR (Fig. 1C). Four IL-1B SNPs and six IL-1RAcP SNPs were polymorphic [i.e., a minor allele frequency (MAF) of >0.05] in the spontaneously recovered group, which represents the general Korean population (Fig. 1A, 1C and Table 2). The frequencies of these SNPs in the spontaneously recovered group, the chronic hepatitis group, the liver cirrhosis group and the HCC group are shown in Table 2. The remaining eleven IL-1B SNPs and seven IL-1RAcP SNPs were monomorphic or had a low MAF (<0.05). The genotype frequencies of these ten SNPs did not deviate from those expected under Hardy-Weinberg equilibrium (HWE) at each locus in the Korean population (P >0.05; HWE in Table 2), except at IL1RAcP -51668T>A SNP in the chronic hepatitis group (P = 0.035).
- 11 -
Table 2. Frequencies of IL-1B and IL-1RAcP genotypes and Hardy-Weinberg equilibrium HWE p-value
Loci Position Genotype MAF
SR CH LC HC L-1B -2023G>C Promoter GG GC CC Total 0.42 0.55 0.732 0.438 0.642 131 198 70 399 -581T>C Promoter TT TC CC Total 0.49 0.93 0.791 0.447 0.158 94 207 98 399 289T>C Intron TT TC CC 0.49 0.93 0.791 0.447 0.158 93 206 100 399 3340A>G Intron AA AG GG 0.40 0.43 0.609 0.657 0.729 135 202 62 399 IL-1RAcP
-51668T>A Promoter TT TA AA Total 0.35 0.831 0.035* 0.157 0.180
169 173 57 399 -8261T>C Intron TT TC CC 0.32 0.910 0.853 0.825 0.771 180 176 43 399 -8183A>G Intron AA AG GG 0.14 0.077 0.929 0.705 0.616 293 95 11 399 -256C>T Intron CC CT TT 0.28 0.290 0.061 0.952 0.507 206 161 32 399 59264G>A Intron GG GA AA 0.19 0.974 0.658 0.450 0.880 257 128 14 399 65445A>G Intron AA AG GG 0.21 0.931 0.154 0.714 0.908 247 135 17 399
SR spontaneously recovered group, CH chronic hepatitis B, LC liver cirrhosis, HCC hepatocellular carcinoma, MAF minor allele frequency, HWE Hardy-Weinberg equilibrium, *p<0.05.
- 12 -
B. The allelic and genotype associations with persistent HBV
infection and the presence of HCC
To determine whether specific IL-1B SNP or IL-1RAcP SNP alleles are associated with persistence of HBV infection, we performed genotypic association analyses by presuming there are two genetic models, namely, the spontaneously recovered group and the others, and then we performed logistic regression analyses. The allele frequencies and association test results are summarized in Table 3. Of the ten polymorphic Korean 1B SNPs and
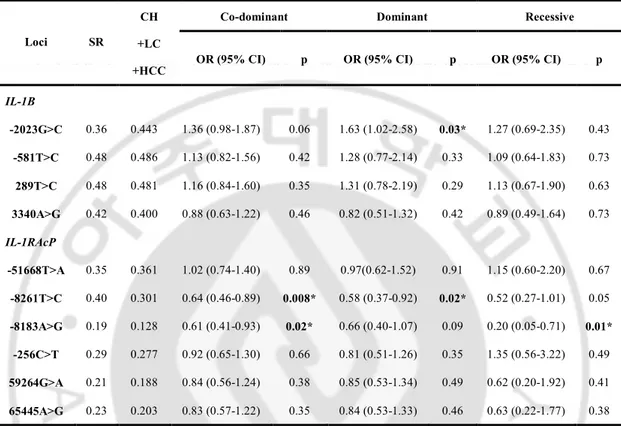
IL-1RAcP SNPs, three SNPs were associated with persistent HBV infection. IL-1B -2023 C
allele was associated with an increased risk of a persistent HBV infection in the dominant model (OR = 1.63; 95% CI = 1.02-2.58; P = 0.03). IL-1RAcP -8261 T allele was associated with HBV persistence in the co-dominant and dominant models (OR = 0.64; 95% CI = 0.460.89; P = 0.008 and OR=0.58; 95% CI = 0.370.92; P = 0.02, respectively). And IL1RAcP -8183 A allele was associated with an increased risk of a persistent HBV infection in the co-dominant and recessive models (OR = 0.61; 95% CI = 0.41-0.93, P = 0.02 and OR = 0.20; 95% CI = 0.05-0.71; P = 0.01, respectively).
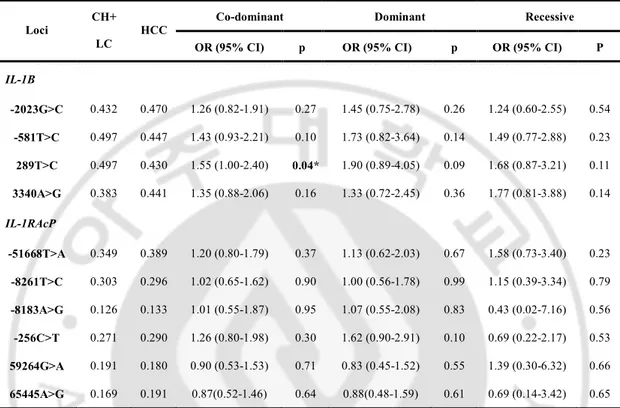
To determine whether a specific IL-1B SNP or the IL-1RAcP SNP alleles are associated with the presence of HCC, we performed other genetic model association analyses and logistic regression analyses between the chronic liver disease group [chronic hepatitis and liver cirrhosis] and the HCC group. One allele of IL-1B SNPs was associated with an increased risk of HCC presence. IL-1B 289 C allele was associated with an increased risk of HCC being present in only the co-dominant model (OR = 1.55; 95% CI = 1.00-2.40; P = 0.04).
- 13 -
Table 3. Logistic analysis of IL-1B and IL-1RAP polymorphisms with HBV
persistence
Co-dominant Dominant Recessive
Loci SR
CH +LC
+HCC OR (95% CI) p OR (95% CI) p OR (95% CI) p
IL-1B -2023G>C 0.36 0.443 1.36 (0.98-1.87) 0.06 1.63 (1.02-2.58) 0.03* 1.27 (0.69-2.35) 0.43 -581T>C 0.48 0.486 1.13 (0.82-1.56) 0.42 1.28 (0.77-2.14) 0.33 1.09 (0.64-1.83) 0.73 289T>C 0.48 0.481 1.16 (0.84-1.60) 0.35 1.31 (0.78-2.19) 0.29 1.13 (0.67-1.90) 0.63 3340A>G 0.42 0.400 0.88 (0.63-1.22) 0.46 0.82 (0.51-1.32) 0.42 0.89 (0.49-1.64) 0.73 IL-1RAcP -51668T>A 0.35 0.361 1.02 (0.74-1.40) 0.89 0.97(0.62-1.52) 0.91 1.15 (0.60-2.20) 0.67 -8261T>C 0.40 0.301 0.64 (0.46-0.89) 0.008* 0.58 (0.37-0.92) 0.02* 0.52 (0.27-1.01) 0.05 -8183A>G 0.19 0.128 0.61 (0.41-0.93) 0.02* 0.66 (0.40-1.07) 0.09 0.20 (0.05-0.71) 0.01* -256C>T 0.29 0.277 0.92 (0.65-1.30) 0.66 0.81 (0.51-1.26) 0.35 1.35 (0.56-3.22) 0.49 59264G>A 0.21 0.188 0.84 (0.56-1.24) 0.38 0.85 (0.53-1.34) 0.49 0.62 (0.20-1.92) 0.41 65445A>G 0.23 0.203 0.83 (0.57-1.22) 0.35 0.84 (0.53-1.33) 0.46 0.63 (0.22-1.77) 0.38
Genotype distributions and P-value for logistic analysis of three alternative models (co-dominant, dominant and recessive models) controlling for age and sex as covariates are shown. SR spontaneously recovered group, CH chronic hepatitis B, LC liver cirrhosis, HCC hepatocellular carcinoma, OR odds ratio, 95% CI 95% confidence interval, * p<0.05.
- 14 -
Table 4. Logistic analysis of IL-1B and IL-1RAcP polymorphisms with the presence of HCC
Co-dominant Dominant Recessive
Loci CH+
LC
HCC
OR (95% CI) p OR (95% CI) p OR (95% CI) P
IL-1B -2023G>C 0.432 0.470 1.26 (0.82-1.91) 0.27 1.45 (0.75-2.78) 0.26 1.24 (0.60-2.55) 0.54 -581T>C 0.497 0.447 1.43 (0.93-2.21) 0.10 1.73 (0.82-3.64) 0.14 1.49 (0.77-2.88) 0.23 289T>C 0.497 0.430 1.55 (1.00-2.40) 0.04* 1.90 (0.89-4.05) 0.09 1.68 (0.87-3.21) 0.11 3340A>G 0.383 0.441 1.35 (0.88-2.06) 0.16 1.33 (0.72-2.45) 0.36 1.77 (0.81-3.88) 0.14 IL-1RAcP -51668T>A 0.349 0.389 1.20 (0.80-1.79) 0.37 1.13 (0.62-2.03) 0.67 1.58 (0.73-3.40) 0.23 -8261T>C 0.303 0.296 1.02 (0.65-1.62) 0.90 1.00 (0.56-1.78) 0.99 1.15 (0.39-3.34) 0.79 -8183A>G 0.126 0.133 1.01 (0.55-1.87) 0.95 1.07 (0.55-2.08) 0.83 0.43 (0.02-7.16) 0.56 -256C>T 0.271 0.290 1.26 (0.80-1.98) 0.30 1.62 (0.90-2.91) 0.10 0.69 (0.22-2.17) 0.53 59264G>A 0.191 0.180 0.90 (0.53-1.53) 0.71 0.83 (0.45-1.52) 0.55 1.39 (0.30-6.32) 0.66 65445A>G 0.169 0.191 0.87(0.52-1.46) 0.64 0.88(0.48-1.59) 0.61 0.69 (0.14-3.42) 0.65
Genotype distributions and P-value for logistic analysis of three alternative models (co-dominant, dominant and recessive models) controlling for age and sex as covariates are shown. SR spontaneously recovered group, CH chronic hepatitis B, LC liver cirrhosis, HCC hepatocellular carcinoma, OR odds ratio, 95% CI 95% confidence interval, * p<0.05.
- 15 -
C. Haplotype association with persistent HBV infection and the
presence of HCC
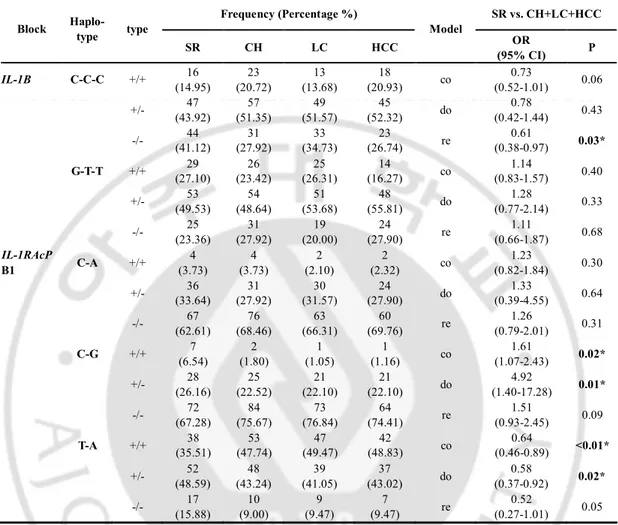
Linkage disequilibrium (LD) blocks were constructed by the Gabriel method and with using HAPLOVIEW software (cut-off values: HWE=0.001 and MAF=0.001, spontaneously recovered group=107, chronic liver disease=206, HCC group=86) (Fig. 1B and 1D). IL-1B had one LD block that comprised -2023, -581 and 289. This block had three haplotypes, the most common of which was ht1 (G-T-T), followed by ht2 (C-C-C) and rare ht3 (G-C-C).
IL-1RAcP had two LD blocks, that is, block 1 (B1) that is comprised -8261 and -8183, while the
LD block 2 (B2) consists of 59264 and 65445. IL-1RAcP B1 had three haplotypes, and these were B1ht1 (T-A), B1ht2 (C-G), and B1ht3 (C-A), and B2 had two haplotypes, B2ht1 (G-A) and B2ht2 (A-G). The haplotype frequencies and the association test results are summarized in Table 4. On the haplotype analysis, we found that IL-1B ht2 (C-C-C) was associated with an increased risk of persistent HBV infection in only the recessive model (OR = 0.61; 95% CI = 0.38-0.97; P = 0.03). IL-1RAcP B1ht1 (T-A) was associated with an increased risk of persistent HBV infection in the co-dominant model and the dominant model (OR = 0.64; 95% CI = 0.46-0.89; P <0.01 and OR = 0.58; 95% CI = 0.37-0.92; P = 0.02, respectively). In contrast, IL-1RAcP B1ht2 (C-G) was associated with viral clearance in the co-dominant model and the dominant model (OR = 1.61, 95% CI = 1.07-2.43; P = 0.02, and OR = 1.92; 95% CI = 1.40-17.28; P = 0.01 respectively). There was no significant haplotype associated with the presence of HCC.
- 16 -
Table 5. Logistic analysis of IL-1B and IL-1RAP Haplotype with HBV persistence
Frequency (Percentage %) SR vs. CH+LC+HCC Block Haplo- type type SR CH LC HCC Model OR (95% CI) P IL-1B C-C-C +/+ 16 (14.95) 23 (20.72) 13 (13.68) 18 (20.93) co 0.73 (0.52-1.01) 0.06 +/- 47 (43.92) 57 (51.35) 49 (51.57) 45 (52.32) do 0.78 (0.42-1.44) 0.43 -/- (41.12) 44 (27.92) 31 (34.73) 33 (26.74) 23 re (0.38-0.97) 0.61 0.03* G-T-T +/+ (27.10) 29 (23.42) 26 (26.31) 25 (16.27) 14 co (0.83-1.57) 1.14 0.40 +/- 53 (49.53) 54 (48.64) 51 (53.68) 48 (55.81) do 1.28 (0.77-2.14) 0.33 -/- (23.36) 25 (27.92) 31 (20.00) 19 (27.90) 24 re (0.66-1.87) 1.11 0.68 IL-1RAcP B1 C-A +/+ 4 (3.73) 4 (3.73) 2 (2.10) 2 (2.32) co 1.23 (0.82-1.84) 0.30 +/- (33.64) 36 (27.92) 31 (31.57) 30 (27.90) 24 do (0.39-4.55) 1.33 0.64 -/- 67 (62.61) 76 (68.46) 63 (66.31) 60 (69.76) re 1.26 (0.79-2.01) 0.31 C-G +/+ (6.54) 7 (1.80) 2 (1.05) 1 (1.16) 1 co (1.07-2.43) 1.61 0.02* +/- 28 (26.16) 25 (22.52) 21 (22.10) 21 (22.10) do 4.92 (1.40-17.28) 0.01* -/- (67.28) 72 (75.67) 84 (76.84) 73 (74.41) 64 re (0.93-2.45) 1.51 0.09 T-A +/+ 38 (35.51) 53 (47.74) 47 (49.47) 42 (48.83) co 0.64 (0.46-0.89) <0.01* +/- 52 (48.59) 48 (43.24) 39 (41.05) 37 (43.02) do 0.58 (0.37-0.92) 0.02* -/- (15.88) 17 (9.00) 10 (9.47) 9 (9.47) 7 re (0.27-1.01) 0.52 0.05
Genotype distributions and P-value for logistic analysis of three alternative models (co-dominant, dominant and recessive models) controlling for age and sex as covariates are shown. SR spontaneously recovered group, CH chronic hepatitis B, LC liver cirrhosis, HCC hepatocellular carcinoma, OR odds ratio, 95% CI 95% confidence interval, co: co-dominant model, do: dominant model, re: recessive model, * p<0.05.
- 17 -
IV. DISCUSSION
In the present study, we found that IL-1B and IL-1RAcP gene polymorphisms were associated with a persistent HBV infection. IL-1B -2023 C allele and IL-1B CCC haplotype that contains -2023 C allele were associated with persistent HBV infection. IL-1RAcP -8261 T allele and -8183 A allele were also associated with persistent HBV infection. The haplotype IL-1RAcP B1ht1 (T-A) that consisted of -8261 and -8183 showed linkage with persistent HBV infection. Concerning the presence of HCC, IL-1B 289 C allele was marginally associated with an increased risk of the HCC presence in only the co-dominant model. Any significant haplotype associated with the increased risk for HCC was not found.
The clearance of HBV requires a coordinated effort by the innate and adaptive immune responses. The innate proinflammatory cytokine IL-1ß initiates a wide spectrum of immunological responses to infection, stress and tissue damage (Dinarello, 1996). IL-1ß attenuates the interferon-induced antiviral activity and it also functions as a costimulator to activate Th2 cells (Tian, et al., 2000; Wang, et al., 2003). Binding of IL-1 to its receptor (IL-1RI) leads to recruitment of the membrane form of IL-1RAcP (mIL-1RAcP), which is essential for signal transduction. One recent study have shown that HBeAg can bind to mIL-1RAcP on the cell surface, and this induces the association of IL-1RI and mIL-mIL-1RAcP, which triggers the recruitment of adaptor protein myeloid differentiation factor 88 to the 1RAcP complex (Yang, et al., 2006). Assembly and activation of the IL-1RI/mIL-1RAcP signaling complex by HBeAg can activate the downstream nuclear factor –kappa B pathway through I kappa B degradation, and this then induces the expression of IL-1 responsive genes, including the genes for IL-1ß, IL-6, TNF-α, inducible nitric oxide
- 18 -
synthase and granulocyte macrophage colony stimulating factor (Yang, et al., 2006). This result raised the possibility that the IL-1ß and IL-1RAcP may contribute to the control of HBV replication during a natural course of HBV infection.
With regard to hepatocellular carcinogenesis, IL-1ß up-regulates mitogen-inducible nitric oxide (NO) and cyclooxygenase (COX) 2. The combined expression of inducible NO and COX-2 may have an important effect on the development of HCC by modulating angiogenesis (Rahman, et al., 2001). In addition, recent evidence has also shown that IL-1ß prevents apoptosis in many cell types, including keratinocytes, chondrocystes, osteoclasts, neutrophils, monocytes and lymphocytes (Kothny-Wilkes, et al., 1998; Watson, et al., 1998; Schmidt, et al., 1999; Kühn, et al., 2000; Lee, et al., 2002). It has been recently reported that the highest IL-1ß mRNA levels are observed in cirrhotic tissues surrounding HCC, suggesting that the consequent chronic inflammation may contribute to inducing the oncogenic mutations that are responsible for initiating or promoting primary liver cancer (Bortolami, et al., 2008).
IL-1B gene was reported to be a possible candidate gene for an increased risk of
persistent HBV infection and fibrosis. Zhang et al. (Zhang, et al., 2004) reported that 1B-511 CC genotype was closely related with HBV-DNA replication. In our previous study,
IL-1B-511 T allele was marginally related to clearance of HBV infection in the Korean patients
with chronic hepatitis B (P =0.049, unpublished data). In addition, the IL-1B -31 T allele was found to be implicated in liver fibrogenesis of patients with chronic HBV infection through the regulation of matrix metalloproteinase production by the hepatic satellite cells(Migita, et al., 2007). IL-1RAcP has been reported to play an important role in modulating the host immune response to HBV infection. However, any data on the
- 19 -
association of the IL-1RAcP polymorphisms and chronic HBV infection has not yet reported. Because our previous study showed only a marginal association between IL-1B -511 T allele and HBV infection in the Korean population, we further evaluated the association of the other SNPs of IL-1B and IL-1RAcP with persistent HBV infection and HCC. Our study showed IL-1B -2023 C allele and its haplotype CCC were associated with a persistent HBV infection (OR = 1.63; 95% CI = 1.02-2.58; P = 0.03, and OR = 0.61; 95% CI = 0.38-0.97; P = 0.03, respectively). IL-1RAcP -8261 T allele and -8183 A allele were also associated with persistent HBV infection (OR = 0.64; 95% CI = 0.46-0.89; P = 0.008 in the co-dominant model and OR = 0.20; 95% CI = 0.05-0.71; P = 0.01 in the recessive model, respectively).
IL-1RAcP -8261 T allele and -8183 A allele comprised the haplotype IL-1RAcP B1ht1, and
this haplotype also showed a relation with a persistent HBV infection (OR = 0.64; 95% CI = 0.46-0.89; P <0.01 in the co-dominant model). On the other hand, the IL-1RAcP -8261C/-8183G haplotype was associated with HBV clearance (OR = 1.92; 95% CI = 1.40-17.28; P = 0.01 in the dominant model). The IL-1B -2023G>C SNP is located in the promoter region, and the IL-1RAcP -8261T>C SNP and -8183A>G SNP are located in the intron region. There is no known amino acid change in these SNPs. Further research is required to understand the functions of the variants that promote persistent HBV infection.
IL-1B has been suggested to be a potential candidate gene for cancer development. In
Caucasians infected with Helicobacter pylori, the IL-1B -31C/-511T haplotype has been associated with an increased risk of gastric cancer (ElOmar, et al., 2000; 2001). The IL1B -31 T/T genotype and the IL-1B --31T/-511C haplotype have been found to be associated with the presence of HCC in Japanese patients with chronic HCV infection (Wang, et al., 2003). It has also been reported that the IL-1B -511C, -31T allele was a genetic marker for the
- 20 -
development of HCC in chronic hepatitis B patients in a Thai population (Hirankarn, et al., 2006). This discordance may be due to the differences of organs and the heterogeneous nature of carcinogenesis in these organs. Our study showed only a marginal relation between the IL-1B 289 C allele and an increased risk of HCC in the co-dominant model (OR=1.55; 95% CI=1.00-2.40; P =0.04). Large-scale case-control or cohort studies are needed in the future to confirm the association of the IL-1B and IL-1RAcP SNPs with the development of HCC.
- 21 -
V. CONCLUSION
In conclusion, the present study suggests that SNPs within IL-1B and IL-1RAcP genes are associated with persistent HBV infection. This study is the first to show that several
IL-1RAcP polymorphisms may be significant genetic markers for a persistent HBV infection.
Our novel findings provide further evidence that genetic factors are important for determining the outcome of HBV infection.
- 22 -
REFERENCES
1. Arend WP, Palmer G, Gabay C: IL-1, IL-18, and IL-33 families of cytokines. Immunol
Rev 223: 20-38, 2008
2. Bortolami M, Kotsafti A, Cardin R, Farinati F: Fas / FasL system, IL-1beta expression and apoptosis in chronic HBV and HCV liver disease. J Viral Hepat 15: 515-522, 2008
3. Cheong JY, Cho SW, Hwang IL, Yoon SK, Lee JH, Park CS, Lee JE, Hahm KB, Kim JH: Association between chronic hepatitis B virus infection and interleukin-10, tumor necrosis factor-alpha gene promoter polymorphisms. J Gastroenterol Hepatol 21: 1163-1169, 2006
4. Dinarello CA: Biologic basis for interleukin-1 in disease. Blood 87: 2095-2147, 1996
5. El-Omar EM, Carrington M, Chow WH, McColl KE, Bream JH, Young HA, Herrera J, Lissowska J, Yuan CC, Rothman N, Lanyon G, Martin M, Fraumeni JF, Jr., Rabkin CS: Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature 404: 398-402, 2000
6. El-Omar EM, Carrington M, Chow WH, McColl KE, Bream JH, Young HA, Herrera J, Lissowska J, Yuan CC, Rothman N, Lanyon G, Martin M, Fraumeni JF, Jr., Rabkin CS: The role of interleukin-1 polymorphisms in the pathogenesis of gastric cancer. Nature
- 23 - 412: 99, 2001
7. Hirankarn N, Kimkong I, Kummee P, Tangkijvanich P, Poovorawan Y: Interleukin-1beta gene polymorphism associated with hepatocellular carcinoma in hepatitis B virus infection. World J Gastroenterol 12: 776-779, 2006
8. Hohler T, Kruger A, Gerken G, Schneider PM, Meyer zum Buschenefelde KH, Rittner C: A tumor necrosis factor-alpha (TNF-alpha) promoter polymorphism is associated with chronic hepatitis B infection. Clin Exp Immunol 111: 579-582, 1998
9. Kühn K, Hashimoto S, Lotz M: IL-1 beta protects human chondrocytes from CD95-induced apoptosis. J Immunol. 164: 2233-2239, 2000
10. Kim YJ, Lee HS, Im JP, Min BH, Kim HD, Jeong JB, Yoon JH, Kim CY, Kim MS, Kim JY, Jung JH, Kim LH, Park BL, Shin HD: Association of transforming growth factor-beta1 gene polymorphisms with a hepatocellular carcinoma risk in patients with chronic hepatitis B virus infection. Exp Mol Med 35: 196-202, 2003
11. Kim YJ, Yoon JH, Kim CY, Kim LH, Park BL, Shin HD, Lee HS: IGF2 polymorphisms are associated with hepatitis B virus clearance and hepatocellular carcinoma. Biochem
Biophys Res Commun 346: 38-44, 2006
- 24 -
Interleukin-1 protects transformed keratinocytes from tumor necrosis factor-related apoptosis-inducing ligand. J Biol Chem. 273: 29247-29253, 1998
13. Lee WM: Hepatitis B virus infection. N Engl J Med 337: 1733-1745, 1997
14. Lee ZH, Lee SE, Kim CW, Lee SH, Kim SW, Kwack K, Walsh K, Kim HH: IL-1alpha stimulation of osteoclast survival through the PI 3-kinase/Akt and ERK pathways. J
Biochem 131: 161-166, 2002
15. Migita K, Maeda Y, Abiru S, Nakamura M, Komori A, Miyazoe S, Nakao K, Yatsuhashi H, Eguchi K, Ishibashi H: Polymorphisms of interleukin-1beta in Japanese patients with hepatitis B virus infection. J Hepatol 46: 381-386, 2007
16. Rahman MA, Dhar DK, Yamaguchi E, Maruyama S, Sato T, Hayashi H, Ono T, Yamanoi A, Kohno H, Nagasue N: Coexpression of inducible nitric oxide synthase and COX-2 in hepatocellular carcinoma and surrounding liver: possible involvement of COX-2 in the angiogenesis of hepatitis C virus-positive cases. Clin Cancer Res 7: 1325-1332, 2001
17. Schmidt M, Pauels HG, Lugering N, Lugering A, Domschke W, Kucharzik T: Glucocorticoids induce apoptosis in human monocytes: potential role of IL-1 beta. J
- 25 -
18. Shin HD, Park BL, Cheong HS, Yoon JH, Kim YJ, Lee HS: SPP1 polymorphisms associated with HBV clearance and HCC occurrence. Int J Epidemiol 36: 1001-1008, 2007
19. Tassopoulos NC, Papaevangelou GJ, Sjogren MH, Roumeliotou-Karayannis A, Gerin JL, Purcell RH: Natural history of acute hepatitis B surface antigen-positive hepatitis in Greek adults. Gastroenterology 92: 1844-1850, 1987
20. Tian Z, Shen X, Feng H, Gao B: IL-1 beta attenuates IFN-alpha beta-induced antiviral activity and STAT1 activation in the liver: involvement of proteasome-dependent pathway. J Immunol 165: 3959-3965, 2000
21. Wang Y, Kato N, Hoshida Y, Yoshida H, Taniguchi H, Goto T, Moriyama M, Otsuka M, Shiina S, Shiratori Y, Ito Y, Omata M: Interleukin-1beta gene polymorphisms associated with hepatocellular carcinoma in hepatitis C virus infection. Hepatology 37: 65-71, 2003
22. Watson RW, Rotstein OD, Parodo J, Bitar R, Marshall JC: The IL-1 beta-converting enzyme (caspase-1) inhibits apoptosis of inflammatory neutrophils through activation of IL-1 beta. J Immunol. 161: 957-962, 1998
23. Wesche H, Korherr C, Kracht M, Falk W, Resch K, Martin MU: The interleukin-1 receptor accessory protein (IL-1RAcP) is essential for IL-1-induced activation of
- 26 -
interleukin-1 receptor-associated kinase (IRAK) and stress-activated protein kinases (SAP kinases). J Biol Chem 272: 7727-7731, 1997
24. Yang CY, Kuo TH, Ting LP: Human hepatitis B viral e antigen interacts with cellular interleukin-1 receptor accessory protein and triggers interleukin-1 response. J Biol
Chem 281: 34525-34536, 2006
25. Zhang PA, Li Y, Xu P, Wu JM: Polymorphisms of interleukin-1B and interleukin-1 receptor antagonist genes in patients with chronic hepatitis B. World J Gastroenterol 10: 1826-1829, 2004
- 27 - - 국문요약 -
Interleukin-1 beta 및 Interleukin-1 Receptor Accessory
Protein 유전자 다형성과 만성 B 형간염 및 간세포암종
발생과의 관련성
아주대학교 대학원의학과 김 순 선 (지도교수 : 조 성 원)목적: B 형 간염 바이러스(HBV)는 감염 후 만성 간염, 간경변증 및 간세포암종 등 다양한 임상 경과를 보이며 이에 영향을 미치는 숙주의 면역 반응 요인으로 사이토카인의 역할은 중요하다. Interleukin (IL)-1 은 자연면역에서의 숙주 반응의 매개자로 대표적인 염증성 사이토카인으로 알려져 있다. 본 연구는 최근 주목받는 single nucleotide polymorphism (SNP) chip 을 이용한 high-throughput 기술을 적용하여 B 형 간염의 만성화 및 간세포암종 발생과 관련된 유전자 다형성을 알아보고자 하였다. 방법: 2000 년 6 월부터 2006 년 2 월까지 아주대학교병원에 내원한 399 명을 대상으로 선정된 1,536 개의 SNP 에 대해 Illumina 사의 chip 으로 주문 제작하였다. 대상 환자들은 바이러스 제거군 (n=107, HBsAg 음성, Anti-HBc 및 Anti-HBs 양성), 만성 간염군 (n=111), 간경변증군 (n=95), 간세포암종군 (n=86)으로 분류하였다.
결과: IL-1β 유전자의 promoter 2 부위 (-581T>C, -2023G>C)와 intron 2 부위 (3340A>G, 289T>C) 및 1L-1 receptor accessory protein (RAcP) 유전자의 promoter 1 부위 (51668T>A)와 intron 5 부위 (8261T>C,
-- 28 --
8183A>G, -256C>T, 59264G>A, 65445A>G)가 다형성을 보였다. 만성 간염군, 간경변증군 및 간세포암종군에서 IL-1B-2023 C allele, IL-1RAcP -8261 T allele 및 -8183 A allele 이 바이러스 제거군에 비해 유의하게 많았다. (OR, 1.63, P = 0.03, OR, 0.64, P <0.01 and OR, 0.20, P = 0.01, respectively). IL-1B 289C allele 이 HCC group 에서 만성 간염군 및 간경변증군에 비해 많았다(OR, 1.55, P = 0.04). Haplotype 분석에서 IL-1B -2023C/-581C/2893C haplotype 과 IL-1RAcP -8261T/-8183A haplotype 이 만성 간염군, 간경변증군 및 간세포암종군에서 바이러스 제거군에 비해 유의하게 많았으며 간세포암종과 연관된 haplotype 은 없었다.
결론: SNP chip 을 이용한 분석에서 IL-1B promoter -2023 부위 및 IL-1 RAcP intron -8261, -8183 부위의 유전자 다형성은 HBV 만성화와 연관된 숙주 인자이다.
핵심어: 만성 B 형간염, 간세포암종, Interleukin-1β, Interleukin-1 receptor accessory protein, Single nucleotide polymorphism