α-amino-5-methyl-4-isoxazole
propionate(AMPA)
α-amino-5-methyl-4-isoxazole
propionate(AMPA)
i 국문요약 –
백서 피질 신경원에서
α
amino 5 methyl 4 isoxazole propionate
로 유도된 뇌독성에 대한
의 효과
(AMPA)
Propofol
세포손상 주요 기전중 하나로 뇌허혈시 세포외 glutamate의 급격한 증가는 N methyl D aspartate (NMDA) 혹은 α amino 5 methyl 4 isoxazole propionate (AMPA),
수용체 등을 과도하게 흥분시켜 를 활성화시킨다
kainate excitotoxic cascade .
그러나, AMPA 수용체 자극시 어떤 형태의 세포손상이 오는지(necrosis vs
자극에 대해 의 뇌보호 작용이 있는지 기전이
apoptosis), AMPA propofol ,
무엇인지 등은 여전히 불명확하다. 저자들은 백서 (Sprague-Dawley) 피질 신경원 배양 모델에서 AMPA 자극을 주며 어떤 형태의 신경세포 손상을 일으키는지, propofol이 AMPA 자극에 대해 뇌보호 효과가 있는지, propofol이 세포내로의 Ca++ influx에 어떤 영향을 미치는지 알아보고자 하였다. 백서를 이용하여 피질신경원 혼합배양을 하였으며 세포배양 시작, 13일째 되는 날 실험을 실시하였다. Propofol이 AMPA자극에 대해 뇌보호 효과가 있는지 보기 위해 세포들은 아무약도 처리하지 않은 대조군, NMDA 100 µM 처리한 Full kill 군, AMPA 50 µM 처리군, AMPA 50 µM + propofol 0.1, 1, 25, 50 µM 처리군,
단독처리군 등으로 나뉘었다 시간 배양후 염색을
propofol 50 µM . 72 trypan blue
이용하여 살아있는 세포수를 세었으며 이를 대조군값, 0, Full kill군 값 100으로 놓고 각 군에서의 세포사율을 구하였다. Apoptosis 발생 여부 및 apoptosis에
투여가 미치는 영향을 보기위해 대조군 처리군
50 µM + propofol 50 µM 처리군으로 나누었으며, TUNEL assay, NeuN stain,
등의 을 하여서 발생비율을 구하였다 자
Hoechst stain Triple stain apoptosis . AMPA 극시의 세포내로의 calcium influx에 propofol 투여가 미치는 영향을 보기위하여 대조군, AMPA 50 µM 처리군, AMPA 50 µM + propofol 50 µM 처리군으로 나누 었으며 confocal 현미경을 이용한 calcium imaging을 실시하였다 통계는. SPSS
을 이용하여 실시하였고 사후 검증은 로
12.0 One way ANOVA test Bonferroni test
실시하였으며, P < 0.05시 의의있는 것으로 간주하였다.
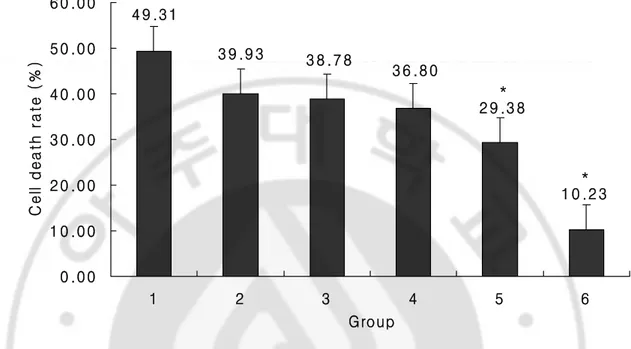
세포사율은 AMPA 50 μM 단독투여군에서 49.31%, AMPA 50 µM + propofol 50 µM 혼합투여군에서 29.38%로 의의있게 감소하였다(n=45/ )(Fig. 1, p<0.05).군
결과 대조군에 비해 투여군에서 일으킨 신
Triple staining AMPA 50 Mμ apoptosis
경원의 수가 의의있게 증가하였다(12.33 vs 28.01 %)(P<0.05). 또한, AMPA 50 Mμ
혼합투여군에서 일으킨 신경원의 수가 의의있게 감소
+ propofol 50 Mμ apoptosis
하였다(28.01 vs 20.07 %)(P<0.05). Calcium imaging결과 AMPA 50 Mμ 단독투여군 에 비해 AMPA 50 μM + propofol 50 μM 혼합투여군에서 intraneural calcium
의 최대치는 의의있는 감소를 보였다
influx (3164.98 nA vs 2276.98 nA)(Fig. 6,7, P<0.05).
결론적으로 백서 피질 신경원 혼합배양 AMPA 자극시 necrosis와 apoptosis가 혼 합된 형태로 추측되는 세포손상을 보였으며, 50 μM propofol의 처치는 뇌세포보 호 효과가 있었고 이는 부분적으로 세포내로의, calcium의 이동의 감소에 기인하 는 것으로 사료된다.
-Fig. 1. The effects of Propofol on Cell Death Rate on 72 hr, AMPA 50 M stimulation
μ ... 11
Fig. 2. Triple stain in Control group ... 12
Fig. 3. Triple stain in 50 μM AMPA Group ... 13
Fig. 4. Triple stain in AMPA 50 μM + propofol 50 μM treated group
... 14
Fig. 5. The effects of propofol 50 μM on 72 hr AMPA 50 μM stimulation induced apoptosis ... 15
Fig. 6. The changes of Ca2+ fluorescence ... 16
I. 서 론
뇌허혈시 세포외 glutamate 와 같은 흥분성 아미노산의 급격한 증가와 N‐methyl‐D‐aspartate (NMDA) 혹은 α-amino-5-methyl-4-isoxazole propionate
(AMPA) 수용체와 같은 glutamate 수용체의 과도한 흥분에 의한 신경흥분독성 연속단계 (excitotoxic cascade)의 활성화가 세포 손상의 주요 기전 중 하나로 알려져있다(Rothman 등, 1986; Koh 등, 1987; Goldberg 등, 1987; Koh 등, 1990; Choi, 1992).
2,3 diisopropylphenol (propofol)은 주로 GABAA 수용체에 작용하여 마취효과를
나타내는 정맥마취제이며, 매우 빠른 각성 등의 장점으로 인해 외래 통원수술에
많이 사용되고 있고, 여러가지 기전에 의해 뇌보호 작용이 있다고 알려지면서
(Kochs 등, 1992; Han 등, 1994; Orser 등, 1995; Yamakura 등, 1995; Peters 등, 2001;
Ligamaneni 등, 2001; Peters 등, 2001; Velly 등, 2003) 신경외과 마취영역 등에서
점차 사용이 증가되고 있다.
AMPA 수용체의 과도한 자극은 괴사(necrosis) 혹은 세포자멸사(apoptosis)
(Garthwaite 등, 1991; Larm 등, 1997; Ligamaneni 등, 2001) AMPA 수용체에 대한
propofol 의 작용과 AMPA 자극에 대한 propofol 의 뇌보호 작용에 관한 보고는
거의 없으며, 보고 내용도 일정치 않다(Yamakura 등, 1995; Zhu 등, 1997; Krampfl 등, 2005). 저자들은 백서 피질 신경원 배양 모델에서 72 시간 동안 AMPA 50 μM 자극과 동시에 0.1‐50 μM 농도의 propofol 을 투여하여 propofol 이 신경세포 손상에 어떤 효과가 있는지, 72 시간 AMPA 50 μM 자극시 apoptosis 형태의 신경세포 손상을 얼마나 일으키며, propofol 을 주면 어느 정도 감소 효과가 있는지, propofol 이
AMPA 자극에 의한 intracellular calcium influx 의 감소에 얼마나 영향을 미치는지
II. 연구대상 및 방법
이 실험은 본 병원 실험실 동물 관리 및 윤리 규정에 따라 행하였다. 몸무게 250-300 g 의 임신 16 일 되는 백서 (Sprague-Dawley)를 물과 먹이가 충분히 공급되는 사육실에서 주위 환경에 적응시키며 하루 동안 사육한 후 임신 17 일째 세포배양 실험을 진행하였다. A. 세포배양실험 전 20 mM glucose, 5% horse serum(HS), 5% fetal bovine serum(FBS), 2 mM glutamine 을 Eagle’s minimum essential medium (MEM)과 혼합하여 만든 100 µg/ml 의 poly D lysine 과 4 µg/ml 의 laminin 으로 24-well 배양 플레이트와 35 mm 배양 접시를 코팅 처리하였다. 이때 일부는 immuno-staining 을 위해서 24-well 배양 플레이트에 세포부착용 플라스틱인 아크라를 코팅처리하였다. 백서를 ether 마취상자에 넣어 마취제를 흡입시켰으며, 호흡이 없어진 것을 확인한 후 꺼내어 경추탈골을 시켰다. 태아를 제왕절개를 통해 꺼낸 후, 20 mM glucose 를 섞은 MEM 용액에 담그고 태아의 뇌피질 부분을 분리하였다. 분리된 뇌 피질을
pasteur pipette 으로 가볍게 분쇄한 뒤 미리 코팅된 24-well 배양 플레이트에 400 ㎕, 35 mm 배양 접시에 2 ml 씩 분주하고, 37℃, 5% CO2 배양기에서 배양하였다.
배양한 세포는 1 주후 glia 가 자라나면 20 mM glucose, 5% HS, 2 mM glutamine 을 섞은 MEM 용액과 혼합하여 만든 10 μM Ara C 로 처리하였고, Ara C 처리 후부터 실험 전까지 이틀 간격으로 Ara C 를 제외한 같은 배양액으로 1/3 씩 바꾸어 주었다. B. 살아있는 세포수 측정 1. 약물처리 세포배양 시작 후 13 일째 되는 날 실시하였으며, 72 시간 동안 다음과 같은 군으로 나누어 약물을 처리하였다. 즉, 약물 처리하지 않은 대조군 (대조군), 100 µM NMDA 처리 대조군 (Full kill 군), 50 µM AMPA 처리군, 50 µM AMPA + propofol 0.1, 1, 25, 50 µM 처리군, propofol 50 µM 단독처리군으로 나누었다.
약물처리 72 시간 후 10% trypan blue stain 을 15 ㎕/well 씩 첨가하여 37℃, 5% CO2 배양기에서 15 분간 배양하여 염색하였다. 이후 eyepiece micrometer square
10×10 grid (Olympus, Japan)를 사용하여 염색되지 않은 살아있는 세포수를 세었다. 모든 군은 5 마리의 쥐에서 결과를 얻었으며, 각 well 당 세 구역씩 선정하였고 군당 45 구역을 선택하여 well 의 중심 부위로부터 겹치지 않게 살아있는 세포수를 세었다(n=45). 세포수를 수식 [100×(대조군값 실험군값/대조군값‐Full‐kill 군값)]을 사용하여 대조군의 값을 0 으로, Full kill 군의 값을 100 으로 하고, 이에 대한 각 실험군의 죽은 세포 백분율을 구하였다. C. Immuno-staining 아크라 위에서 키운 13 일째 세포를 아무 약도 처리하지 않은 대조군, 50 µM AMPA 처리군, 50 µM A M P A + propofol 50 µM 처리군의 세포를 72 시간 후 phosphated-buffered saline (PBS)으로 3 회 씻어낸 후 4% paraformaldehyde (Sigma, USA)로 상온에서 1 시간 동안 세포를 고정시킨다. PBS 로 3 회 씻은 후 1% Triton X 100 (USB, USA)이 포함된 PBS 로 4℃에서 2 분 동안 둔다.
Neuronal nuclei (NeuN) Stain 은 neuronal cell 의 존재를 알기 위한 형광염색이며 (초록색), 1 차 항체인 mouse anti‐neuronaleuclein(NeuN) monoclonal antibody (Jackson ImmunoReserch Laboratories, Inc., West Grove, PA, USA)를 1:250 비율로 20µl 첨가하여 2 시간 동안 incubation 시켰다. 이후 PBST 로 3 회 씻어내었으며, 2 차 항체인 Fluorescein (FITC)‐conjugated affinipure Goat Anti‐Mouse IgG (Jackson ImmunoReserch Laboratories, Inc., West Grove, PA, USA)를 1:200 비율로 20µl 첨가 후2 시간 동안 반응시켰다.
Hoechst Stain 은 DNA 나 핵이 있는지 알기 위한 형광염색이며 (파랑색), PBS 로 2 회 씻어낸 후 Hoechst 33258 (Molecular Probes, Eugene, OR, USA)을 1:1000 비율로 5 분 동안 염색한 후 PBS 로 2 회, 증류수로 1 회 씻어내었다.
Terminal deoxynucleotidyl transferase (TdT)‐mediated dUTP‐biotin nick‐end labeling (TUNEL) stain 은 apoptotic cell 을 알아보기 위한 형광염색이며 (빨간색), In situ cell death detection kit (TMR red, Roche, Mannheim, Germany) 을 사용하였다. Mounting solution 을 사용하여 slide glass 에 붙인 후, confocal 현미경 (Carl Zeiss Laser Scanning System LSM 510, Oberkochen, Germany)을 이용하여 ×400 과 ×800 배율로 관찰하였다. 각 군에서 ×400 배율 하에서 5 구역을 선택하여, NeuN 과
Hoechst stain 으로 염색된 전체 신경원의 수를 센 후 TUNEL assay 에서 형광 염색된 세포 중 NeuN 과 Hoechst stain 이 동시에 되었는지 확인하여 apoptosis 가 일어난 세포수를 세었으며, 전체 신경원수 중 apoptosis 가 일어난 세포의 비율을 구하였다.
D. Calcium imaging
35 mm 배양접시에 키운 13 일째 세포에 AMPA 및 propofol 처리하여 propofol 이 AMPA 자극에 의한 세포 내로의 Ca++ 유입에 미치는 영향을 살펴보았는데, 약물
처리 전 세포를 HCSS buffer (NaCl 70.1 g, KCl 4 g, MgCl2 1.6 g, CaCl2 2.6 g, glucose 27
g, HEPES 47.7 g, NaOH 4 g) 2 ml 로 3 번 씻어낸 후, fluo‐3 AM (molecular probesTM,
USA) dye 를 5 μM 이 되도록 HCSS buffer 와 섞어 1.5 ml/dish 씩 넣어주고, 5% CO2
배양기에서 30 분간 배양하였다. 이후 세포를 HCSS buffer 로 3 번 씻어낸 후 HCSS buffer 1ml 씪 넣어준 후 400 배의 Argon laser 렌즈가 부착된 confocal 현미경(Carl Ziess, Germany)에서 live cell image 로 0 초에서 400 초까지의 시간 경과에 따른 세포 내의 칼슘이온의 변화를 관찰하였는데, 대조군은 HCSS buffer 1 ml, propofol 50 µM , AMPA 50 µM , AMPA 50 µM + propofol 50 µM 처리군은
HCSS buffer 1 ml 에 각 농도의 약제를 섞은 후, 각 용액을 50 초에 주입하여 400 초까지 세포의 변화를 촬영하였다. 촬영된 각 군의 image 내의 세포 중 유의한 밝기를 보이는 12 개 세포를 선택하여 12 개의 세포의 변화 그래프와 수치 값을 얻었다. 그래프와 값은 Carl Zeiss laser scanning system LSM 510 (Carl
Ziess, Germany) 프로그램에서 산출되었으며, 프로그램에서 산출된 수치 값을
엑셀 (Microsoftware Office 2000 EXCEL, USA )에서 각 군별로 평균값을 낸 후, 각 군의 평균값을 모아 그래프를 얻었다. 또한, 각 군에서 Ca2+ influx 값 최대치를
측정하였다.
통계는 SPSS 12.0 을 이용하여 Oneway ANOVA test 실시하였고 사후검증은 Bonferroni test 로 실시하였으며, p < 0.05 시 의의있는 것으로 간주하였다.
실험에 필요한 시료들 중 Eagle’s minimum essential medium (MEM), Glucose, 5% horse serum(HS), 5% fetal bovine serum(FBS), trypan blue stain 은 Gibco BRL (Carlsbad, CA, USA)로 부터 Glutamine, poly D lysine, cytosine, β D arabino furanoside hydrochloride (Ara C)은 Sigma-Aldrich (St. Louis, MO, USA)로 부터 laminin 은 Invitogen (Carlsbad, CA, USA)으로 부터 구입하였다.
III. 결 과
세포사된 신경원의 비율은 AMPA 50 μM 단독투여군 (49.31%)에 비해, AMPA 50
μM + propofol 50 μM 혼합투여군에서 29.38%로 의의있게 감소하였다 (P<0.05) (Fig.
1).
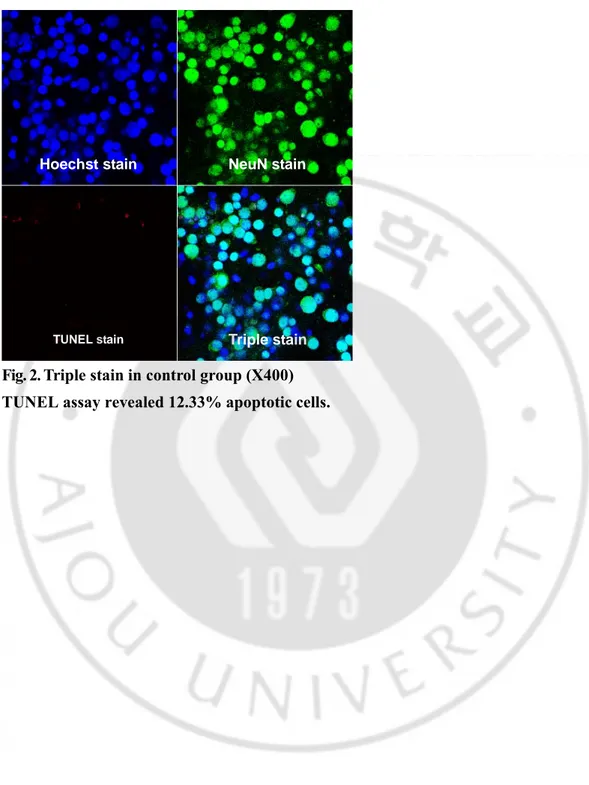
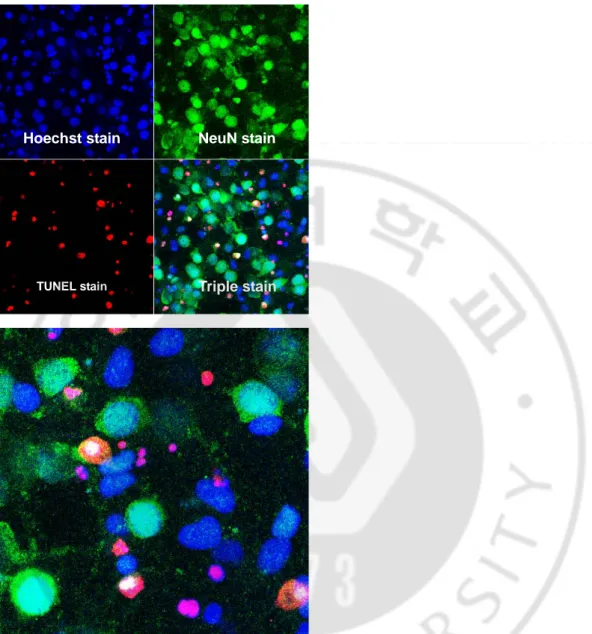
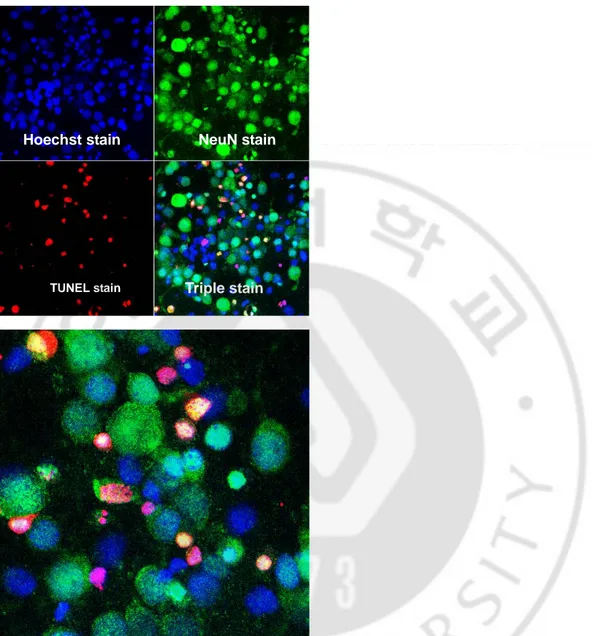
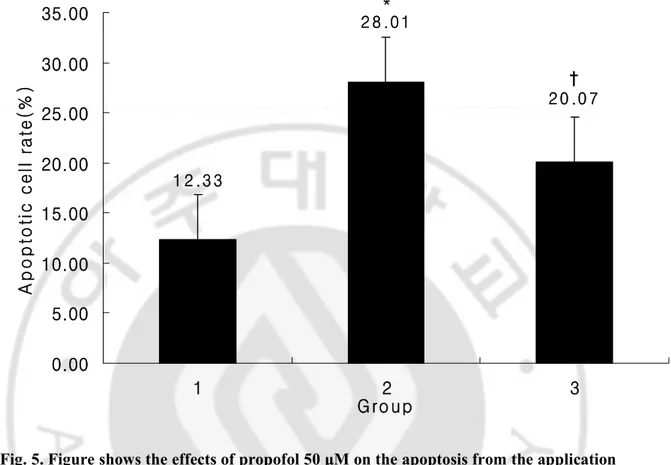
Triple staining 결과 apoptosis 일으킨 신경원의 비율은 대조군에서 12.33%,
AMPA 50 μM 단독투여군에서 28.01%, AMPA 50 μM + propofol 50 μM
혼합투여군에서 20.07% 였으며, 대조군에 비해 AMPA 50 μM 단독투여군에서
의의있게 증가하였으며 (P<0.05), AMPA 50 μM 단독투여군에 비해 AMPA 50 μM +
propofol 50 μM 혼합투여군에서 의의있게 감소되었다 (P<0.05) (Fig. 2, 3, 4, 5).
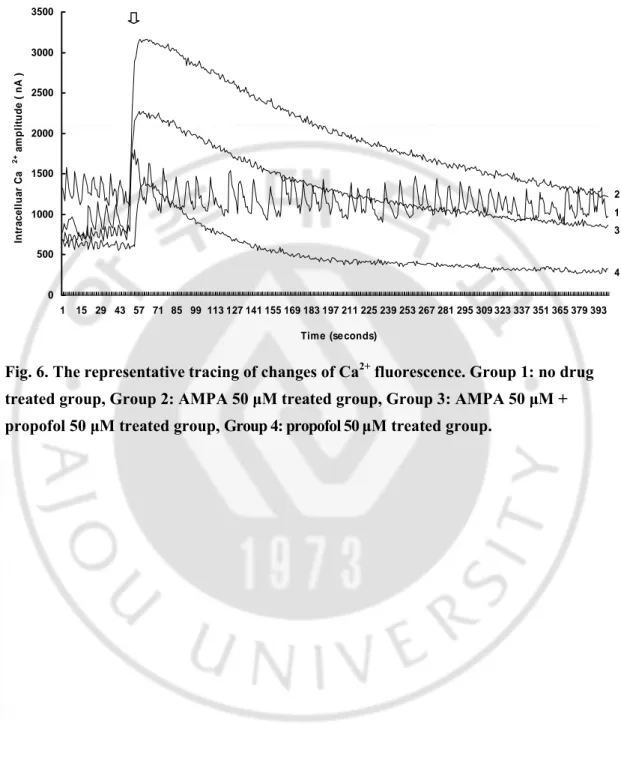
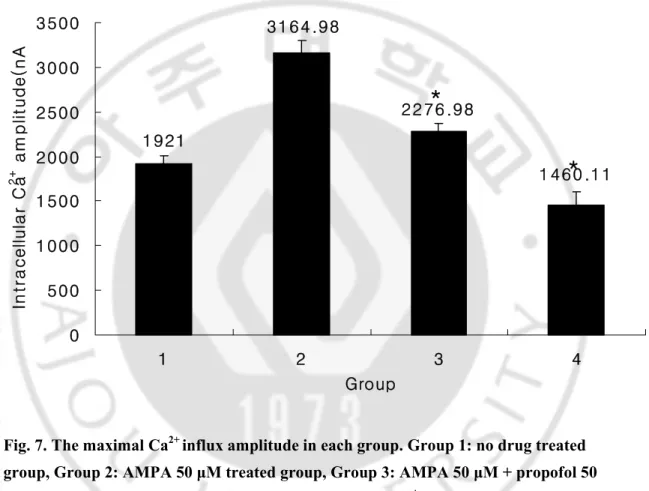
Calcium imaging 결과 AMPA 50 μM 단독투여군에 비해 AMPA 50 + propofol 50 μM 혼합투여군에서 intraneural calcium influx 의 최대치의 의의있는 감소를 보였으며 (3164.98 nA vs 2276.98 nA, P<0.05)(Fig. 6, 7), 각 군에서 약물 처치 전 최저치 calcium influx 와 약물 처치 후 최고치 calcium influx 를 구한 후 calcium influx 증가치를 백분율로 나타내면, 대조군은 164.67%, AMPA 50 μM
단독투여군은 450.26%, AMPA 50 μM + propofol 50 μM 혼합투여군은 351.26%, propofol 50 μM 단독투여군은 222.75% 였다.
* 1 0 .2 3 * 2 9 .3 8 3 6 .8 0 3 9 .9 3 4 9 .3 1 3 8 .7 8 0 .0 0 1 0 .0 0 2 0 .0 0 3 0 .0 0 4 0 .0 0 5 0 .0 0 6 0 .0 0 1 2 3 4 5 6 Group C e ll d e a th r a te ( % )
Fig. 1. Figure shows the cell death rate in each group. Relative cell death rate was computed with the set as 0 in control group and 100 in Full-kill group applied with AMPA 50 μM for 72 hr. Adding propofol 50 μM to AMPA decreased cell death rate. Number of each group is 45. Values are mean ± SD. Group 1: no drug treated group, Group 2, 3, 4, 5: AMPA 50 μM + Propofol 0.1, 1, 25, 50 μM treated group, Group 6: Propofol 50 μM treated group. *: P<0.05 vs, AMPA 50 μM treated group.
Fig. 2. Triple stain in control group (X400) TUNEL assay revealed 12.33% apoptotic cells.
NeuN stain Hoechst stain
Triple stain
Fig. 3. Triple stain in 50 μM AMPA treated group. left (X400), right (X 800). TUNEL assay revealed 28.01% apoptotic cells.
NeuN stain
TUNEL stain Triple stain Triple stain
Fig. 4. Triple stain in AMPA 50 μM + propofol 50 μM treated group, left (X400), right (X 800). TUNEL assay revealed 20.07% apoptotic cells.
Hoechst stain
Triple stain Triple stain
NeuN stain
럏. 5.
Fig. 5. Figure shows the effects of propofol 50 μM on the apoptosis from the application with AMPA 50 μM for 72 hr. Number of neurons/field were counted by Neuronal nuclei and Hoechst stain and apoptotic cells were counted by TUNEL assay. Then, apoptotic cell rate were calculated. Cell number of each group is 15. Values are mean ± SD. Group 1: no drug treated group, Group 2: AMPA 50 μM treated group, Group 3: AMPA 50 μM + propofol 50 μM treated group. *: P<0.05 vs. group 1, †:P<0.05 vs. group 2. 1 2 .3 3 * 2 8 .0 1 † 2 0 .0 7 0.00 5.00 10.00 15.00 20.00 25.00 30.00 35.00 1 2 3 G ro up A p o p to ti c c e ll r a te (% )
0 500 1000 1500 2000 2500 3000 3500 1 15 29 43 57 71 85 99 113 127 141 155 169 183 197 211 225 239 253 267 281 295 309 323 337 351 365 379 393 Time (se conds)
In tr acel lu ar Ca 2+ a m pli tud e ( n A ) 2 1 3 4
Fig. 6. The representative tracing of changes of Ca2+ fluorescence. Group 1: no drug treated group, Group 2: AMPA 50 μM treated group, Group 3: AMPA 50 μM + propofol 50 μM treated group, Group 4: propofol 50 μM treated group.
Fig. 7. The maximal Ca2+ influx amplitude in each group. Group 1: no drug treated group, Group 2: AMPA 50 μM treated group, Group 3: AMPA 50 μM + propofol 50 μM treated group, Group 4 : propofol 50 μM treated group. *: P < 0.05 vs. Group 2.
1921 3164.98 2276.98 1460.11 0 500 1000 1500 2000 2500 3000 3500 1 2 3 4 Group In tr a c el lu la r C a 2+ a mp li tu d e (n A
*
*
IV. 고 찰
뇌허혈이나 뇌손상 같은 임상상황에서 glutamate 와 같은 흥분성 아미노산들이 synaptic cleft 에 급격히 증가하게 되며 이들에 의한 excitotoxicity 가 뇌손상의 주요 기전 중 하나이다(Benveniste 등, 1984; Rothman 등, 1986; Goldberg 등, 1987; Koh 등, 1987; Koh 등, 1990; Choi 등, 1990; Choi, 1992; Nishizawa 등, 2001).
증가된 glutamate 는 시냅스 후부의 N‐methyl‐D‐aspartate (NMDA), α-amino-5-methyl-4-isoxazole propionate (AMPA), kainate 등의 ionotropic 수용체를 흥분시키게 되며 (Watkins 등, 1987; Dingledine 등, 1999), 이에 의해 급격히 증가된 세포내 Ca2+에
의해 일련의 뇌손상 작용이 일어나게 되는데 (Rothman 등, 1986; Goldberg 등, 1987; Koh 등, 1990; Choi 등, 1990; Choi, 1992), 첫째로 Ca2+ 과 함께 Na+, Cl 등이
세포내로 같이 유입되어 세포의 삼투성 팽창을 가져오며 (Rothman 등, 1985) 둘째로 Ca2+ 이 공통의 2 차 매개체로 작용하기 때문에 여러가지 해로운 효소들을
활성화시킬 수 있는데, neuronal nitric oxide synthase (nNOS)에 의한 NO 의 생성, reactive oxygen species (ROS)의 생성, arachidonic acid 생성 등이 이에 속한다(Lazarewicz 등, 1990; Gunasekar 등, 1995; Li 등, 2002). 이와 같은 일련의 손상 과정은 Na+ 과부하에 의한 세포질성 구조물들의 팽창을 주 특징으로 하는
괴사 (necrosis) 형태의 세포사멸 (cell death)를 주로 일으키지만 (Choi 등, 1987; Gwag 등, 1997), 최근에는 DNA 분열과 염색질 응축을 특징으로 하는 apoptosis 형태의 세포사멸도 glutamate 수용체에 의한 excitotoxicity 에서 나타난다는 보고가 많다(Bonfoco 등, 1995; Ankarcrona 등, 1995).
본 연구에서 사용된 AMPA 자극에 의해 활성화되는 AMPA 수용체는 4 개의
단백질인 GluR1-4 (A-D)의 조합에 의해 이루어져 있으며 (Madden 등, 2002),
이들의 조합이나 RNA editing 에 의해 AMPA 수용체의 기능이 달라지는데
(Hollmann 등, 1991; Seeburg 등, 1996), GluR2 가 많이 함유되어 있으면 Ca2+
투과성이 낮고, GluR2 가 적게 함유되어 있으면 Ca2+ 투과성이 높다고 하며,
대부분의 신경세포는 전자에 속하며, 해마에 분포하는 neuron, neocortex 등이
후자에 속한다(lino 등, 1990; Hollmann 등, 1991; Burnasheve 등, 1992; Jonas 등, 1994;
Seeburg 등, 1996; Madden 등, 2002). 그러나, 전뇌허혈과 같은 임상 상황에서는
GluR2 의 표현이 억제되는 새로운 형태의 AMPA 수용체가 형성되며 Ca2+
허혈 등에 의해 지연되어 나타나는 뇌병변 (delayed neurodegeneration)의 주요
원인이고, 주로 apoptosis 형태의 병변을 일으켰다 한다(Pellegrini-Giampietro 등,
1992; Pellegrini-Giampietro 등, 1997). 본 연구에서는 대뇌 피질 신경원을 사용하여
연구하였으며, Ca2+ 투과성이 높다고 알려진 세포는 아니었지만, fluo-3 AM 를
이용한 calcium imaging 실험에서 AMPA 50 μM 자극 시 450.26%의 신경세포
내로의 calcium influx 의 증가율을 보여, 세포내 calcium 증가에 의한 세포 손상의
가능성을 보여주었으며, propofol 50 μM 혼합투여 시 351.26%로 calcium
influx 증가율이 감소하여, 본 연구에서 72 시간 동안, 50 μM AMPA 자극 시
세포사율이 49.31%에서 AMPA 50 μM + propofol 50 μM 혼합투여 시 세포사율이
29.38%로 의의있게 감소한 원인 중 하나로 제시될 수 있겠다.
Glutamate 수용체의 과도한 흥분에 의해 일어나는 excitotoxicity 는 대부분
NMDA 수용체의 의해 일어나지만, AMPA 수용체 등의 비 NMDA 수용체도
Frandsen 등은 AMPA 등의 비-NMDA 수용체도 excitotoxicity 에 관여한다
보고하였다(Frandsen 등, 1989). Garthwaite 등은 AMPA 에 의한 세포손상이 세가지
기전으로 일어남을 관찰하였는데, 소뇌의 golgi 세포에서 빠르게 팽창성으로 오는 괴사 형태의 손상은 NMDA 자극 시와 마찬가지로 Ca2+ 의 세포 내로의 유입에 의해 일어나며, 나머지 뇌부분에서는 apoptosis 양상으로 서서히 뇌손상을 나타낸다 하였는데 (Garthwaite 등, 1991), 이는 본 연구에서와 같은 대뇌피질 신경원을 사용한 연구는 아니었다. Gwag 등은 쥐의 피질신경원 배양 모델에서 3-100 µM AMPA 자극을 주며 24 시간 관찰한 결과 대부분 necrosis 형태의 손상이 오나, apoptosis 형태의 손상이 혼재되어 있을 가능성을 배제하지 못한다 하였으며 (Gwag 등, 1997), Koh 등은 쥐의 피질신경원 배양에 모델에서 500 µM AMPA 5 분 자극을 주어 세포손상을 주지 못하였지만, 5 µM AMPA 20-24 시간 자극은 용량의존적으로 lactic dehydrogenase (LDH, necrosis 형태의 세포손상 시 증가) 증가와 세포손상 면적을 증가시켜 강력한 뇌독성을 나타내었다 하였으며, 짧은 고농도의 자극보다 긴
저농도의 자극이 뇌독성을 나타내는 이유는 AMPA 수용체가 NMDA 수용체와 달리Ca2+ 투과성이 낮기 때문이라 하였다(Koh 등, 1990). 본 연구에서는 Gwag, Koh 등의 연구에서와 같은 백서 피질신경원 배양모델을 사용하였는데, 8 또는 24 시간 동안 30-100 µM AMPA 자극을 통한 예비실험 결과 세포사율이 높지 않아 72 시간 동안 50 µM 의 AMPA 자극주어 49.31%의 세포사율을 보였다. 또한, 전체 신경 세포중 apoptosis 일으킨 비율 28.01% 이어서 결국 세포사를 일으킨 신경원 중 apoptosis 에 의한 세포사는 56.8% 정도였다. LDH 측정 등 necrosis 형태의 세포손상을 보는 실험을 하지 않아 정확하지는 않지만, 위의 연구들로 미루어볼 때 necrosis 와 apoptosis 가 혼재된 형태의 세포사가 일어난 것으로 추정된다. 또한, Larm 등도 백서의 피질 신경원세포 배양모델에서 100-300 µM 의 (S)–AMPA 를 24 시간 투여하여본 결과 50% 정도의 세포에서 apoptosis 형태의 세포사를 일으킨다 하였는데 (Larm 등, 1997), 이는 본 연구의 결과보다 높은 수치이며, AMPA 자극 농도와 시간이 다르기 때문이
뇌보호를 위해 MK-801 과 같은 NMDA 수용체 길항제 들이 많이 연구되어 왔었는데 (Choi 등, 1990), NMDA 수용체 길항제의 여러가지 부작용으로 인해 GYK-52466 같은 AMPA/kainite 수용체 길항제들에 대한 연구, NMDA 와 AMPA 길항제의 혼합투여에 대한 연구들이 이루어지고 있다(Strasser 등, 1995; Arias 등, 1999).
본 연구에서 사용한 propofol 은 흔히 쓰이는 정맥마취제이며, 뇌보호 효과에 대해서는 in vivo 와 in vitro 에서 긍정적 혹은 부정적 의견들이 있다(Kochs 등, 1992; Hans 등, 1994; Orser 등, 1995; Yamakura 등, 1995; Zhu 등, 1997; Young 등, 1997; Peters 등, 2001; Shibuta 등, 2001; Engelhard 등, 2004; Chen 등, 2008).
Propofol 의 뇌보호 기전에 대해서는 여러가지 기전들이 제시되었는데, 두개내압의 감소, 뇌대사율의 감소, 뇌의 산소요구량 감소 (Kochs 등, 1992), glutamate 수용체에 길항제로서의 작용 (Hans 등, 1994; Orser 등, 1995; Yamakura 등, 1995), Na+ 이온채널 의존적 glutamate 분비 감소 (Ligamaneni 등, 2001), glutamate
흡수의 증가 (Peters 등, 2001; Velly 등, 2003), 세포외 glutamate 농도의 감소, 항산화제로 작용하여 유리기 제거자(free radical scavenger)로 작용하는 기전 (Sagara 등, 1999)등이 보고되고 있다.
Yamakura 등은 recombinant glutamate 수용체 channel 을 이용한 연구에서,
AMPA 수용체의 subtype 에 따라 propofol 에 대한 반응이 억제되기도 하고
증가되기도 함을 보고하였는데, 억제효과는 500 μM 의 고농도(임상농도: ~35 μM)
propofol 에서만 일어났고 임상용량에서는 억제효과를 나타내지 않았다(Yamakura
등, 1995). 이는 50 μM 의 propofol 투여가 생존세포수를 47.04 % 에서 65.13 %로
증가시킨 본 연구 결과와는 일치하지 않는다.
Krampfl 등은 recombinant AMPA-type glutamate 수용체 channel 을 이용하여 patch
clamp 실험을 한 결과 100 μM 이상의 propofol 은 AMPA 수용체의 탈감작에
관여하여 AMPA 자극을 오히려 강화한다 하였으며 (Krampfl 등, 2005), Zhu 등은
112 μM 의 propofol 이 AMPA 로 유도된 뇌독성의 증감에 아무 영향도 미치지
않는다 하였는데 (Zhu 등, 1997), 이들 결과들도 본 연구와 일치하지 않았다.
Young 등은 백서를 이용한 in vivo 뇌허혈-재관류 실험에서 propofol 을 뇌허혈
49.31% 에서 29.38%로 낮춘 본 연구의 신경세포 손상에 대한 보호 효과와
비슷하였으나, in vivo 와 in vitro 실험 간의 차이가 있으므로 단순 비교는 어렵다.
Engelhard 등은 백서의 허혈-재관류 모델에서 propofol 이 허혈 4 시간 후에
necrosis 뿐 아니라 apoptosis 를 유도하는 단백질인 Bax 의 농도를 감소시킨다
보고하였으며 (Engelhard 등, 2004), Chen 등은 백서 국소 허혈-재관류 모델에서 propofol 이 apoptosis 에 의한 뇌독성에 미치는 영향을 시간대 별로 살펴본 결과, 국소허혈-재관류는 necrosis 와 apoptosis 가 혼재된 형태의 뇌손상을 일으키는데, apoptosis 는 재관류 6 시간 후부터 일어나기 시작하여 3 일 후까지 존재하다 감소하며, propofol 은 뇌경색면적을 감소시키는 등 뇌보호 효과가 있으며, apoptosis 감소에도 효과가 있었다 보고하였다( Chen 등, 2008). 뇌허혈-재관류 시 glutamate 가 상승하며, 이 중 일부는 AMPA 수용체를 자극하여 necrosis 혹은 apoptosis 형태의 뇌세포 손상이 온다 가정할 경우
(Garthwaite 등, 1991; Garthwaite 등 1991), propofol 50 μM 첨가 시 AMPA 자극에
본 연구의 결과와 비슷하였다. 그러나, 본 연구에서 보인 apoptosis 의 억제가
AMPA 수용체 간의 직접적인 상호작용에 의한 것인지, Engelhard 나 Chen 등의
연구에서와 같이(Engelhard 등, 2004; Chen 등, 2008) propofol 이 AMPA 자극에 의해
유도된 apoptosis 과정 중 일부에 관여해서 인지는 확실치 않으며, 추가 연구가 필요하다. 백서 피질 신경원 배양 모델에서 72 시간 동안 AMPA50 μM 자극과 동시에 0.1-50 μM 농도의 propofol 을 투여하여 propofol 이 뇌보호 효과가 있는지, 72 시간 동안 AMPA 50 μM 자극 시 apoptosis 형태의 신경세포 손상을 얼마나 일으키며, propofol 을 주면 어느 정도 감소 효과가 있는지 살펴본 결과, 49.31%의 세포사율을 보였으며 50 μM propofol 은 세포사율을 29.38%로 감소시켰고, 세포사율 감소 원인 중 일부는 propofol 에 의한 apoptosis 감소에 기인한다 생각되어지며, 추후 뇌보호 효과의 기전 등에 관한 추가 연구가 이루어져야겠다.
V. 결론
백서 피질 신경원 혼합배양 모델에서 72 시간, 50 μM AMPA 자극은 49.31%의 세포사율과 28%의 apoptosis 비율을 보여, necrosis 와 apoptosis 가 혼합된 형태로 추정되는 세포사멸을 일으켰으며, 50 μM propofol 의 처치는 세포사율을 29.38%로, apopotosis 비율을 20%로 의의있게 낮추어 뇌보호 효과가 있었으며, 신경 세포내로의 calcium 이동의 억제가 기전 중 하나로 제시될 수 있겠다. 또한, necrosis 가 어느 정도 관여하는지, apoptosis 의 기전 중 어느 부위에 관여하는지, 증가된 calcium 의 기원이 무엇인지 알려면 추가 연구가 필요하다.
참고 문헌
1. Ankarcrona M, Dypbukt JM., Bonfoco E, Zhivotovsky B, Orrenius S, Lipton SA, Nicotera P: Glutamate-induced neuronal death: a succession of necrosis or apoptosis depending on mitochondrial function. Neuron 15: 961–973, 1995
2. Arias RL, Tasse JR, Bowlby MR: Neuroprotective interaction effects of NMDA and AMPA receptor antagonists in an in vitro model of cerebral ischemia. Brain Res 816(2): 299-308, 1999
3. Benveniste H, Drejer J, Schousboe A, Diemer NH: Elevation of the extracellular concentrations of glutamate and aspartate in rat hippocampus during transient cerebral ischemia monitored by intracerebral microdialysis. J Neurochem 43(5): 1369-1374, 1984
4. Bonfoco E, Krainc D, Ankarcrona M, Nicotera P, Lipton SA: Apoptosis and necrosis: two distinct events induced, respectively, by mild and intense insults with N-methyl-D-aspartate or nitric oxide/superoxide in cortical cell cultures. Proc Natl Acad Sci U S A. 92(16): 7162-7166, 1995
5. Burnashev N, Khodorova A, Jonas P, Helm PJ, Wisden W, Monyer H, et al: Calcium-permeable AMPA-kainate receptors in fusiform cerebellar glial cells. Science 256(5063): 1566-1570, 1992
6. Chen L, Xue Z, Jiang H: Effect of propofol on pathologic time-course and apoptosis after cerebral ischemia-reperfusion injury. Acta Anaesthesiol Scand 52(3): 413-419, 2008
7. Choi DW: Ionic dependence of glutamate neurotoxicity. J Neurosci 7: 369–379, 1987
8. Choi DW, Rothman SM: The role of glutamate neurotoxicity in hypoxic-ischemic neuronal death. Annu Rev Neurosci 13: 171-182, 1990
9. Choi DW: Methods for antagonizing glutamate neurotoxicity. Cerebrovasc Brain Metab Rev 2(2): 105-147, 1990
11. Dingledine R, Borges K, Bowie D, Travnelis SF: The glutamate receptor ion channels. Pharmacol Rev 51(1): 7-61, 1999
12. Engelhard K, Werner C, Eberspächer E, Page M, Blobner M, Hutzler P, et al: Sevoflurane and propofol influence the expression of apoptosis-regulating proteins after cerebral ischaemia and reperfusion in rats. Eur J Anaesthesiol 21(7): 530-537, 2004
13. Frandsen A, Dreier J, Schousboe A: Direct evidence that excitotoxicity in cultured neurons is mediated via N-methyl-D-aspartate (NMDA) as well as non-NMDA receptors. J Neurochem 53(1): 297-299, 1989
14. Garthwaite G, Garthwaite J: AMPA Neurotoxicity in Rat Cerebellar and Hippocampal Slices: Histological Evidence for Three Mechanisms. Eur J Neurosci 3(8): 715-728, 1991
15. Gaerthwaite G, Garthwaite J: Mechanisms of AMPA Neurotoxicity in Rat Brain Slices. Eur J Neurosci 3(8): 729-736, 1991
16. Goldberg MP, Weiss JH, Pham PC, Choi DW: N-methyl-D-aspartate receptors mediate hypoxic neuronal injury in cortical culture. J Pharmac Exp Ther 243: 784-791, 1987
17. Gunasekar PG, Kanthasamy AG, Borowitz JL, Isom GE: NMDA Receptor Activation Produces Concurrent Generation of Nitric Oxide and Reactive Oxygen Species: Implications for Cell Death. J Neurochem 65: 2016-2021, 1995
18. Gwag BJ, Koh JY, DeMaro JA, Ying HS, Jacquin M, Choi DW: Slowly triggered excitotoxicity occurs by necrosis in cortical cultures. Neuroscience 77: 393–401, 1997
19. Hans P, Bonhomme V, Collette J, Albert A, Moonen G: Propofol protects cultured rat hippocampal neurons against N-methyl-D-aspartate receptor-mediated glutamate toxicity. J Neurosurg Anesthesiol 6: 249–253, 1994
20. Hollmann M, Hartley M, Heinemann S: Ca2+ permeability of KA-AMPA-gated glutamate receptor channels depends on subunit composition. Science 252(5007): 851-853, 1991
21. Iino M, Ozawa S, Tsuzuki K: Permeation of calcium through excitatory amino acid receptor channels in cultured rat hippocampal neurones. J Physiol 424: 151-165, 1990
22. Jonas P, Racca C, Sakmann B, Seeburg PH, Monyer H: Differences in Ca2+
permeability of AMPA-type glutamate receptor channels in neocortical neurons caused by differential GluR-B subunit expression. Neuron 12(6): 1281-1289, 1994
23. Kochs E, Hoffmann WE, Werner C, Thomas C, Albrecht RF, am Esch JS: The effects of propofol on brain electrical activity: neurologic outcome and neuronal damage following incomplete ischemia in rats. Anesthesiology 76: 245-252, 1992
24. Koh JY, Goldberg MP, Hartley DM, Choi DW: Non-NMDA receptor-mediated neurotoxicity in cortical culture. J Neurosci 10(2): 693-705, 1990
25. Krampfl K, Cordes AL, Schlesinger F, Wolfes H, Bufler J: . Effects of propofol on recombinant AMPA receptor channels. Eur J Pharmacol 511(1): 1-7, 2005
26. Larm JA, Cheung NS, Beart PM: Apoptosis induced via AMPA-selective glutamate receptors in cultured murine cortical neurons. J Neurochem 69(2): 617-622, 1997
27. Lazarewicz JW, Wroblewski JT, Costa E: N-methyl-D-aspartate-sensitive glutamate
receptors induce calcium-mediated arachidonic acid release in primary cultures of cerebellar granule cells. J Neurochem 55: 1875–1881, 1990
28. Li C, Jackson RM: Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am J Physiol Cell Physiol 282(2): C227-241, 2002
29. Ligamaneni R, Birch ML, Hemmings Jr HC: Widespread inhibition of sodium channel-dependent glutamate release from isolated nerve terminals by isoflurane and propofol. Anesthesiology 95(6): 1460-1466, 2001
30. Madden DR: The structure and function of glutamate receptor ion channels. Nat Rev Neurosci 3(2): 91-101, 2002.
31. Nishizawa Y: Glutamate release and neuronal damage in ischemia. Life Sci 69: 369-381, 2001.
32. Orser BA, Bertlik M, Wang LY, MacDonald JF: Inhibition by propofol (2,6 di-isopropylphenol) of the N-methyl-D-aspartate subtype of glutamate receptor in cultured hippocampal neurones. Br J Pharmacol 116: 1761–1768, 1995
33. Pellegrini-Giampietro DE, Zukin RS, Bennett MV, Cho S, Pulsinelli WA:Switch in glutamate receptor subunit gene expression in CA1 subfield of hippocampus following global ischemia in rats. Proc Natl Acad Sci U S A 89(21): 10499-10503, 1992
34. Pellegrini-Giampietro DE, Gorter JA, Bennett MV, Zukin RS:The GluR2 (GluR-B) hypothesis: Ca(2+)-permeable AMPA receptors in neurological disorders. Trends
Neurosci 20(10): 464-470, 1997
35. Peters CE, Korcok J, Gelb AW, Wilson JX: Anesthetic concentrations of propofol protect against oxidative stress in primary astrocyte cultures. Anesthesiology 94: 313-321, 2001
36. Rothman SM, Thurston JH, Hauhart RE: The neurotoxicity of excitatory amino acids is produced by passive chloride influx. J Neurosci 5(6): 1483-1489, 1985
37. Rothman SM, Olney JW: Glutamate and the pathophysiology of hypoxic-ischemic brain damage. Ann Neurol 19: 105-111, 1986
38. Sagara Y, Hendler S, Khoh-Reiter S, Gillenwater G, Carlo D, Schubert D, Chang J: Propofol hemisuccinate protects neuronal cells from oxidative injury. J Neurochem 73(6): 2524-2530, 1999
39. Seeburg PH: The role of RNA editing in controlling glutamate receptor channel properties. J Neurochem 66(1): 1-5, 1996
40. Shibuta S, Sriranganath V, Inoue T, Shimizu T, Tomi K, Mashimo T: The effects of propofol on NMDA or nitric oxide mediated neurotoxicity in vitro. Neuroreport 12(2): 295-298, 2001
41. Strasser U, Fischer G: Protection from neuronal damage induced by combined oxygen and glucose deprivation in organotypic hippocampal cultures by glutamate receptor antagonists. Brain Res 687(1-2): 167-174, 1995
42. Velly LJ, Guillet BA, Masmejean FM, Nieoullon AL, Brauder NJ, Gouin FM, Pisano PM: Neuroprotective effects of propofol in a model of ischemic cortical cell
cultures: role of glutamate and its transporters. Anesthesiology 99(2): 368-375, 2003
43. Watkins JC, Olverman HJ: Agonists and antagonists for excitatory amino acid receptors. Trends Neurosci 10: 265-272, 1987
44. Yamakura T, Sakimura K, Shimoji K, Mishina M: Effects of propofol on various AMPA-, kainite- and NMDA-selective glutamate receptor channels expressed in Xenopus oocytes. Neurosci Lett 188: 187–190, 1995
45. Young Y, Menon DK, Tisavipat N, Matta BF, Jones JG: Propofol neuroprotection in a rat model of ischemia reperfusion injury. Eur J Anesthesiol 14: 320-326, 1997
46. Zhu H, Cottrell JE, Kass IS: The effect of thiopental and propofol on NMDA- and AMPA-mediated glutamate excitotoxicity. Anesthesiology 87: 944–951, 1997
- ABSTRACT -
The Effects of propofol on
α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) induced neurotoxicity in rat mixed
cortical cultures
Myung-Sin Seo
Department of Medical Sciences The Graduate School, Ajou University
(Supervised by Associate Professor Sook-Young Lee)
Background: Excessive extracellular glutamate accmulation due to cerebral ischemia is belived to induce excitotoxic cascade by overstimulation of glutamate receptors such as N-methyl-D-aspartate (NMDA), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
(AMPA), and kainite. The pattern of AMPA mediated neurotoxicity (necrosis vs apoptosis) and the neuroprotective effect of propofol to AMPA mediated neurotoxicity still remain unclear. We applied 72 hr, 50 µM AMPA stimulation to cultured rat cortical neurons and have examined the pattern of AMPA neurotoxicity and the neuroprotective effects of propofol. Also, we have examined the effect of propofol on AMPA mediated intraneural calcium influx.
Methods: Sprague-Dawley rats (250-300 g) were used. Thirteen-day-old primary mixed cortical cultures were exposed to 50 µM AMPA stimulation (AMPA group). Propofol 0.1, 1, 25, 50, μM were co-administered with 50 µM AMPA (propofol-treated group) or propofol 50 μM alone. In control and Full-kill group, no drug or NMDA 100 μM were administered. 72 hr after, the survived cells were counted using trypan-blue staining. And, the cell death rate (CDR) were calculated (control group CDR set as 0, Full kill group CDR set as 100). After confirming the presence of neuronal cells by neuronal nuclei(NeuN) and Hoechst stain, we quantified apoptotic cells by terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end-labeling (TUNEL) assay. The effect of propofol on AMPA mediated intracellular calcium increase was measured with calcium-sensitive fluorochrome calcium green-5N-acetoxymethylester with confocal laser scanning microscopy. Statistical analysis was done by Oneway-ANOVA test and Bonferroni test. P<0.05 was considered as statistically significant.
Results: AMPA 50 μM stimulation demonstrated 49.31% CDR and adding propofol 50 μM
decreased CDR to 29.38% (P<0.05). In TUNEL assay, cells with no drug treatment demonstrated 12.33% apoptotic cell rate (ACR) and 50 μM AMPA incrcresed ACR to
28.01% (P<0.05). Adding 50 μM propofol to AMPA decreased the ACR to 20% (P<0.05). In
calcium imaging, the maximum intracellular calcium influx was decreased in AMPA 50 μM
+ propofol 50 μM group compared to AMPA 50 μM group (3164.98 nA vs 2276.98 nA,
respectively)(P<0.05).
Conclusion: 50 μM propofol demonstrated neuroprotective effects against AMPA 50 μM stimulation and the underlying mechanism propofol’s effect may be caused partly by the reduction of the intracellulr calcium influx.