저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Master’s Thesis in Molecular Medicine
Effects of Scopolin on osteoblast
And osteoclast differentiation
Ajou University Graduate School
Major in Molecular Medicine
Department of Biomedical Sciences
Effects of Scopolin on osteoblast
and osteoclast differentiation
Seon-Yong Jeong, Advisor
I submit this thesis as the
Master’s thesis in Molecular Medicine
February 2018
Ajou University Graduate School
Major in Molecular Medicine Department
of Biomedical Sciences
-Abstract-
Effects of Scopolin on osteoblast and osteoclast differentiation
Osteoporosis is a bone disease that increases the risk of fractures owing to low bone mineral density (BMD). An imbalance between bone formation (osteoblasts) and bone resorption (osteoclasts) causes osteoporosis. In our previous study, the anti-osteoporotic effects of Lycii Radicis Cortex (LRC) were reported in vitro and in vivo.
Fractionation and isolation of the bioactive compound responsible for the anti-osteoporotic effects of LRC extract were performed and Scopolin was identified as a candidate component. Next, the osteoporotic effects of Scopolin were investigated in osteoblasts and osteoclasts. Scopolin significantly increased alkaline phosphatase (ALP) activity and ALP positive-stained colonies expressed in the osteoblastic cell line MC3T3-E1 as compared to non-treated cells, indicating an increase in osteoblast differentiation. The mRNA levels of osteoblast markers such as Alp, Runx2, Bglap,
and Sp7 as well as mineralized nodule formation significantly increased in
Scopolin-treated MC3T3-E1 cells.
In addition, Scopolin reduced tartrate-resistant acid phosphatase (TRAP) activity and TRAP-positive staining of primary-cultured monocytes derived from mouse
ii
of osteoclast differentiation markers such as Tnfsf11 and Nfatc1 significantly decreased. Consistently, Scopolin induced osteoblast differentiation but reduced osteoclast differentiation upon co-culture with osteoblastic MC3T3-E1 and primary cultured monocyte cells.
In the in vivo experiments using ovariectomized (OVX) mice as an osteoporosis animal model, the Scopolin-administered OVX mouse group showed significantly reduced BMD loss as compared to the OVX control mouse group. The microcomputed tomography and single photon emission computed tomography results indicate that Scopolin prevented trabecular bone loss and osteoclastogenesis in OVX mice. In addition, the serum OPG/RANKL ratio, which is used as a bone formation marker, was higher in the Scopolin-administered group than in the OVX control group.
Collectively, the results of this study indicate that Scopolin effectively enhances bone formation and reduces bone resorption during the bone remodeling process and suggest that Scopolin might be a potential therapeutic agent for the treatment of osteoporosis.
__________________________________________________________________ Keywords: Scopolin, Lycii Radicis Cortex, osteoporosis, osteoblast, osteoclast, ovariectomized mice, bone mineral density
TABLE OF CONTENTS
ABSTRACT ··· i
TABLE OF CONTENTS ··· iii
LIST OF FIGURES ··· v
I. INTRODUCTION ··· 1
II. MATERIALS AND METHODS ··· 6
A. Fractionation, isolation, and structural elucidation of the bioactive component ··· 6
B. Reagents and cell culture materials ··· 10
C. Water-soluble tetrazolium salt (WST) assay ··· 10
D. Osteoblast bone formation and ALP assay ··· 10
E. Mineralized nodule formation in osteoblast cells ··· 11
F. TRAP staining and activity ··· 11
G. Quantitative reverse-transcription PCR (qRT-PCR) ··· 12
H. In vitro generation of osteoclasts and the coculture system ··· 13
I. Ovariectomized mouse model ··· 13
J. Measurement of bone mineral density (BMD) ··· 14
K. Microcomputed tomography (Micro-CT) and single photon emission computed tomography (SPECT) scan ··· 14
iv
III. RESULTS ··· 17
A. Scopolin induced osteoblast differentiation and mineralized nodule formation in the osteoblastic cell line MC3T3-E1 ··· 17
B. Scopolin increased the mRNA expression of Alp, Runx2, Bglap, and Sp7 osteoblastic markers ··· 20
C. Scopolin decreased differentiation of osteoclast cells isolated from bone marrow ··· 22
D. Scopolin suppressed the mRNA expression of the osteoclastic markers Tnfsf11 and Nfatc1 ··· 25
E. Scopolin increased osteoblast differentiation and decreased osteoclast differentiation in a co-culture of isolated monocytes and the MC3T3-E1 osteoblastic cell line ··· 27
F. Scopolin prevented OVX-induced bone loss in mice ··· 29
IV. DISCUSSION ··· 35
V. CONCLUSION ··· 40
REFERENCES ··· 41
LIST OF FIGURES
Figure 1. Bone remodeling ··· 5
Figure 2. Fractionation and isolation of the bioactive component fraction enhancing osteoblast differentiation from 70% ethanol extract of Lycii Radicis
cortex ··· 7
Figure 3. Analysis of nuclear magnetic resonance of the D2-5 fraction ··· 8
Figure 4. Chemical structure of the isolated Scopolin ··· 9
Figure 5. The effects of Scopolin on cellular differentiation, proliferation, and mineralized nodule formation in the osteoblast cell line MC3T3-E1. ··· 18
Figure 6. The mRNA expression of osteoblastic markers Alp, Runx2, Bglap, and
Sp7 in MC3T3-E1 treated with Scopolin ··· 21
Figure 7. Inhibition of osteoclast differentiation in monocytes isolated from bone marrow treated with Scopolin. ··· 23
vi
Figure 8. The mRNA expression of the osteoclastic markers Tnfsf11 and Nfatc1 · 26
Figure 9. Differentiation of osteoblasts and osteoclasts in a co-culture of isolated monocytes and the osteoblastic MC3T3-E1 cell line ··· 28
Figure 10. Inhibition of OVX-induced bone loss in mice treated with Scopolin ··· 31
I. INTRODUCTION
Bone is an important body tissue; its functions include holding and protecting major organs and regulating the homeostasis of minerals (Pacifici, 2017). Bone consists of three kinds of cells: osteoblasts, osteoclasts, and osteocytes (Florencio-Silva et al., 2015). Osteoblasts, or bone-forming cells, are found at the surface of the bone and are stimulated to proliferate and differentiate into osteocytes (Florencio-Silva et al., 2015). Osteoclasts are bone-resorbing cells derived from stem cells in the bone marrow that also give rise to monocytes and macrophages (Florencio-Silva et al., 2015). Osteocytes derived from osteoblasts play an important role in bone function and synthesis of type 1 collagen and other substances that make up the bone extracellular matrix (Florencio-Silva et al., 2015).
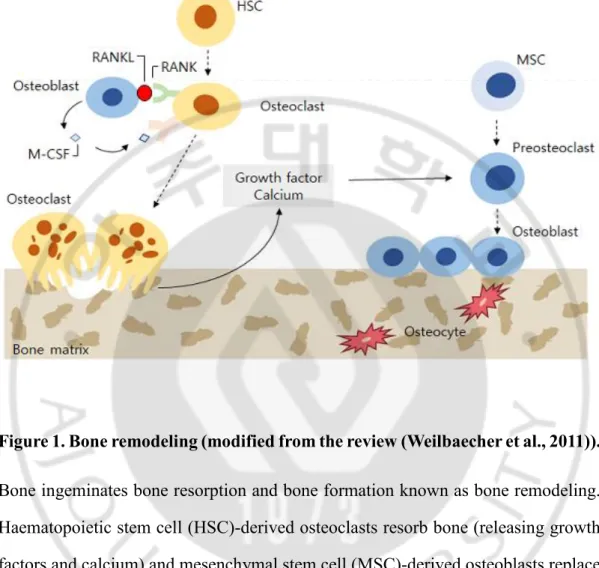
Unlike other connective tissues, bone undergoes mineralization and remodeling throughout its lifetime (Katsimbri, 2017). The process of bone remodeling comprises bone formation (osteoblasts) and bone resorption (osteoclasts) (Raggatt and Partridge, 2010) (Figure 1). Osteoblasts and osteoclasts interact with each other during bone remodeling. Mesenchymal stromal cells (MSCs), preosteoblasts, and osteoblasts produce cytokines that affect preosteoclasts and osteoclasts in the bone marrow. Cytokines such as interleukin (IL)-1 and -6 and tumor necrosis factor (TNF)-α from osteoblasts control osteoclast differentiation by activating specific
- 2 -
growth factor-β (TGF-β) produced by osteoblasts controls osteoclast function. During bone resorption, TGF-β protects osteoclasts from apoptosis and promotes osteoclast differentiation and bone resorption (Wang et al., 2005; Mohammad et al., 2009). Furthermore, preosteoblasts and osteoblasts express receptor activator toward nuclear factor κB ligand (RANKL), a molecule that binds to the receptor activator of nuclear factor κB (RANK) on preosteoclast cells and stimulates osteoclast differentiation (Zhang et al., 2017). Preosteoblasts and osteoblasts also release osteoprotegerin (OPG), which blocks RANKL interaction with its ligand and inhibits osteoclast differentiation (Fu et al., 2013).
Bone formation is regulated by mineralization (Gough et al., 2004), collagen synthesis, osteoblast differentiation (Ducy et al., 2000), and alkaline phosphatase (ALP) activity (Watts, 1999). Bone resorption is associated with osteoclast differentiation, tartrate-resistant acid phosphatase (TRAP) activity (Salamanna et al., 2016), and expression of Tnfsf11 and Nfatc1 (Nakagawa et al., 2015). During bone remodeling, old bone tissues are removed by the action of osteoclasts and the removed tissue parts are filled with new bone tissues by osteoblasts. An imbalance between bone formation (osteoblasts) and bone resorption (osteoclasts) causes many bone-related diseases.
Osteoporosis is a progressive bone disease often seen in postmenopausal women, which increases bone resorption and causes reduction of bone mass and density, consequently leading to an increased risk of bone fracture (Lacombe et al., 2016).
Approximately 100 million people have debilitating bone disease worldwide, especially postmenopausal women (Johnell and Kanis, 2005). In postmenopausal women, Estrogen which plays important roles like maintenance of bone formation and inhibition of bone resorption has been reported to be frequently reduced (Zeng et al., 2015). Key risk factors for osteoporosis include genetics, lack of calcium and vitamin D, cigarette smoking, excessive alcohol consumption, history of rheumatoid arthritis, and low body weight (Rosen, 2000). Osteoporosis can be diagnosed with the use of X-rays and confirmed by tests to measure bone density. Treatments for osteoporosis, in addition to prescription osteoporosis medications, include stopping use of alcohol and cigarettes and adequate exercise and intake of calcium and vitamin D (Ensrud and Crandall, 2017).
Herbal medicines have been used as an alternative therapy for bone heath and osteoporosis (Liu et al., 2017). Many studies have suggested that ingredients derived from natural plants have potential uses as herbal medicines (Kim et al., 2016; Jin et al., 2017). Lycii Radices Cortex (LRC), Lycium chinense root bark, has been used as a traditional medicine in eastern Asia (Potterat, 2010; Kim et al., 2016). LRC extract has beneficial effects on insulin resistance and secretion, lipid metabolism in liver steatosis, antioxidation, and the immune system (Ye et al., 2008). In addition, our previous study suggested that LRC has therapeutic effects on the differentiation of osteoblasts in vitro and in vivo (Park et al., 2014). Despite findings that extract of
- 4 -
In this thesis, I investigated whether an anti-osteoporotic compound, Scopolin, isolated from LRC extract has potential to be used for osteoporosis treatment. First, osteoblastic cells (MC3T3-E1) were treated with Scopolin and its effects on osteoblast differentiation were tested by analyzing ALP activity, ALP staining, Alizarin red S staining, and mRNA levels of osteoblast markers (Alp, Runx2, Bglap, and Sp7). Second, I assessed the effects of Scopolin on osteoclasts. Preosteoclast monocyte cells were isolated from mouse bone marrow and treated with Scopolin. Osteoclastic differentiation of primary cultured monocytes was measured by evaluating TRAP activity, and Scopolin was found to decrease TRAP activity in a dose-dependent manner. In addition, TRAP-positive colonies and mRNA levels of osteoclast inhibitory markers (Tnfsf11 and Nftac1) were suppressed in osteoclasts.
Next, I identified the role of Scopolin under conditions where osteoblast and osteoclast cells coexist by using a co-culture of preosteoclast monocytes and preosteoblast MC3T3-E1 cells. Scopolin enhanced osteoblast differentiation (ALP activity) and suppressed osteoclast differentiation (TRAP activity) in the co-culture system. Finally, OVX-induced bone mineral density (BMD) loss was recovered by Scopolin treatment in an osteoporotic mouse model. In addition, analysis of 2D and 3D micro-CT images of the right femur in Scopolin-treated OVX mice showed significantly improved bone volume (BV/TV), thickness (Tb.Th), trabecular number (Tb.N), and trabecular separation (Tb.Sp) as compared to the OVX control. The present study suggests that Scopolin could be a therapeutic candidate for osteoporosis.
Figure 1. Bone remodeling (modified from the review (Weilbaecher et al., 2011)). Bone ingeminates bone resorption and bone formation known as bone remodeling. Haematopoietic stem cell (HSC)-derived osteoclasts resorb bone (releasing growth factors and calcium) and mesenchymal stem cell (MSC)-derived osteoblasts replace it with new bone (Weilbaecher et al., 2011). When osteoblasts are trapped in the bone matrix, they become osteocytes. Osteoblasts express RANKL, a molecule that binds to RANK on preosteoclast cells and stimulates osteoclast differentiation (Zhang et al., 2017).
- 6 -
II. MATERIALS AND METHODS
A. Fractionation, isolation, and structural elucidation of the bioactive component.
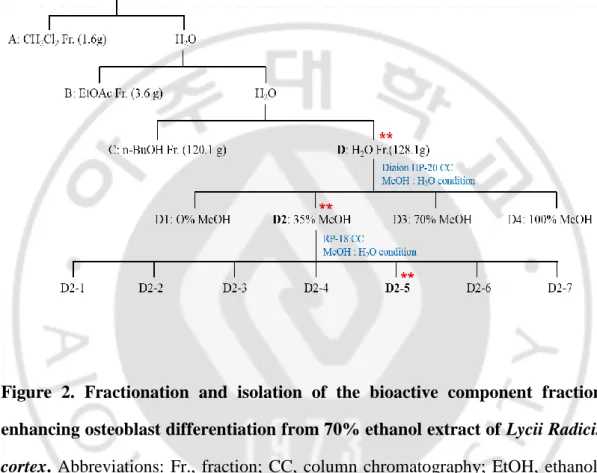
A 70% ethanol extract of LRC (254 g) was evaporated, suspended in H2O, and then partitioned successively with CH2Cl2 (A, 1.6 g), EtOAc (B, 3.6 g), n-BuOH (C, 120.1 g), and aqueous (D, 128.1 g) to obtain the respective fractions. Fraction D was chromatographed over a Diaion HP-20 gel (1000 g) column and eluted with a gradient H2O-MeOH solvent system (0% MeOH, 35% MeOH, 70% MeOH, and 100% MeOH) to give four fractions (D1–D4). Fraction D2 (14.5 g) was subjected to RP-18 gel (200 g) column chromatography and eluted with H2O-MeOH (100:0 to 0:100) to afford seven subfractions (D2-1 to D2-7). Subfraction D2-5 (524 mg) was subjected to preparative HPLC and eluted with MeOH-H2O/0.1% formic acid (10:90) to yield Compound 1 (2.1 mg) (Figure 2). ALP activity was measured in each fraction for assessment of osteoblast differentiation effect. The structure of the isolated compound was elucidated by 1H-NMR, 13C-NMR, and mass spectrometry analyses (Figure 3). As a result, Scopolin (formula: C16H18O9) was identified (Figure 4).
Figure 2. Fractionation and isolation of the bioactive component fraction enhancing osteoblast differentiation from 70% ethanol extract of Lycii Radicis cortex. Abbreviations: Fr., fraction; CC, column chromatography; EtOH, ethanol; CH2Cl2, dichloromethane; EtOAc, ethyl acetate; n-BuOH, n-butanol; MeOH, methanol. **: Bioactivity-guided fraction.
- 8 -
Figure 3. Analysis of nuclear magnetic resonance of the D2-5 fraction. Results of proton nuclear magnetic resonance(1H-NMR) (A), carbon-13 nuclear magnetic resonance (13C-NMR) (B), and mass spectrum (C) analyses of the D2-5 fraction of Figure 1.
A
B
Figure 4. Chemical structure of the isolated Scopolin. 6-methoxy-7-[(2S, 3R, 4S, 5S, 6R)-3, 4, 5-trihydroxy-6-(hydroxymethyl) oxan-2-yl] oxychromen-2-one
- 10 - B. Reagents and cell culture materials.
Mouse MC3T3-E1 (preosteoblast) cells were purchased from the RIKEN Cell Bank (Tsukuba, Japan) and cultured in α-modified minimal essential (α-MEM) medium, supplemented with 10% FBS, penicillin (100 U/mL), and streptomycin (100 μg/mL). Osteoblast differentiation was induced by adding osteogenic medium containing ascorbic acid (50 μg/mL) and β-glycerophosphate (10 mM) after allowing 24 h for cell adherence (day 0). All cultured cells were incubated in a humidified atmosphere at 37° C and 5% CO2.
C. Water-soluble tetrazolium salt (WST) assay.
Osteoblast cells were incubated in a 96-well plate overnight and co-treated with different concentrations of Scopolin (1, 5, and 10 μM) in the medium for 48 h. The cell viability was determined by WST assay. WST solution (20 μL, 5 mg/mL in PBS) was added to each well, the cells were incubated for another 4 h, and the media were carefully removed. Formazan crystals were dissolved in acidified isopropyl alcohol (40 mM HCl in isopropanol), and their absorbances were measured at 450 nm and 655 nm using a microplate reader (BioTek, Winooski, USA).
D. Osteoblast bone formation and ALP assay.
Preosteoblastic mouse cells were treated with three different concentrations of Scopolin (1, 5, and 10 μM) for either 12 or 72 h. After osteoblast induction, ALP activity was measured in total cell lysates after homogenization in buffer containing
1 mmol/L of Tris–HCl (pH 8.8), 0.5% Triton X-100, 10 mmol/L of Mg2+, and 5 mmol/L of p-nitrophenyl phosphate as substrate, and the absorbance was read at 405 nm (BioTek).
E. Mineralized nodule formation in osteoblast cells.
MC3T3-E1 osteoblast cells were cultured in a 48-well plate and treated with Scopolin for 21 days. The mineralization of the matrix and nodule formation were determined by Alizarin red S (Sigma-Aldrich; St. Louis, USA) staining. The medium was removed and gently washed with PBS. The cells were fixed in cold 70% ethanol for 10 min, and calcium deposits in the mineralized cells were assessed by staining with 40 mM of Alizarin red S (Sigma-Aldrich). Positive Alizarin red S staining for calcium was determined under a light microscope.
F. TRAP staining and activity.
Preosteoclast monocyte cells were isolated from bone marrow cells of the femurs of 7-week-old mice in the presence of 1 mM ascorbate-2-phosphate (Sigma-Aldrich; St. Louis, USA). The isolated monocyte cells were identified by immunophenotypic analysis with a CD11b antibody using an FACS Aria III cell sorter (BD Biosciences, San Jose, USA) and FACS Diva software (BD Biosciences). The monocyte cells were cultured in the presence of 30 ng/mL of M-CSF (PeproTech, Rocky Hill, USA) and 50 ng/mL of RANKL (PeproTech) to induce differentiation to osteoclasts (Sun
- 12 -
incubation of the monocytes, the medium was removed and gently washed with PBS. The cells were fixed in cold 4% paraformaldehyde for 10 min and washed with PBS. The differentiated osteoclast cells from monocytes were measured by TRAP activity assay and staining using an Acid-Phosphatase Kit (Sigma-Aldrich; St. Louis, USA). TRAP-positive multinucleated cells containing three or more nuclei were counted under a light microscope. The absorbance was measured at 405 nm (BioTek), and TRAP activity was expressed as the percent of the untreated control.
G. Quantitative reverse-transcription PCR (qRT-PCR).
Total RNA was extracted from cultured cells using TRIzol reagent (Invitrogen, Carlsbad, USA), following the manufacturer′s instructions (Beckman Coulter, Brea, USA). The extracted RNA was subsequently reverse-transcribed using a RevertAid™ H Minus First Strand cDNA Synthesis Kit (Fermentas Inc., Hanover, USA), with oligo(dT)15–18 as a random primer. All real-time RT-PCR measurements were performed using the ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster City, USA). All PCR amplifications (40 cycles) were performed in a total volume of 10 μL containing 150 ng of cDNA using an SYBR Green I qPCR kit (TaKaRa, Shiga, Japan), according to the manufacturer′s recommendations. The specific primers for osteoblast-inducing markers were as follows: 5′-TCC CAC GTT TTC ACA TTC GG-3′ and 5′-GGC CAT CCT ATA TGG TAA CGG G-3′ for mouse
Alp, 5′-TAA AGT GAC AGT GGA CGG TCC C-3′ and 5′-CCT CAG TGA TTT
5′-ATG GCT TGA AGA CCG CCT ACA-3′ for mouse Bglap, and 5′-ATG GCG TCC TCT CTG CTT G-3′, 5′-TGA AAG GTC AGC GTA TGG CTT-3′ for mouse Sp7. The specific primers for osteoclast inhibitory markers were as follows: 5′-TGC CTA CAG CAT GGG CTT T-3′ and 5′-AGA GAT GAA CGT GGA GTT ACT-3′ for mouse Tnfsf11, 5′-TCA TCC TGT CCA ACA CCA AA-3′ and 5′-TCA CCC TGG TGT TCT TCC TC-3′ for mouse Nfatc1.
By normalizing to Gapdh, relative quantification of gene expression was performed using the comparative threshold (Ct) method, as described by the manufacturer (Applied Biosystems). The values were expressed as the fold change over the control. Relative gene expression was displayed as 2-ΔCt (ΔCt = Ct target gene - Ct Gapdh). The fold change was calculated as 2-ΔΔCt (ΔΔCt = ΔCt control - Ct treatment).
H. In vitro generation of osteoclasts and the coculture system.
The isolated monocyte cells (4 × 104 cells/well) and MC3T3-E1 (2 × 104 cells/well) cells were co-cultured for 5 days in differentiation media containing ascorbic acid (50 µg/mL) and β-glycerophosphate (10 mM) to induce osteoblast differentiation. Osteoblast and osteoclast differentiation were measured by ALP and TRAP activity, respectively.
I. Ovariectomized mouse model.
- 14 -
(Hamamatsu, Japan). Mice were maintained on a diet (5.0 g/day) of Formula-M07 (Feedlab Co., Ltd., Hanam, Korea) and tap water (15 mL/day). All mice were housed individually in clear plastic cages under controlled temperature (23 ± 2° C), humidity (55 ± 5%), and illumination (12 h light/dark cycle). Mice were administered orally with different concentrations of Scopolin (20 or 40 mg/kg/day) using a stomach tube for 12 weeks. The animal research protocol was approved by the Animal Care and Use Committee of the Ajou University School of Medicine, and all experiments were conducted according to the institutional guidelines established by the committee.
J. Measurement of bone mineral density (BMD).
During the in vivo experiment, BMD was measured using a PIXI-mus bone densitometer (GE Lunar, Madison, USA). After anesthetization using tiletamine/zolazepam (Zoletil; Virbac Laboratories, Carros, France), the mice were placed on the specimen tray for measurement. All mice were carefully placed in the same position. Whole body BMC and % fat and right femur BMD were measured using on-board PIXI-mus software for small animals, and adjusted in relation to body weight.
K. Microcomputed tomography (Micro-CT) and single photon emission computed tomography (SPECT) scan.
in the GSTEP (INVEON, SIEMENS, Germany) with a voltage of 60kV, a current of 400 µA, an exposure time of 400 ms, a magnification of M-H, a CCD readout of 1280x1280 and a rotation step of 360. Reconstruction was performed using Inveon Research Workplace and COBRA_Exxim (SIEMNS, Germany). Two-dimensional axial and 3D images were reconstructed for qualitative and quantitative analyses. For region of interest (ROI) measurement and analyses, axial images were displayed using Inveon Research Workplace (Siemens). Axial reformats were performed to allow slice-by-slice manual tracing of the contours of trabecular bone. For cross-sectional study of trabecular bone, a region was analyzed approximately 300 μm below the growth plate. The analysis yielded information about the main histomorphometric parameters, including BV/TV (%) and Tb.N (1/mm), Tb.Th (l mm), and separation Tb.Sp (l mm). For the SEPCT scan, mice were anesthetized by isoflurane/N2O/O2 inhalation anesthesia and injected with Tc-99m HDP (Mallinckrodt Medical B.V, Netherlands) via the tail vein. In vivo planar images were obtained using an Inveon SPECT scanner (Siemens Preclinical Solutions, Malvern, PA, USA) equipped with a low energy all-purpose collimator. The images were acquired until they reached 100,000 counts per body.
L. Serum OPG and RANKL levels.
Serum levels of OPG and RANKL were assessed by multiplex assays using luminex (Merck Millipore, USA), according to the manufacturer’s instructions. OPG is an osteoblast-secreted decoy receptor that functions as a negative regulator of bone resorption (Khosla, 2001). Moreover, RANKL is a ligand for OPG and functions as a key factor for osteoclast differentiation and activation (Khosla, 2001). The ratio of
- 16 -
bone formation coupled with bone resorption (Li et al., 2015).
M. Statistical analysis.
A statistical software package (SPSS 11.0 for Windows, SPSS Inc., Chicago, IL, USA) was used to perform the statistical tests. The statistical significance of differences was assessed by Student′s t-test. A value of P < 0.05 was considered significant. The results were expressed as mean ± SEM.
III. RESULTS
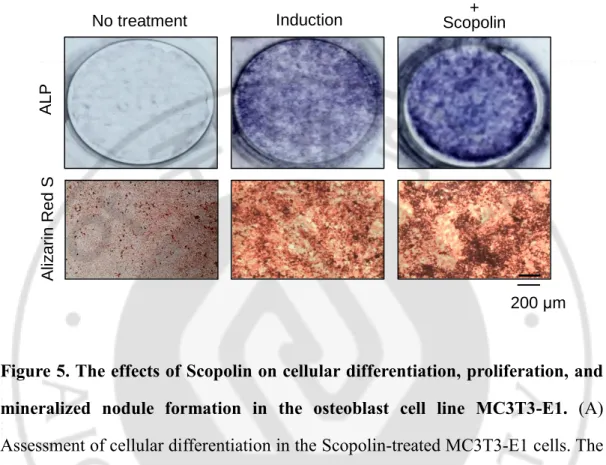
A. Scopolin induced osteoblast differentiation and mineralized nodule formation in the osteoblastic cell line MC3T3-E1.
To investigate the effects of Scopolin on osteoblast differentiation, I evaluated ALP activity, ALP staining, and Alizarin red S staining in the osteoblast cell line MC3T3-E1. Cells were treated with three different concentrations of Scopolin (1, 5, and 10 μM) for either 3 or 5 days. ALP activity at 3 days was significantly higher than that at 5 days, because ALP was observed at an earlier stage of osteoblast differentiation. The highest ALP activity was observed at 5 μM of Scopolin (Figure 5A). However, there was no difference in osteoblast proliferation, indicating no toxic effects of Scopolin on osteoblast cells (Figure 5B). To further confirm the effects of Scopolin on osteoblast differentiation, ALP, and calcium deposits, Alizarin red S staining was measured. The ALP-positive colonies increased in 5 μM Scopolin-treated cells as compared to that in control osteoblast cells. Moreover, MC3T3-E1 cells treated with 5 μM Scopolin for 21 days showed an increase in Alizarin red S-positive cells (Figure 5C). Thus, Scopolin induced osteoblast differentiation and mineralization in MC3T3-E1 cells.
- 18 - Con trol Sco 1 Sco 5 Sco 10 0 50 100 R elat iv e cel l prol if e rat ion
B
A
Day 3 5 0 40 80 120 Control Sco1 Sco5 Sco10*
*
*
R elat iv e A LP ac ti v it yFigure 5. The effects of Scopolin on cellular differentiation, proliferation, and mineralized nodule formation in the osteoblast cell line MC3T3-E1. (A) Assessment of cellular differentiation in the Scopolin-treated MC3T3-E1 cells. The cells were treated with three different concentrations of Scopolin (1, 5, and 10 μM) for 3 days, and ALP activity was measured. (B) Assessment of cellular proliferation in the Scopolin-treated MC3T3-E1 cells. WST was performed to confirm that Scopolin has no effects on cell proliferation. (C) Assessment of cellular differentiation and mineralized nodule formation in Scopolin-treated MC3T3-E1 cells. Mineralized nodule formation in MC3T3-E1 cells treated with 5 μM Scopolin for 21 days was confirmed. Control: non-Scopolin-treated cells. *: p < 0.05 vs.
C
A LP A liz arin R ed S No treatment Induction + Scopolin Induction 200 μm- 20 -
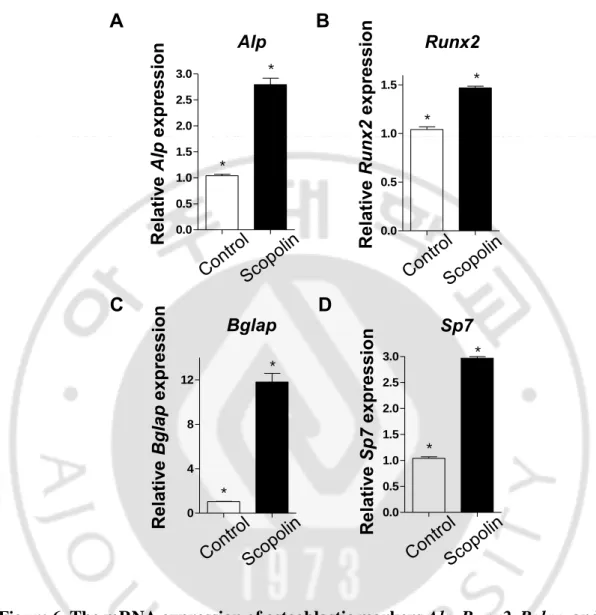
B. Scopolin increased the mRNA expression of Alp, Runx2, Bglap, and Sp7 osteoblastic markers.
To investigate the effects of Scopolin on the osteoblast markers Alp, Runx2, Bglap, and Sp7 in the osteoblast cell line E1, qRT-PCR was performed. MC3T3-E1 cells were treated with 5 μM/mL of Scopolin for 3 days. Total RNA was extracted and mRNA expression of Alp, Runx2, Bglap, and Sp7 was measured by qRT-PCR. The expression of Alp, Runx2, Bglap, and Sp7 significantly increased in the Scopolin-treated group (Figure 6). The results suggest that Scopolin induces the differentiation of osteoblasts by upregulating the mRNA expression of Alp, Runx2,
Figure 6. The mRNA expression of osteoblastic markers Alp, Runx2, Bglap, and Sp7 in MC3T3-E1 treated with Scopolin. MC3T3-E1 cells were incubated with 5 μM of Scopolin for 3 days. The mRNA expression of osteoblastic markers Alp,
Runx2, Bglap, and Sp7 was evaluated by reverse-transcription PCR (RT-PCR) using
targeted gene-specific primers and then normalized to Gapdh mRNA expression. 0.0 0.5 1.0 1.5 2.0 2.5 3.0 * * Alp Rel a tiv e Alp e x pre ss ion A Sp7 0.0 0.5 1.0 1.5 2.0 2.5 3.0 * * R el ativ e Sp7 ex pr es sion D Runx2 0.0 0.5 1.0 1.5 * * Rel a tiv e Ru nx 2 e x pre s s ion B Bglap 0 4 8 12 * * Rel a tiv e Bg la p e x pre s s ion C
- 22 -
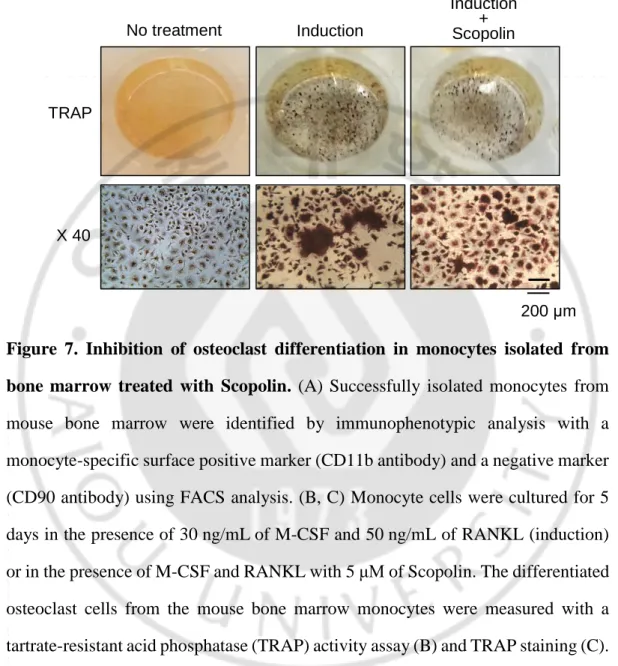
C. Scopolin decreased differentiation of osteoclast cells isolated from bone marrow.
Bone continuously undergoes formation and resorption. However, an imbalance of bone formation and resorption causes osteoporosis. To investigate the effects of Scopolin on the differentiation of osteoclasts, primary cultured monocytes were isolated from the bone marrow of 7-week-old mice. The isolated monocytes were confirmed by immunophenotypic analysis with monocyte-specific surface markers (CD11b antibody) using FACS analysis (Figure 7A). Monocytes were cultured in the presence of macrophage colony-stimulating factor-1 (M-CSF) and RANKL to induce differentiation to osteoclasts. Moreover, monocytes were treated with three different concentrations (1, 5, and 10 μM) of Scopolin for either 3 days or 5 days; differentiation was significantly inhibited by 5 μM Scopolin at 5 days. Monocytes were evaluated for TRAP activity, which decreased in a Scopolin dose-dependent manner. (Figure 7B). TRAP-positive colonies decreased in the cells treated with 5 μM Scopolin (Figure 7C). As a result, Scopolin decreased the differentiation of osteoclasts in isolated monocytes from mouse bone marrow.
A
CD11b- CD11b+ 100 101 102 103 104 PE-A 50 100 C ell co un t (% ) 0B
0 50 100 Control Sco1 Sco5 Sco10*
*
*
*
Day 3 5 R elat iv e T R A P ac ti v it y- 24 -
Figure 7. Inhibition of osteoclast differentiation in monocytes isolated from bone marrow treated with Scopolin. (A) Successfully isolated monocytes from mouse bone marrow were identified by immunophenotypic analysis with a monocyte-specific surface positive marker (CD11b antibody) and a negative marker (CD90 antibody) using FACS analysis. (B, C) Monocyte cells were cultured for 5 days in the presence of 30 ng/mL of M-CSF and 50 ng/mL of RANKL (induction) or in the presence of M-CSF and RANKL with 5 μM of Scopolin. The differentiated osteoclast cells from the mouse bone marrow monocytes were measured with a tartrate-resistant acid phosphatase (TRAP) activity assay (B) and TRAP staining (C). Control: Monocyte cells cultured with M-CSF and RANKL. *: p < 0.05 vs. control.
C
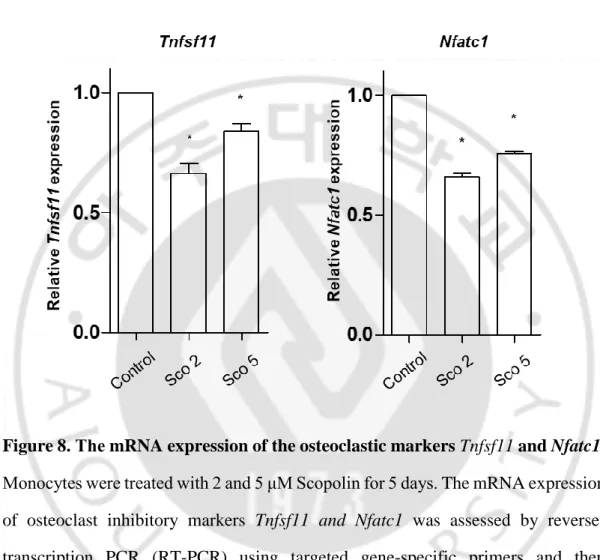
No treatment Induction Induction + Scopolin TRAP X 40 200 μmD. Scopolin suppressed the mRNA expression of the osteoclastic markers Tnfsf11 and Nfatc1.
To further confirm the inhibitory effects of Scopolin on osteoclast differentiation, osteoclast markers (Tnfsf11 and Nfatc1) were measured. Isolated monocytes derived from mouse bone marrow were treated with 2 and 5 μM Scopolin. After 5 days of incubation, total RNA was extracted and amplified by RT-PCR, and the expression of osteoclast inhibitory markers such as Tnfsf11 and Nfatc1 was assessed. Scopolin treatment decreased the mRNA expression of Tnfsf11 and Nfatc1 as compared to the control. These results suggest that Scopolin suppressed the differentiation of osteoclasts by downregulating the mRNA expression of Tnfsf11 and Nfatc1 (Figure 8).
- 26 -
Figure 8. The mRNA expression of the osteoclastic markers Tnfsf11 and Nfatc1. Monocytes were treated with 2 and 5 μM Scopolin for 5 days. The mRNA expression of osteoclast inhibitory markers Tnfsf11 and Nfatc1 was assessed by reverse-transcription PCR (RT-PCR) using targeted gene-specific primers and then normalized to Gapdh mRNA expression. *: p < 0.05 vs. control.
E. Scopolin increased osteoblast differentiation and decreased osteoclast differentiation in a co-culture of isolated monocytes and the MC3T3-E1 osteoblastic cell line.
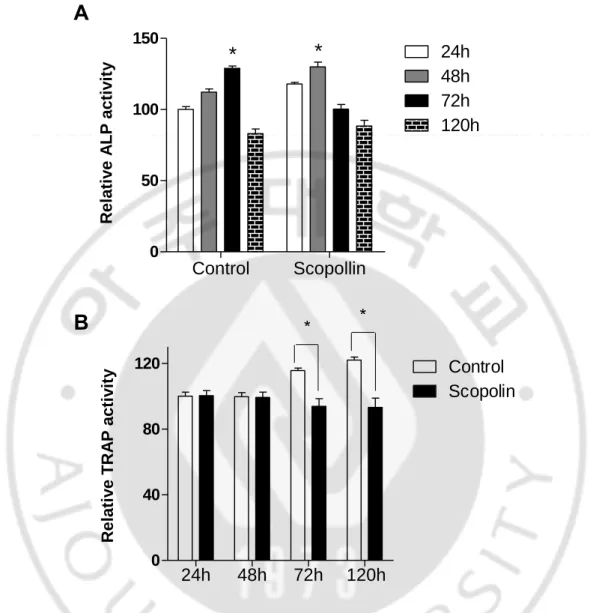
Since bone continuously undergoes formation by osteoblasts and resorption by osteoclasts, I tested the anti-osteoporotic effects of Scopolin in a co-culture of osteoblasts and osteoclasts. Preosteoblastic MC3T3-E1 and preosteoclastic monocytes isolated from mouse bone marrow were co-cultured and treated with Scopolin for 24, 48, 72, and 120 h. ALP activity in Scopolin-treated cells increased at 48 h compared to that in the control, which reached its peak at 72 h (Figure 9A). In the co-culture system, osteoclast differentiation increased in a time-dependent manner. However, the ALP activity in monocytes treated with 5 μM Scopolin decreased as compared to the control (Figure 9B). Therefore, Scopolin promoted osteoblast differentiation and reduced bone loss in the co-culture system.
- 28 -
Figure 9. Differentiation of osteoblasts and osteoclasts in a co-culture of isolated monocytes and the osteoblastic MC3T3-E1 cell line. Isolated monocyte cells (4 × 104 cells/well) and MC3T3-E1 (2 × 104 cells/well) were co-cultured for 24, 48, 72, and 120 h, and (A) the alkaline phosphatase (ALP) and (B) tartrate-resistant acid phosphatase (TRAP) activity were analyzed to determine osteoblast and osteoclast differentiation, respectively. *: p < 0.05, **: p < 0.005. Control Scopollin 0 50 100 150 24h 48h 72h 120h
*
*
A
R elat iv e A LP ac ti v it y 24h 48h 72h 120h 0 40 80 120 Control Scopolin * *B
R elat iv e T R A P ac ti v it yF. Scopolin prevented OVX-induced bone loss in mice.
Finally, I evaluated the effects of Scopolin on bone formation in an OVX mouse model. Thirty of the 8-week-old female ddY mice had either an ovariectomy or sham surgery. Mice were divided into five groups of six mice each: (1) Sham, (2) OVX control, (3) OVX with 10 mg/kg/day of SrCl2 positive control, (4) OVX with 20 mg/kg/day of Scopolin, and (5) OVX with 40 mg/kg/day of Scopolin. The mice were housed for 12 weeks and their total body weight, BMD, and BMC were measured using a PIXI-mus bone densitometer.
Transverse micro-CT images of the right femur were scanned using micro-CT, and BV/TV and Tb.Th, Tb.N, and Tb.Sp were also analyzed. Significant trabecular bone loss was found in the OVX group as compared to the sham group, and the positive control SrCl2 and Scopolin treatment prevented OVX-induced bone loss. Treatment with the positive control SrCl2 and Scopolin 20 and 40 mg/kg/day for 12 weeks inhibited OVX-induced BMD loss in the right femur bone, and trabecular bone structural properties such as BV/TV, Tb.Th, Tb.N and Tb.Sp improved as compared to the OVX control group (Figure 10A–C).
In addition, for a detailed analysis of bone loss, the 2D and 3D micro-CT images of right femur were evaluated. Significant trabecular bone loss was found in the OVX group as compared to the sham group. However, treatment with the positive control SrCl2 and Scopolin prevented OVX-induced bone loss (Figure 10B). In planar
- 30 -
technetium Tc-99m HDP presents as a function of skeletal blood flow and osteogenic activity. For comparison, paired mice were scanned in the same panel. A higher level of radioactivity was detected in the sham and Scopolin-treated groups than in the OVX group. Similar radiographic patterns were found in the SrCl2 and Scopolin-treated groups (Figure 10D). High radioactive signals detected in the bladder are the radioisotopes that are not interacted with bone and thus gathered in the bladder. These results suggested that Scopolin prevents OVX-induced bone loss in an osteoporotic mouse model.
To further investigate the effects of Scopolin on serum levels of bone metabolic makers, blood samples were collected at the last day of treatment, and bone metabolic makers (OPG and RANKL) were measured using ELISA. The positive control SrCl2 increased serum levels of the osteoblast marker OPG and decreased those of the osteoclast marker RANKL as compared to OVX mice. Similarly, treatment with Scopolin significantly increased serum OPG levels and decreased serum levels of RANKL (Figure 10E). The ratio of OPG/RANKL significantly increased following treatment with Scopolin (Figure 10E), indicating inhibition of osteoclastogenesis. These results and the in vivo study data suggest that Scopolin prevents trabecular bone loss and osteoclastogenesis in OVX mice.
OVX Sham Sco20 Sco40 SrCl2 * * B MD ( g /cm 2 ) 0.07 0.08 0.10 0.11 0 6 12 week 0.06 0.09
A
B
Sham OVX SrCl2 Sco20 Sco40- 32 -
C
B V /T V ( % ) 20 0 40 60 80 * * * * T b.T h ( m m) * * * * 0.010 0.015 0.020 0.025 T b.Sp ( m m) * * * * 0.00 0.05 0.10 0.20 0.15 T b.N ( 1/ m m) 0 2 3 4 * * * * 1D
400 450 500 550 600 650 S erum O P G ( pg /m l) * * * * 0 20 40 60 80 100 S erum R A N K L ( pg /m l) * * * * 0 5 10 15 20 25 S erum O P G /R A N K L ( % ) * * * *
E
- 34 -
Figure 10. Inhibition of OVX-induced bone loss in mice treated with Scopolin. The OVX mice were administered either SrCl2 (10 mg/kg/day) or Scopolin (20 or 40 mg/kg/day) for 12 weeks. Sham: sham-operated, OVX: non-Scopolin-administered mice. (A) Right femur bone mineral density (BMD) was measured using a PIXI-mus bone densitometer at 0, 6, and 12 weeks. (B) Transverse micro-CT images of the right femur were scanned using a microcomputed tomography (CT) and (C) bone volume (BV/TV) and trabecular thickness (Tb.Th), number (Tb.N) and spacing (Tb.Sp) were analyzed at 12 weeks of treatment. (D) Technetium Tc-99m HDP bone scan was assessed using an Inveon SPECT scanner. (E) The blood samples were collected after 12 weeks of administration. Osteoprotegerin (OPG) and RANKL were measured using ELISA, and the ratio of OPG/RANKL was analyzed. *:p <
IV. DISCUSSION
Bone ingeminates bone remodeling by bone formation (osteoblasts) and bone resorption (osteoclasts) (Raggatt and Partridge, 2010). Osteoporosis is a bone disease characterized by an imbalance in bone remodeling, which increases the risk of fractures because of low BMD (Bover et al., 2017). Recently, the incidence of osteoporosis owing to the process of aging has increased (Drake et al., 2015). Various pharmacological medications for the treatment of osteoporosis have been discovered. Bisphosphonates can treat osteoporosis by inhibiting osteoclasts without new bone formation, but it is not suitable for long-term use because of its side effects (Alwahhabi and Alsuwaine, 2017). However, plants have been used as alternative treatments for diverse diseases for a long time and have few side effects (Remport and Blazovics, 2017). Many studies have suggested that various herbal extracts can be used as osteoporosis drugs with fewer side effects (Kim et al., 2016; Jin et al., 2017).
Our previous study demonstrated the beneficial effects of LRC extract on osteoporosis both in vitro and in vivo (Park et al., 2014). In this study, I isolated a specific bioactive compound, Scopolin, from LRC extract for its bone formation-enhancing effects. Scopolin is an important secondary metabolite synthesized in plants as a defense mechanism against various environmental stresses (Siwinska et
- 36 -
osteoblast differentiation promotes bone formation (Ducy et al., 2000). Osteoblasts mineralize the bone matrix and lead to calcium-induced mineralization of bone (Gough et al., 2004). In this study, Scopolin significantly increased ALP activity and mineralized nodule formation in MC3T3-E1 cells.
A previous study suggested the clinical utility of biochemical markers of bone remodeling (Watts, 1999). ALP increases during active bone formation with the induction of osteoblast activity, suggesting that ALP plays a crucial role in the mineralization of newly formed bone (Watts, 1999). Runx2 is a key transcription factor related to osteoblast differentiation (Gilbert et al., 2002). Osteocalcin (OCN), which is secreted by osteoblasts, is an important factor in the regulation of bone metabolism and in the implication of bone mineralization and calcium ion homeostasis (Lee et al., 2007). Sp7 is a downstream target of Runx2 and is a transcription factor of osteoblast differentiation (Rashid et al., 2014). Osteoblast markers were upregulated in MC3T3-E1 cells treated with Scopolin. As a result, Scopolin promoted osteoblast differentiation and mineralization by upregulating osteoblast markers in new bone formation.
The imbalanced regulation of the bone-remodeling process promotes abnormal metabolic bone diseases, especially osteoporosis (Feng and McDonald, 2011). Osteoporosis patients experience excessive bone resorption rather than bone formation, leading to increased risk of bone fragility and susceptibility to fractures (Yoshida et al., 2002). TRAP is a histochemical marker of osteoclasts (Salamanna et
al., 2016). Tumor necrosis factor (Ligand) superfamily, member 11 (TNFSF11) encodes RANKL which mediates osteoclastogenesis, and Nuclear Factor of Activated T-Cells 1 (NFATC1) regulates osteoclast-related genes for osteoclast differentiation and function (Nakagawa et al., 2015).
A previous study demonstrated that Scopolin promotes inhibitory effects on osteoclast differentiation in pro-osteoclastic RAW 264.7 cells via the suppression of RNAKL-induced osteoclast differentiation (Lee et al., 2013). Accordingly, this study revealed that Scopolin decreased osteoclast differentiation in primary cultured monocytes from mouse bone marrow. Furthermore, osteoclast markers (Rankl and
Nfatc1) were significantly reduced by treatment with Scopolin in primary cultured
monocytes, indicating inhibition of osteoclast differentiation. These results suggested that Scopolin prevented RANKL-induced osteoclast differentiation by downregulating osteoclast markers (Tnfsf11 and Nfatc1) in primary cultured monocytes. Similar results were found in a co-culture system of monocyte and MC3T3-E1; Scopolin treatment enhanced osteoblast differentiation and inhibited osteoclast differentiation. In summary, Scopolin may have enhancing effects on bone formation in osteoblast cells and suppressive effects on resorbent bone loss in osteoclast cells.
Based on the in vitro study, I further investigated the anti-osteoporotic effects of Scopolin treatment on BMD loss in the osteoporotic mouse model. The OVX mouse model is well known as a model for human postmenopausal women, and is
- 38 -
characterized by significant low bone mass and BMD (Wu et al., 2013). I inducted SrCl2 as a positive control for the anti-osteoporotic agent, which has previously shown protective effects of SrCl2 on bone loss (Pemmer et al., 2011). As expected, OVX mice had significantly reduced BMD in the right femur bone and impaired trabecular bone structural properties. However, Scopolin prevented a decrease in BMD in OVX mice, and image analysis of 2D and 3D micro-CT of the right femur further confirmed protective BMD loss in Scopolin-treated mice. In addition, impaired bone structural parameters such as BV/TV, Tb.Th, Tb.N and Tb.Sp were improved by Scopolin treatment.
Next, I tested the effects of Scopolin on bone metabolic markers in blood samples. Many studies have demonstrated the OPG/RANKL signaling system as a key mediator of mechanical strain on bone metabolism (Li et al., 2015). RANKL is a key mediator of bone resorption, and OPG is well known as an osteoclastogenesis inhibitory factor; it is a decoy receptor for RANKL and prevents RANK-mediated nuclear kappa B (NF-κB) activation (Li et al., 2015). OVX rats have been associated with increased serum RANKL (Jabbar et al., 2011). However, the inhibition of RANKL by OPG decreases the bone loss in estrogen ablation animal models through the suppression of bone resorption. In this study, Scopolin prevented the OVX-induced elevation of RANKL and decreased OPG serum levels. Notably, Scopolin treatment prevented unbalanced serum levels of OPG and RNAKL in OVX mice, subsequently promoting bone formation with preserved trabecular architecture. Taken together with the in vivo results, Scopolin prevented osteoporotic bone loss in
OVX mice.
The biological activities of LRC improve insulin resistance and lipid metabolism in a Type 2 diabetes rat known to be associated with osteoporosis (Inzerillo and Epstein, 2004). Scopolin also has effects on hepatic steatosis induced by a high-fat diet, which is used in the diabetes mellitus mouse model; it improves hepatic steatosis involving SIRT1-mediated signaling cascades (Yoo et al., 2017). SIRT1 upregulates osteoblast transcription factor Runx2 in osteoporosis (Shakibaei et al., 2012; Zainabadi et al., 2017). SRT2183 and SRT3025, SIRT1 activators, inhibit RANKL-induced osteoclastogenesis in monocytes isolated from bone marrow (Gurt et al., 2015). In addition, SIRT1 induces RANK inhibition and then suppresses RANKL-induced osteoclast differentiation (Park et al., 2015). From these previous studies, it is possible that Scopolin may control osteoblast and osteoclast differentiation through SIRT1 signaling. Further studies need to investigate this hypothesis.
In summary, Scopolin increased osteoblast differentiation by upregulating osteoblast markers. However, osteoclast differentiation was suppressed by Scopolin treatment via downregulation of osteoclast markers. Scopolin consistently inhibited osteoporotic bone loss in the OVX mouse model. This study suggests that Scopolin may be a potential therapeutic agent for the treatment and prevention of bone loss.
- 40 -
V. CONCLUSION
Scopolin, a bioactive compound isolated from LRC, was revealed to have bone formation-enhancing and bone resorption-inhibiting effects in in vitro and in vivo experiments. Scopolin treatment enhanced osteoblast differentiation through the upregulation of osteoblastic markers and suppressed osteoclast differentiation through the downregulation of osteoclast markers. In vivo experiments demonstrated that Scopolin promotes the inhibition of osteoporotic bone loss.
In conclusion, this study suggests that Scopolin may be a good therapeutic compound for the treatment and prevention of osteoporosis.
REFERENCES
1. Alwahhabi BK, Alsuwaine BA: Long-term use of bisphosphonates in osteoporosis. Saudi Med J 38: 604-608, 2017
2. Bover J, Bailone L, Lopez-Baez V, Benito S, Ciceri P, Galassi A, Cozzolino M: Osteoporosis, bone mineral density and CKD-MBD: treatment considerations. J Nephrol 30: 677-687, 2017
3. Ding J, Ghali O, Lencel P, Broux O, Chauveau C, Devedjian JC, Hardouin P, Magne D: TNF-alpha and IL-1beta inhibit RUNX2 and collagen expression but increase alkaline phosphatase activity and mineralization in human mesenchymal stem cells. Life Sci 84: 499-504, 2009
4. Drake MT, Clarke BL, Lewiecki EM: The Pathophysiology and Treatment of Osteoporosis. Clin Ther 37: 1837-1850, 2015
5. Ducy P, Schinke T, Karsenty G: The osteoblast: a sophisticated fibroblast under central surveillance. Science 289: 1501-1504, 2000
6. Ensrud KE, Crandall CJ: Osteoporosis. Ann Intern Med 167: ITC17-ITC32, 2017
7. Feng X, McDonald JM: Disorders of bone remodeling. Annu Rev Pathol 6: 121-145, 2011
8. Florencio-Silva R, Sasso GR, Sasso-Cerri E, Simoes MJ, Cerri PS: Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells.
- 42 -
Biomed Res Int 2015: 421746, 2015
9. Fu YX, Gu JH, Zhang YR, Tong XS, Zhao HY, Yuan Y, Liu XZ, Bian JC, Liu ZP: Osteoprotegerin influences the bone resorption activity of osteoclasts. Int
J Mol Med 31: 1411-1417, 2013
10. Gilbert L, He X, Farmer P, Rubin J, Drissi H, van Wijnen AJ, Lian JB, Stein GS, Nanes MS: Expression of the osteoblast differentiation factor RUNX2 (Cbfa1/AML3/Pebp2alpha A) is inhibited by tumor necrosis factor-alpha. J
Biol Chem 277: 2695-2701, 2002
11. Gough JE, Jones JR, Hench LL: Nodule formation and mineralisation of human primary osteoblasts cultured on a porous bioactive glass scaffold.
Biomaterials 25: 2039-2046, 2004
12. Gurt I, Artsi H, Cohen-Kfir E, Hamdani G, Ben-Shalom G, Feinstein B, El-Haj M, Dresner-Pollak R: The Sirt1 Activators SRT2183 and SRT3025 Inhibit RANKL-Induced Osteoclastogenesis in Bone Marrow-Derived Macrophages and Down-Regulate Sirt3 in Sirt1 Null Cells. PLoS One 10: e0134391, 2015
13. Inzerillo AM, Epstein S: Osteoporosis and diabetes mellitus. Rev Endocr
Metab Disord 5: 261-268, 2004
14. Jabbar S, Drury J, Fordham JN, Datta HK, Francis RM, Tuck SP: Osteoprotegerin, RANKL and bone turnover in postmenopausal osteoporosis.
J Clin Pathol 64: 354-357, 2011
Herbal Medicine for Osteoporosis: A Meta-analysis of Randomized Controlled Trials. J Clin Densitom 20: 516-525, 2017
16. Johnell O, Kanis J: Epidemiology of osteoporotic fractures. Osteoporos Int 16 Suppl 2: S3-7, 2005
17. Katsimbri P: The biology of normal bone remodelling. Eur J Cancer Care
(Engl) 26, 2017
18. Khosla S: Minireview: the OPG/RANKL/RANK system. Endocrinology 142: 5050-5055, 2001
19. Kim JH, Kim EY, Lee B, Min JH, Song DU, Lim JM, Eom JW, Yeom M, Jung HS, Sohn Y: The effects of Lycii Radicis Cortex on RANKL-induced osteoclast differentiation and activation in RAW 264.7 cells. Int J Mol Med 37: 649-658, 2016
20. Lacombe J, Cairns BJ, Green J, Reeves GK, Beral V, Armstrong ME, Million Women Study c: The Effects of Age, Adiposity, and Physical Activity on the Risk of Seven Site-Specific Fractures in Postmenopausal Women. J Bone
Miner Res 31: 1559-1568, 2016
21. Lee NK, Sowa H, Hinoi E, Ferron M, Ahn JD, Confavreux C, Dacquin R, Mee PJ, McKee MD, Jung DY, Zhang Z, Kim JK, Mauvais-Jarvis F, Ducy P, Karsenty G: Endocrine regulation of energy metabolism by the skeleton. Cell 130: 456-469, 2007
- 44 -
macrophage RAW 264.7 cells by scavenging reactive oxygen species. J Nat
Prod 76: 615-620, 2013
23. Li CW, Liang B, Shi XL, Wang H: Opg/Rankl mRNA dynamic expression in the bone tissue of ovariectomized rats with osteoporosis. Genet Mol Res 14: 9215-9224, 2015
24. Liu Y, Sun M, Yao H, Liu Y, Gao R: Herbal Medicine for the Treatment of Obesity: An Overview of Scientific Evidence from 2007 to 2017. Evid Based
Complement Alternat Med 2017: 8943059, 2017
25. Mohammad KS, Chen CG, Balooch G, Stebbins E, McKenna CR, Davis H, Niewolna M, Peng XH, Nguyen DH, Ionova-Martin SS, Bracey JW, Hogue WR, Wong DH, Ritchie RO, Suva LJ, Derynck R, Guise TA, Alliston T: Pharmacologic inhibition of the TGF-beta type I receptor kinase has anabolic and anti-catabolic effects on bone. PLoS One 4: e5275, 2009
26. Nakagawa T, Ohta K, Kubozono K, Ishida Y, Naruse T, Takechi M, Kamata N: Zoledronate inhibits receptor activator of nuclear factor kappa-B ligand-induced osteoclast differentiation via suppression of expression of nuclear factor of activated T-cell c1 and carbonic anhydrase 2. Arch Oral Biol 60: 557-565, 2015
27. Pacifici R: Bone Remodeling and the Microbiome. Cold Spring Harb
Perspect Med, 2017
28. Park E, Jin HS, Cho DY, Kim J, Kim MC, Choi CW, Jin Y, Lee JW, Park JH, Chung YS, Huh D, Jeong SY: The effect of Lycii Radicis Cortex extract on
bone formation in vitro and in vivo. Molecules 19: 19594-19609, 2014 29. Park SY, Lee SW, Kim HY, Lee SY, Lee WS, Hong KW, Kim CD:
Suppression of RANKL-induced osteoclast differentiation by cilostazol via SIRT1-induced RANK inhibition. Biochim Biophys Acta 1852: 2137-2144, 2015
30. Pemmer B, Hofstaetter JG, Meirer F, Smolek S, Wobrauschek P, Simon R, Fuchs RK, Allen MR, Condon KW, Reinwald S, Phipps RJ, Burr DB, Paschalis EP, Klaushofer K, Streli C, Roschger P: Increased strontium uptake in trabecular bone of ovariectomized calcium-deficient rats treated with strontium ranelate or strontium chloride. J Synchrotron Radiat 18: 835-841, 2011
31. Potterat O: Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med 76: 7-19, 2010
32. Raggatt LJ, Partridge NC: Cellular and molecular mechanisms of bone remodeling. J Biol Chem 285: 25103-25108, 2010
33. Rashid H, Ma C, Chen H, Wang H, Hassan MQ, Sinha K, de Crombrugghe B, Javed A: Sp7 and Runx2 molecular complex synergistically regulate expression of target genes. Connect Tissue Res 55 Suppl 1: 83-87, 2014 34. Remport J, Blazovics A: [Phytoestrogens in the treatment of menopause].
- 46 -
(eds. De Groot LJ, Chrousos G, Dungan K, Feingold KR, Grossman A, Hershman JM, Koch C, Korbonits M, McLachlan R, New M, Purnell J, Rebar R, Singer F, Vinik A) South Dartmouth (MA), 2000
36. Salamanna F, Pagani S, Maglio M, Borsari V, Giavaresi G, Martelli AM, Buontempo F, Fini M: Estrogen-deficient osteoporosis enhances the recruitment and activity of osteoclasts by breast cancer cells. Histol
Histopathol 31: 83-93, 2016
37. Shakibaei M, Shayan P, Busch F, Aldinger C, Buhrmann C, Lueders C, Mobasheri A: Resveratrol mediated modulation of Sirt-1/Runx2 promotes osteogenic differentiation of mesenchymal stem cells: potential role of Runx2 deacetylation. PLoS One 7: e35712, 2012
38. Siwinska J, Kadzinski L, Banasiuk R, Gwizdek-Wisniewska A, Olry A, Banecki B, Lojkowska E, Ihnatowicz A: Identification of QTLs affecting scopolin and scopoletin biosynthesis in Arabidopsis thaliana. BMC Plant
Biol 14: 280, 2014
39. Sun L, Peng Y, Sharrow AC, Iqbal J, Zhang Z, Papachristou DJ, Zaidi S, Zhu LL, Yaroslavskiy BB, Zhou H, Zallone A, Sairam MR, Kumar TR, Bo W, Braun J, Cardoso-Landa L, Schaffler MB, Moonga BS, Blair HC, Zaidi M: FSH directly regulates bone mass. Cell 125: 247-260, 2006
40. Wang Y, Zheng Q, Guo X, Wu Y, Hao J: Influence of exogenous TGFbeta1 on the expression of smad2 and smad3 in rat bone marrow-derived mesenchymal stem cells. J Huazhong Univ Sci Technolog Med Sci 25: 68-71,
2005
41. Watts NB: Clinical utility of biochemical markers of bone remodeling. Clin
Chem 45: 1359-1368, 1999
42. Weilbaecher KN, Guise TA, McCauley LK: Cancer to bone: a fatal attraction.
Nat Rev Cancer 11: 411-425, 2011
43. Wu K, Lin TH, Liou HC, Lu DH, Chen YR, Fu WM, Yang RS: Dextromethorphan inhibits osteoclast differentiation by suppressing RANKL-induced nuclear factor-kappaB activation. Osteoporos Int 24: 2201-2214, 2013
44. Ye Z, Huang Q, Ni HX, Wang D: Cortex Lycii Radicis extracts improve insulin resistance and lipid metabolism in obese-diabetic rats. Phytother Res 22: 1665-1670, 2008
45. Yoo A, Narayan VP, Hong EY, Whang WK, Park T: Scopolin ameliorates high-fat diet induced hepatic steatosis in mice: potential involvement of SIRT1-mediated signaling cascades in the liver. Sci Rep 7: 2251, 2017 46. Yoshida K, Oida H, Kobayashi T, Maruyama T, Tanaka M, Katayama T,
Yamaguchi K, Segi E, Tsuboyama T, Matsushita M, Ito K, Ito Y, Sugimoto Y, Ushikubi F, Ohuchida S, Kondo K, Nakamura T, Narumiya S: Stimulation of bone formation and prevention of bone loss by prostaglandin E EP4 receptor activation. Proc Natl Acad Sci U S A 99: 4580-4585, 2002
- 48 -
48. Zeng QM, Liu DC, Zhang XC, Wu JP, Ding L, Meng DH, Zhang J: Estrogen deficiency inducing shifted cytokines profile in bone marrow stromal cells inhibits Treg cells function in OVX mice. Cell Mol Biol (Noisy-le-grand) 61: 64-68, 2015
49. Zhang S, Wang X, Li G, Chong Y, Zhang J, Guo X, Li B, Bi Z: Osteoclast regulation of osteoblasts via RANKRANKL reverse signal transduction in vitro. Mol Med Rep 16: 3994-4000, 2017
-국문요약-
Scopolin의 조골세포 및 파골세포 분화 조절 기전 규명
아주대학교 대학원 의학과 김 지 은 (지도교수: 정 선 용) 골다공증은 뼈의 밀도가 낮아져 그 강도가 약해지면서 골절이 증가되 는 질환을 말한다. 최근 고령화의 진전에 따른 골다공증 환자 수가 급격 하게 증가하고 있다. 세계적으로 가장 많이 사용되는 골다공증 치료제로 비스포스포네이트 같은 약제가 있으나, 부작용이 많아 장기 투여가 어려 우며, 새로운 뼈 생성에 의한 골밀도 증가에는 큰 역할을 하지 않는다는 단점이 있다. 따라서 장기 복용에도 부작용이 적고 골밀도 개선에 효과 가 있는 천연물 유래물질의 치료제의 개발이 필요하다. 본 연구에서는 골다공증에 효과가 있다고 밝혀진 천연물인 지골피 (Lycii Radicis Cortex)에서 추출한 물질 중 하나인 Scopolin의 골다공 증 개선 효능에 대한 in vitro 및 in vivo 연구를 수행하였다. 먼저 Scopolin의 조골세포의 골형성(bone formation) 효능에 대한 실험을 시 행하였다. 조골모세포인 MC3T3-E1에 Scopolin을 처리하여 조골세포 분화능 지표인 alkaline phosphatase(ALP)의 활성을 분석한 결과- 50 -
며 조골세포 분화 마커인 Alp, Runx2, Bglap, Sp7의 mRNA 발현도 유 의하게 증가되었다. 또한, Alizarin red S 염색 결과에서 뼈형성 과정에 나타나는 조골세포의 석회화 결절형성(mineralized nodule formation)이 증가되었다.
다음은, Scopolin의 파골세포 골흡수(bone resorption)에 대한 영향을 조사하였다. 마우스 골수로부터 분리·배양한 초대배양 단핵세포 (primary-cultured monocyte)에 Scopolin을 처리하였을 때 tartrate-resistant acid phosphatase(TRAP) 활성과 파골세포 분화 마커인 Tnfsf11과 Nfatc1의 mRNA의 발현량이 유의하게 감소하였다.
신체내에서는 조골세포와 파골세포가 공존하며 뼈 리모델링(bone remodeling)이 조절된다. 이러한 신체내 조건에 가까운 환경에서의 Scopolin 효과를 분석하였다. 조골세포 전구체인 MC3T3-E1과 파골세 포 전구체인 monocyte의 두가지 세포를 함께 공배양(co-culture)하여 Scopolin을 처리한 결과, ALP 증가와 TRAP 감소가 나타났다.
끝으로, Scopolin의 in vivo 효과를 조사하였다. 골다공증동물 모델은 난 소절제(ovariectomized, OVX) 마우스를 제작하여 실험에 사용하였다. Scopolin을 12주간 구강주입한 OVX 마우스는 대조군과 비교시에 골밀 도(bone mineral density), 혈액에서의 OPG/RANKL 비율, micro-CT 와 뼈 스캔(SPECT scan)에서의 뼈의 미세구조 및 뼈 굵기 등의 전반적 인 골다공증 평가 자료에서 유의한 개선 효과가 나타났다. 결론적으로, 지골피 에탄올 추출물의 효능성분인 Scopolin은 조골세포 의 분화 증진을 통한 뼈 형성 촉진과 파골세포의 분화억제를 통한 뼈 흡 수 억제에 효능이 있음을 증명하였다. 또한, 동물실험에서 Scopolin이 난소절제에 의해 나타나는 골다공증의 개선 또는 치료에 좋은 효과가 있
음을 밝혔다. 본 논문에서 밝혀진 Scopolin에 대한 연구결과가 향후 Scopolin를 이용한 골다공증 치료제 개발에 기초자료로 활용될 수 있기 를 기대한다. __________________________________________________________________ 핵심어: 천연물, 지골피, Scopolin, 골다공증, 조골세포, 파골세포, 뼈 형성, 뼈 흡수, 난소절제 마우스, 골밀도, 뼈 미세구조


![Figure 4. Chemical structure of the isolated Scopolin. 6-methoxy-7-[(2S, 3R, 4S, 5S, 6R)-3, 4, 5-trihydroxy-6-(hydroxymethyl) oxan-2-yl] oxychromen-2-one](https://thumb-ap.123doks.com/thumbv2/123dokinfo/4688434.3610/19.773.84.687.256.854/figure-chemical-structure-isolated-scopolin-trihydroxy-hydroxymethyl-oxychromen.webp)