저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Master's Thesis in Medicine
Transcriptomes of CD27-CD28- and
CD27+CD28+ CD8+ T subsets
disclose differentially expressed
novel genes and signaling pathways
in Behçet's disease
Graduate School of Ajou University
Department of Medical Sciences
Transcriptomes of CD27-CD28- and
CD27+CD28+ CD8+ T subsets
disclose differentially expressed
novel genes and signaling pathways
in Behçet's disease
Eun-So Lee, M.D., Ph.D., Advisor
I submit this thesis as the
Master's thesis in medicine.
August, 2019
Graduate School of Ajou University
Department of Medical Sciences
The Master's thesis of So Min Kim in medicine is
hereby approved.
Thesis Defense Committee Chair
Eun-So LeeMember
Seonghyang SohnMember
Hee Young Kang
Graduate School of Ajou University
i -ABSTRACT-
Transcriptomes of CD27-CD28- and CD27+CD28+ CD8+ T subsets disclose
differentially expressed novel genes and signaling pathways in Behçet's disease
Background and objectives: Behçet’s disease (BD) is a chronic inflammatory disease characterized
by recurrent mucocutaneous ulceration and other complications such as blindness and large vessel inflammation. Immunosenescence, aging of immune system, is related to increased susceptibility to infectious diseases, vaccine failure, and chronic low grade systemic inflammation. Our previous study showed increased frequency of senescent CD8+ T cells in the peripheral blood of patients with BD. In this study, to find the global genet-expression characteristics of senescent CD8+ T cells in relation with BD, we examined the transcriptome of CD8+ T lymphocyte subsets (CD27-CD28- senescent and CD27+CD28+ nonsenescent) from BD patients and healthy control (HC) subjects.
Material and methods: Peripheral blood mononuclear cells (PBMCs) were collected from BD
patients (n=18) and HCs (n=18). CD8+ T cells were isolated through CD8 microbeads, and those were labelled with conjugated monoclonal antibodies as follows: FITC anti-CD8, allophycocyanin (APC)-H7 anti-CD27 and APC anti-CD28. Using fluorescence-activated cell sorting (FACS), senescent CD8+ T cells (CD8+CD27-CD28-) and non-senescent CD8+ T cells (CD8+CD27+CD28+) were sorted. After sorting, each group of cells were pooled together and cultured in medium (RPMI 1640). Cells were stimulated with anti-CD3 (500ng/ml, clone OKT3) for 72 hours. Total RNA was extracted from anti-CD3-stimulated cells with the RNA isolation kit. Transcriptome analysis was performed. We analyzed the differentially expressed genes from the four different groups (BD patients vs. controls, senescent CD8+ T cells (CD8+ CD27- CD28-) vs. non-senescent CD8+ T cells (CD8+ CD27+ CD28+)). Differentially expressed genes were submitted to Ingenuity Pathway Analysis (IPA) for functional evaluation and identification of significant biological pathways. For the validation of genes that show large fold change value in transcriptome sequencing, additional five BD patients and five HCs were enrolled to collect RNA and perform the real-time PCR on eight genes.
Results: Differentially expressed 1103 genes were identified in BD CD27-CD28- subsets compared to
HC, while 652 genes were differentially expressed in BD CD27+CD28+ subsets compared to HC. As a result of the real-time PCR, COL5A1, ARHGEF10 showed the same tendency with the transcriptome analysis in the BD CD27-CD28- subsets compared to HCs, among which the statistical significance was shown in COL5A1. Meanwhile, TRPV3, ARHGEF10, UBE2F-SCLY, CD302, and SHANK1 showed the same tendency with the transcriptome analysis in the BD CD27+CD28+ subsets compared to HCs, and the statistical significance was found in TRPV3 and ARHGEF10. Of the significant canonical pathways identified in IPA, 11 pathways showed activity in the opposite direction between
CD27+CD28+ and CD27-CD28- subsets, while two pathways tended to be same. The most significant canonical pathway resulted from the IPA was the cAMP-mediated signaling.
Conclusion: This is the first study for transcriptome analysis of CD8+ T cells of BD patients
compared to HCs. This study was also the first to separate senescent CD8+ T cells and non-senescent CD8+ T cells to perform RNA sequencing, respectively. Transcriptome analysis found differentially expressed genes in patients with BD. And IPA suggested the pathways in which these differences in gene expression may be involved in the pathogenesis of BD. Using these differentially expressed genes may be useful for developing biomarkers in BD that can predict the treatment response as well as help diagnosing easily. Consequently, we hope that this genetic profiling can be used as a key for approaching the pathogenesis of BD.
Keywords: Behçet's disease, transcriptome analysis, RNA sequencing, CD8+ T cell, immunosenescence
iii
TABLE OF CONTENTS
ABSTRACT ... i
TABLE OF CONTENTS ... iii
LIST OF FIGURES ...v
LIST OF TABLES ... vi
INTRODUCTION ...1
MATERIAL AND METHODS ...3
A. Subjects ...3
B. Isolation of peripheral blood mononuclear cells (PBMCs) ...4
C. Sorting with cell surface marker flow cytometry ...4
D. Cell culture and stimulation ...6
E. RNA extraction ...6
F. RNA transcriptome analysis ...6
G. Identification of differentially expressed genes (DEGs) ...6
H. Ingenuity Pathway Analysis (IPA®) ...6
I. RNA preparation and real-time polymerase chain reaction (PCR) ...6
J. Statistical analysis ...7 RESULTS ...8 DISCUSSION ...17 CONCLUSION ...22 REFERENCES ...23 국문요약 ...27
LIST OF FIGURES
Figure 1. Dot plots of immunosenescent cells of one representative Behçet’s disease patient analyzed with cell surface flow cytometry ...5 Figure 2. Venn diagram of differentially expressed genes in senescent and non-senescent CD8+ T cells of Behçet’s disease patients compared to healthy controls ...10 Figure 3. Real-time PCR ...13 Figure 4. Significant canonical pathways in Ingenuity Pathway Analysis (IPA) ...15 Figure 5. cAMP-mediated signaling in Behçet’s disease patients vs. healthy controls (senescent cells) with prediction: One of the significant canonical pathways in IPA ...16
v
LIST OF TABLES
Table. 1. Baseline demographics of Behçet’s disease patients and healthy controls...3 Table. 2. Clinical characteristics of Behçet’s disease patients ...9 Table. 3. Top ten genes based on expression fold change order in CD8+CD27-CD28- T cells of Behçet’s disease patients compared to healthy controls ... 11 Table. 4. Top ten genes based on expression fold change order in CD8+CD27+CD28+ T cells of Behçet’s disease patients compared to healthy controls ...12 Table. 5. Top biological functions of differentially expressed genes between Behçet’s disease patients and healthy controls as presented by Ingenuity Pathway Analysis (IPA) ...14
INTRODUCTION
Behçet’s disease (BD) is a chronic systemic inflammatory disease characterized by recurrent oro-genital ulcers, and ocular, skin lesions. It is a multisystemic disease involving multiple joints, gastrointestinal system, cardiovascular system, and central nervous system (Sakane et al., 1999; Hatemi et al., 2008). Although the exact pathogenesis of BD is unclear, it is supposed to have relationship with dysfunction of immune system in genetically predisposed hosts, which has properties of both autoimmune disease and autoinflammatory disease (Takeuchi et al., 2015). Excessive response of T helper type 1 (Th1), increased proinflammatory cytokines including interleukin (IL) 2 and interferon gamma (IFNγ) (Ben Ahmed et al., 2004), expected role of human heat-shock protein (HSP) 60 as a candidate autoantigen (Direskeneli and Saruhan-Direskeneli, 2003) and association with IL-17 (Ekinci et al., 2010; Na et al., 2013) are features that show relevance to autoimmunity. While, no specific high-titer of auto-antibodies or antigen-specific T-cells, association with M694V MEFV mutation (Imirzalioglu et al., 2005), increase of C reactive protein (CRP) and erythrocyte sedimentation rate (ESR) in active phase of disease, and IL-1β dysregulation (Liang et al., 2013; Yuksel et al., 2014) are the features related to autoinflammation. The relationship between autoimmune features and autoinflammatory features needs to be further clarified.
Immunosenescence is changes in the function and structure of the immune system that occurs with aging process, which lead to elderly people’s vulnerability to infectious diseases, vaccine failure, and possibly autoimmunity and cancer (Pawelec, 1999; Targonski et al., 2007). Among them, T cell immmunosenescence is characterized by a decrease in the number of naïve T cells due to atrophy of thymic activity, uneven and less diverse repertoire of T cells with expansions of clonal populations, and loss of costimulatory molecules expression, such as CD28, CD27, and CD40L (Saurwein-Teissl et al., 2002; Goronzy et al., 2007; Weiskopf et al., 2009; Weng et al., 2009). In addition to the elderly, these senescent T cells have been observed to increase in frequency in other autoimmune diseases such as RA, SLE and multiple sclerosis (Markovic-Plese et al., 2001; Vallejo et al., 2004). In this regard, we have conducted a study based on the possibility that BD is also associated with immunosenescence. As a result of our previous study, the frequency of senescent CD8+ T cells (CD8+CD27-CD28- T cells) was significantly higher in active BD than in disease controls and healthy controls (Yang et al., 2018). There were studies on gene expression of BD patients, but only to identify the levels of several mRNAs or conducted by microarray method, and no genetic profiling was conducted in relation to immunosenescent cells (Mizuki et al., 2010; Kirino et al., 2013; Kappen et al., 2015; Sousa I, 2015). This study was designed to reveal the genetic characteristics of BD CD8+ T cell by dividing groups into senescent (CD8+CD27-CD28-) and non-senescent (CD8+CD27+CD28+) T cells and to provide clue in
- 2 -
pathogenesis of BD and diagnostic biomarker for BD by conducting and comparing the whole transcription sequencing.
MATERIALS AND METHODS
A. Subjects
Patients with BD either who presented themselves for the first time or were monitored at the outpatient clinic of Department of Dermatology, Ajou University Hospital from November 2017 to February 2018, and healthy controls were enrolled. The diagnosis was made by BD criteria of either of International Study Group or Behçet’s Syndrome Research Committee of Japan (Mizushima Y, 1987; International Study Group for Behcet's Disease, 1990). For transcriptome analysis, a total of 18 patients and 18 healthy controls were enrolled and additional 5 patients and 5 HCs were selected to perform real-time PCR (Table 1). Patients with infectious or inflammatory disease and other systemic disease including liver or renal failure, diabetes mellitus, malignancies, and also BD patients with insufficient medical records were excluded. Informed consent was obtained from all subjects prior to enrollment. This study was approved by the Institutional Review Board (IRB number: AJIRB-BMR-GEN-14-462).
Table. 1. Baseline demographics of Behçet’s disease patients and healthy controls
A. Transcriptome analysis
total number male:female mean age, yr. (SD, range) BD 18 6:12 49.7 (9.0, 33-64) HC 18 7:11 44.3 (4.1, 38-53)
B. Real-time PCR
total number male:female mean age, yr. (SD, range) BD 5 3:2 51.6 (6.2, 42-61) HC 5 2:3 42.8 (3.0, 38-46)
- 4 -
B. Isolation of peripheral blood mononuclear cells (PBMCs)
Blood samples from the subjects were collected in standard 10mL heparin-treated vacutainer tubes (Vacutainer® Tubes; BectonDickinson, Stockholm, Sweden). PBMCs were isolated from each sample by Ficoll-Paque density gradient centrifugation (Ficoll Paque™ plus; Stem Cell Technologies, Vancouver, BC, Canada). Among them, CD8+ T cells were positively selected using CD8 microbeads and MS column. The purity of the obtained CD8 positive population was over 90%.
C. Sorting with cell surface marker flow cytometry
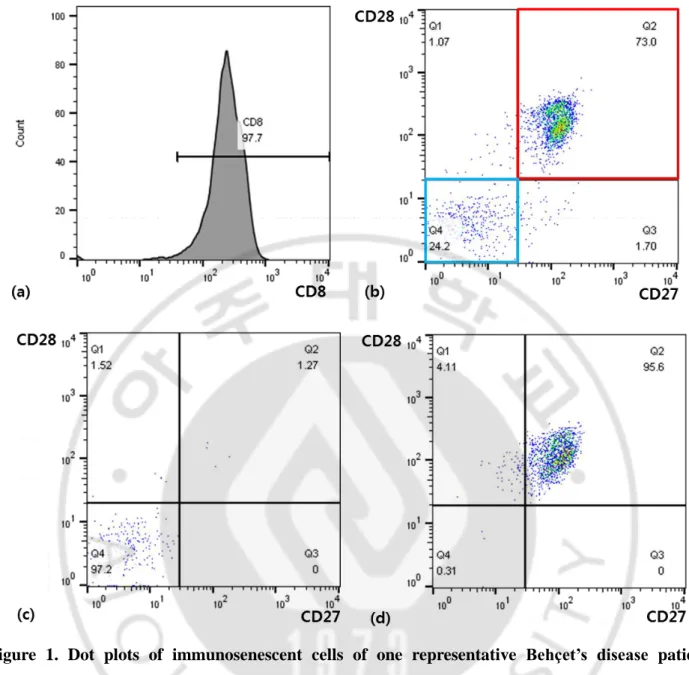
Expression of different cell surface markers was analyzed by flow cytometry including markers of immunosenescent cells (Figure 1). PBMCs were labelled with conjugated monoclonal antibodies as follows: FITC-anti-CD8, allophycocyanin (APC)-H7-anti-CD27 and APC-anti-CD28 (BD Pharmingen; BD Biosciences, San Jose, CA, USA). All antibodies were titrated to optimal concentrations according to the manufacturer’s protocol. PBMCs were incubated with the fluorescent-labelled monoclonal antibodies in dark at 4°C for 30 minutes. Using fluorescence-activated cell sorting (FACS) (BD FACS Aria Ⅲ; BD Biosciences, Moutain View, CA, USA), senescent CD8+ T cells (CD8+ CD27- CD28- cells) and non-senescent CD8+ T cells (CD8+ CD27+ CD28+ cells) were sorted.
Figure 1. Dot plots of immunosenescent cells of one representative Behçet’s disease patient analyzed with cell surface flow cytometry
a CD8+ T cells were positively selected using CD8 microbeads and MS column (Miltenyli Biotec,
CA, USA). The purity of the obtained CD8 positive population was over 90%. b Dot plots analyzing CD27 and CD28 expression on the CD8+ cells. Red box shows gated population of CD8+CD27+CD28+ cells and blue box shows that of CD8+CD27-CD28- cells. c Dot plots confirming CD27 and CD28 expression on the gated CD8+CD27-CD28- cells. d Dot plots confirming CD27 and CD28 expression on the gated CD8+CD27+CD28+ cells.
- 6 -
D. Cell culture and stimulation
After sorting, each group of cells were pooled together and cultured in RPMI 1640 medium supplemented with 2mM L-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin (RPMI 1640; Gibco-BRL, Grand Island, NY, U.S.A.) and 10% fetal bovine serum at 37℃ and 5% carbon dioxide. Cells were stimulated with anti-CD3 (500ng/ml, clone OKT3) (eBiosciences, San Diego, CA, U.S.A.) for 72 hours.
E. RNA extraction
Total RNA was extracted from stimulated sorting cells with the RNA isolation kit (NocleoSpin RNA XS kit) (Macherey-nagel, Düren, Germany) according to the protocol provided by the manufacturer.
F. RNA transcriptome analysis
We commissioned a RNA sample to Macrogen, Inc. for transcriptome analysis. cDNA libraries were made from the TruSeq RNA library kit by using 1 mg of total RNA. The protocol is composed of polyA-selected RNA extraction, RNA fragmentation, random hexamer-primed reverse transcription, and 100-nt paired-end sequenced by Illumina HiSeq2500. Quantification of the libraries was done by using quantitative polymerase chain reaction in accordance with the qPCR Quantification Protocol Guide, and qualification was done by using and Agilent Technologies 2100 Bioanalyzer. The RNA-seq reads were mapped to the human genome using TopHat (Trapnell et al., 2009) , the program that traces split-read alignments across splice junctions, and determined using Cufflinks software (Trapnell et al., 2010) with default options to measure the level of expression and to search for alternative spliced transcripts. The genome sequence used as reference (hg19, Genome Reference Consortium GRCh37) and footnote data was obtained from the UCSC website (http://genome.uscs.edu). The transcript counts were computated at the isoform and gene levels, and the relative transcript abundances were counted in Fragments per Kilobase of exon per Million fragments mapped (in FPKM) using Cufflinks.
G. Identification of differentially expressed genes (DEGs)
Transcripts with at least one zeroed FPKM value in every sample were excluded from the analysis to identify the difference in gene expression levels. Every FPKM value was added 1 to simplify the log2 transformation. Modified data were logarithm-transformed and normalized by the quantile method. Differentially expressed genes were adjusted to |log2 fold change| >2 for each comparison pair.
H. Ingenuity Pathway Analysis (IPA®)
Genes with a |log2 fold change| greater than 2 were considered significant and submitted to Ingenuity
Pathway Analysis for functional study and identification of significant biological pathways.
I. RNA preparation and real-time polymerase chain reaction (PCR)
For validation of genes that show large fold change value in the results of transcriptome sequencing, five additional BD patients and five HCs were selected to collect RNA and perform a real-time PCR. RNA was extracted using trizol (Invitrogen, Carlsbad, CA, USA) and reverse transcription of RNA was
performed using dNTP’s and an oligo (dT) primer (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The samples were incubated for 5 minutes at 65℃. The cDNA was amplified in a 20-μl total volume using the SuperscriptTM III kit (Invitrogen, Carlsbad, CA, USA), following the manufacturer’s protocol. After the amplification, cDNA was analyzed by real-time PCR using the ABI Prism 7000 Sequence Detection System (Applied Biosystems, Foster, CA, USA) according to the instruction of manufacturer using the 2–ΔΔCt method. Primers and internal probes for RGS11 (Hs01594651_m1), SHANK1 (Hs00211718_m1), AHRGEF10 (Hs00405154_m1), LOC102724428 (Hs00545020_m1), UBE2F-SCLY (Hs00996436_m1) were purchased from Thermo Scientific. The primer sequences of the tests performed using SYBR Green Supermix were as follows: TRPV3; 5’-GTTAGCTACCCGCATTAAGCCTGA-3’, 5’-AGCAATTCTGGAATTCCCAGCTC-3’, COL5A1; 5’-CCTGCATTGTCTGTGGTGTGA-3’, 5’-AGCGCATGGCATGACTGAA-3’, CD302; GCCCAGATGATATCCTACTAGGC-3’, GCAGAAAAGCACAGGTGTCAACT-3’, NBL1; 5’-TCCACAGAGTCCCTGGTTCACT-3’, 5’-GCTACAGTGCAGGATCTTCTCC-3’.
J. Statistical analysis
Statistical analysis was performed using SPSS software (Version 22.0, IBM, Armonk, NY, USA). The Pearson Chi-square test and Fisher’s exact test were used for dichotomous and nominal variables. And the Mann-Whitney test was used for comparison of continuous variables. p value less than 0.05 was considered statistically significant.
- 8 -
RESULTS
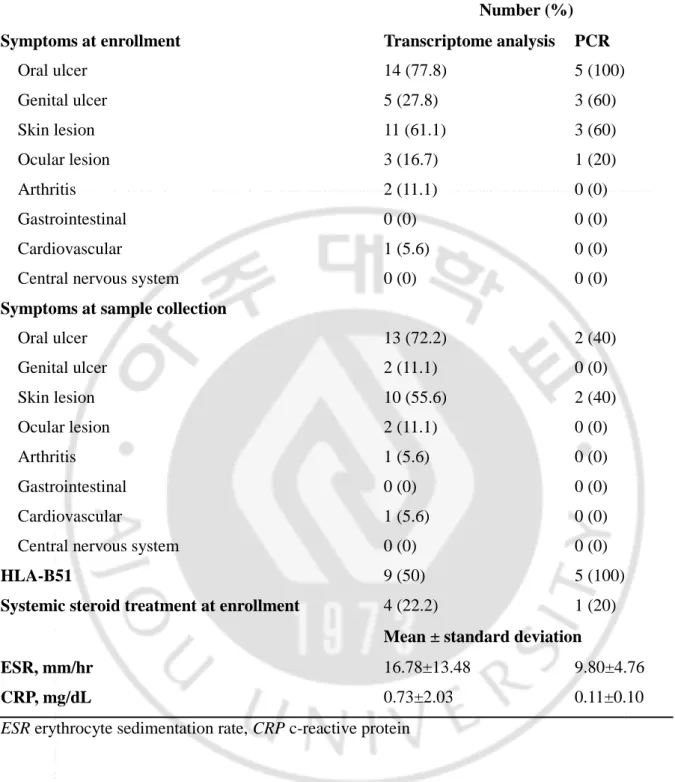
A total of 18 BD patients and 18 HCs for transcriptome analysis and additional 5 patients and 5 HCs for real-time PCR were enrolled in the study. Clinical characteristics of BD patients are summarized in Table 2. Many patients accompanied by mucocutaneous symptoms such as the oro-genital ulcer and skin lesion. For patients with transcriptome analysis, the number of patients who were carrying out a
systematic steroid treatment at the time of enrollment was four, accounting for 22.2% of patients. The average ESR level in patients was 16.78 and the CRP was 0.73. In nine cases, half of the total patients were positive for HLA-B51.
Table 2. Clinical characteristics of Behçet’s disease patients
Number (%)
Symptoms at enrollment Transcriptome analysis PCR
Oral ulcer 14 (77.8) 5 (100) Genital ulcer 5 (27.8) 3 (60) Skin lesion 11 (61.1) 3 (60) Ocular lesion 3 (16.7) 1 (20) Arthritis 2 (11.1) 0 (0) Gastrointestinal 0 (0) 0 (0) Cardiovascular 1 (5.6) 0 (0) Central nervous system 0 (0) 0 (0)
Symptoms at sample collection
Oral ulcer 13 (72.2) 2 (40) Genital ulcer 2 (11.1) 0 (0) Skin lesion 10 (55.6) 2 (40) Ocular lesion 2 (11.1) 0 (0) Arthritis 1 (5.6) 0 (0) Gastrointestinal 0 (0) 0 (0) Cardiovascular 1 (5.6) 0 (0) Central nervous system 0 (0) 0 (0)
HLA-B51 9 (50) 5 (100)
Systemic steroid treatment at enrollment 4 (22.2) 1 (20)
Mean ± standard deviation
ESR, mm/hr 16.78±13.48 9.80±4.76
CRP, mg/dL 0.73±2.03 0.11±0.10
- 10 -
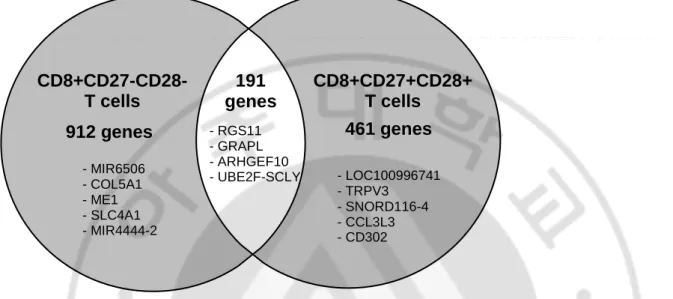
RNA sequencing confirmed that a total of 1,564 genes showed differential expression over the fold change 2 (fc2) between BD patients and healthy controls. Figure 2 is a Venn diagram depicting the number of differentially expressed genes in senescent and non-senescent CD8+ T cells of BD patients compared to HCs. mRNA levels of 1103 genes were different in CD8+CD27-CD28- T cells between BD patients and HCs, whereas, mRNA levels of 652 genes differed in CD8+CD27+CD28+ T cells.
Figure 2. Venn diagram of differentially expressed genes in senescent and non-senescent CD8+ T cells of Behçet’s disease patients compared to healthy controls
mRNA levels of 1103 genes were different in CD8+CD27-CD28- T cells between Behçet’s disease patients and healthy controls, whereas, mRNA levels of 652 genes differed in CD8+CD27+CD28+ T cells. Genes showing the biggest fold change within each group were marked within the Venn diagram.
CD8+CD27-CD28- T cells 912 genes 461 genes 191 genes CD8+CD27+CD28+ T cells - MIR6506 - COL5A1 - ME1 - SLC4A1 - MIR4444-2 - RGS11 - GRAPL - ARHGEF10 - UBE2F-SCLY - LOC100996741 - TRPV3 - SNORD116-4 - CCL3L3 - CD302
For the validation and utilization as a biomarker of RNA sequencing result, top ten upregulated and downregulated genes showing a high absolute value of fold change in BD patients compared to HCs were selected to be shown in Table 3 and Table 4 according to their cell group. These tables show top ten upregulated and downregulated genes found among BD patients within the senescent and non-senescent CD8+ T cell group respectively. Positive and negative log2 fold change values mean
up-and-downregulation of gene expression in BD patients relative to the gene expression in healthy controls. RGS11 appeared to be the only gene in both groups that was included in Top 10. In addition, RGS11 represented positive values for the fold change in both groups.
Table 3. Top ten genes based on expression fold change order in CD8+CD27-CD28- T cells of Behçet’s disease patients compared to healthy controls
Gene ID Gene Symbol Description log2 fold change of
RNA sequencing
4681 NBL1 neuroblastoma 1, DAN family BMP antagonist 131.4
8786 RGS11 regulator of G protein signaling 11 107.8
50944 SHANK1 SH3 and multiple ankyrin repeat domains 1 73.0
102465252 MIR6506 microRNA 6506 37.2
1289 COL5A1 collagen type V alpha 1 chain 36.9
4199 ME1 malic enzyme 1 32.9
57282 SLC4A10 solute carrier family 4 member 10 30.7
6571 SLC18A2 solute carrier family 18 member A2 29.4
5783 PTPN13 protein tyrosine phosphatase, non-receptor type 13 27.9
100846999 MIR4444-2 microRNA 4444-2 -88.7
Table 3 is a summary of top ten up-and-downregulated genes found in Behçet’s disease patients vs. healthy controls among senescent cells. Positive and negative fold change values mean upregulation and downregulation respectively, of gene expression in senescent cells of Behçet’s disease patients relative to the gene expression in senescent cells of healthy controls.
- 12 -
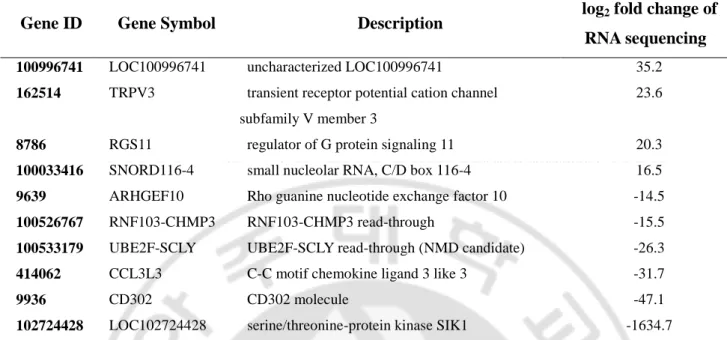
Table 4. Top ten genes based on expression fold change order in CD8+CD27+CD28+ T cells of Behçet’s disease patients compared to healthy controls
Gene ID Gene Symbol Description log2 fold change of
RNA sequencing
100996741 LOC100996741 uncharacterized LOC100996741 35.2
162514 TRPV3 transient receptor potential cation channel subfamily V member 3
23.6
8786 RGS11 regulator of G protein signaling 11 20.3
100033416 SNORD116-4 small nucleolar RNA, C/D box 116-4 16.5
9639 ARHGEF10 Rho guanine nucleotide exchange factor 10 -14.5
100526767 RNF103-CHMP3 RNF103-CHMP3 read-through -15.5
100533179 UBE2F-SCLY UBE2F-SCLY read-through (NMD candidate) -26.3
414062 CCL3L3 C-C motif chemokine ligand 3 like 3 -31.7
9936 CD302 CD302 molecule -47.1
102724428 LOC102724428 serine/threonine-protein kinase SIK1 -1634.7
Table 4 is a summary of top ten up-and-downregulated genes found in Behçet’s disease patients vs. healthy controls among senescent cells. Positive and negative fold change values mean upregulation and downregulation respectively, of gene expression in senescent cells of Behçet’s disease patients relative to the gene expression in non-senescent cells of healthy controls.
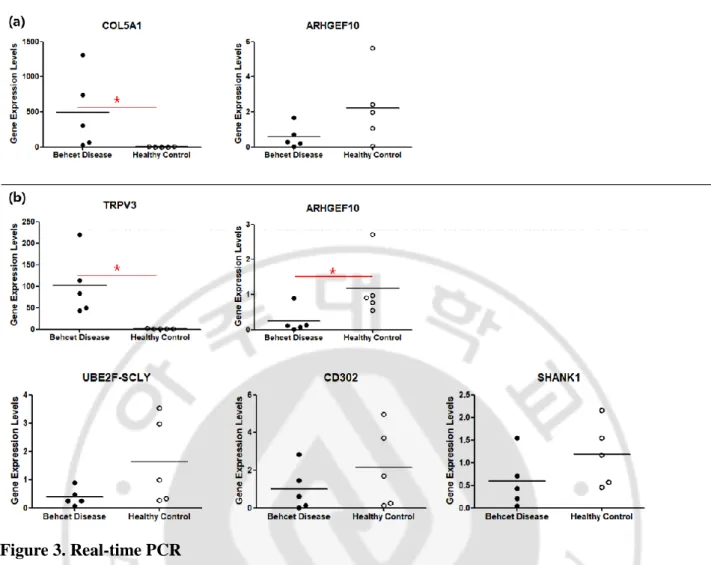
For the validation of RNA sequencing result, we performed the real-time PCR on eight genes: RGS11, SHANK1, AHRGEF10, LOC102724428, CD302, COL5A1, TRPV3 and UBE2F-SCLY. The genes to perform PCR were selected from top DEGs except for the miRNA and uncharacterized genes. Among them, Figure 3 included genes that showed the consistent tendency with the result of transcriptome analysis. COL5A1, ARHGEF10 showed the same tendency with the transcriptome analysis in the BD CD27-CD28- subsets compared to HCs, among which the statistical significance was shown in COL5A1. Meanwhile, TRPV3, ARHGEF10, UBE2F-SCLY, CD302, and SHANK1 showed the same tendency with the transcriptome analysis in the BD CD27+CD28+ subsets compared to HCs, and the statistical significance was found in TRPV3 and ARHGEF10.
Figure 3. Real-time PCR
It presents expression levels of genes in CD8+CD27-CD28- T cell group (a) and CD8+CD27+CD28+ T cell group (b) respectively. It shows genes only that the results of the PCR match the results of the transcriptome analysis. a Relative comparison of the mean PCR bands value between Behçet’s disease patients and healthy controls in CD8+CD27-CD28- T cell group b Relative comparison of the mean PCR bands value between Behçet’s disease patients and healthy controls in CD8+CD27+CD28+ T cell group (* p<0.05)
- 14 -
DEGs with log2 value of fold change higher than 2 were submitted to IPA for functional study and
identifying significant pathways. The result of IPA was presented in Table 5 and contains the
information of top biological functions of DEGs between BD and HCs including subcategories such as “Diseases and Disorders”, “Molecular and Cellular Functions” and “Physiological System Development and Function” with their P-value. The P-value in this table was determined by comparing the number of genes found in each category (number of genes) with the total number of known genes within the IPA software database; “Query Ingenuity Knowledge Base” .
Table 5. Top biological functions of differentially expressed genes between Behçet’s disease patients and healthy controls as presented by Ingenuity Pathway Analysis (IPA)
Biological Functions P-value Number of molecules Disease and disorders:
Cancer 1.30E-03 - 1.48E-25 997
Organismal Injury and Abnormalities 1.33E-03 - 1.48E-25 1012
Dermatological Diseases and Conditions 9.09E-04 - 1.50E-20 620
Gastrointestinal Disease 1.33E-03 - 1.05E-19 932
Reproductive System Disease 7.38E-04 - 2.16E-13 633
Molecular and Cellular Functions:
Cellular Movement 1.29E-03 - 9.33E-11 256
Cell-To-Cell Signaling and Interaction 1.30E-03 - 2.68E-07 232
Molecular Transport 8.52E-04 - 2.70E-06 161
Cellular Function and Maintenance 1.33E-03 - 6.48E-06 296
Cellular Assembly and Organization 1.17E-03 - 8.77E-06 200 Physiological System Development and Function:
Immune Cell Trafficking 1.29E-03 - 6.70E-09 127
Hematological System Development and Function 1.33E-03 - 5.21E-08 192
Nervous System Development and Function 1.30E-03 - 2.68E-07 186
Tissue Development 1.33E-03 - 4.58E-06 234
Cardiovascular System Development and Function 1.30E-03 - 7.73E-06 146
Top biological functions of differentially expressed genes between Behçet’s disease patients and healthy controls in this study provided by Ingenuity Pathway Analysis (IPA®). The P-value in this table was determined by IPA by comparing the number of genes found in each category with the number of known genes within the IPA software system.
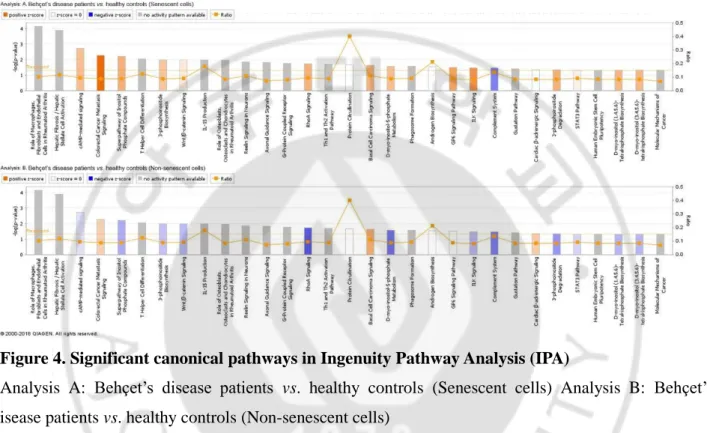
Major canonical pathways found by IPA are illustrated in Figure 4. Analysis A and analysis B represents the canonical pathways of senescent and non-senescent cell respectively through –log (P-value). Among the pathways in which we could find its activity pattern, eleven pathways showed their activity in the opposite direction depending on whether the cell was immunosenescent or not. Of those pathways, cAMP-mediated signaling, RhoA signaling, and ILK signaling showed positive Z-score in patients with BD in the senescent cells, while showed negative Z-score in patients with BD in the non-senescent cell groups.
Figure 4. Significant canonical pathways in Ingenuity Pathway Analysis (IPA)
Analysis A: Behçet’s disease patients vs. healthy controls (Senescent cells) Analysis B: Behçet’s disease patients vs. healthy controls (Non-senescent cells)
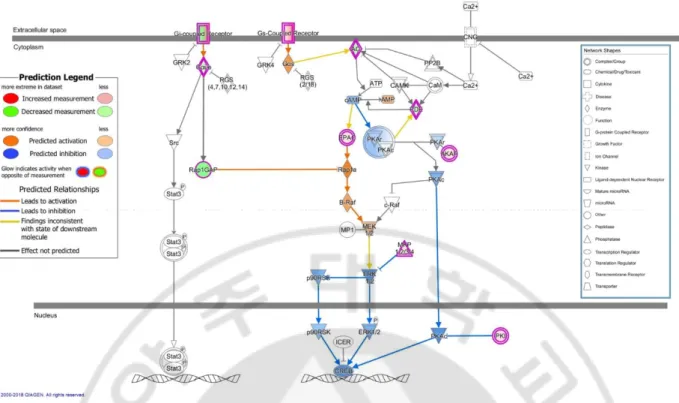
Figure 5 is a detailed diagram of the cAMP-mediated signaling which is identified as the most significant pathway among the major canonical pathways. The significancy of canonical pathway was determined by IPA by comparing the number of genes found in each pathway of known genes within the IPA software system. In our study, there were a total of 24 DEGs corresponding to cAMP-mediated signing pathways, including HTR6, P2RY14, TBXA2R, PDE7B, and AKAP12. This diagram shows the “cAMP-mediated signaling” as related to significant DEGs in BD patients from the sample population. Genes that are highlighted in green are considered to be downregulated in BD patients and genes that are highlighted in red are considered to be upregulated in BD patients. In case of molecules with orange and blue colors were not included in the DEG list of our study but expected to be activated and inhibited with the prediction function of IPA software system. There are differential expressions on cAMP-mediated signal pathways through IPA, including G-coupled receiver, Rap1GAP, Gay/o, AO, PDE, EPAC, AKAP, MKP1/2/2/4PKI.
- 16 -
Figure 5. cAMP-mediated signaling in Behçet’s disease patients vs. healthy controls (senescent cells) with prediction: One of the significant canonical pathways in IPA
The c-AMP-mediated signaling pathway showing differentially expressed genes in Behçet’s disease patients vs. healthy controls (senescent cells) with prediction. Canonical pathways were identified and pictorialized by Ingenuity Pathway Analysis (IPA®) software. This pathway shows the c-AMP-mediated signaling as related to significant differentially expressed genes in Behçet’s disease patients vs. healthy controls (senescent cells) with prediction. Genes that are colored in green are considered to be downregulated and colored in blue are predicted to be inhibited in Behçet’s disease patients. Genes that are colored in red are considered to be upregulated and colored in orange are predicted to be activated in Behçet’s disease patients.
DISCUSSION
In this study, we revealed the whole gene expression profiling of CD8+ T cell subpopulation of BD patients compared to HCs. Before the sequencing, we divided the CD8+ T cell population into
CD8+CD27-CD28- and CD8+CD27+CD28+ subgroups and compared the result of each groups. Previous genetic studies of BD were confirmed through microarray method or RNA sequencing on some limited pre-selected mRNAs (Chen et al., 2016; Oguz et al., 2016; Okuzaki et al., 2016; Qin et al., 2016; Kang et al., 2017; Takeuchi et al., 2017; Yu et al., 2017; Puccetti et al., 2018). In contrast, we analyzed using a new technique “whole RNA sequencing”, which was the first to be implemented in BD. For rheumatoid arthritis, which is thought to share a similar pathogenesis with BD, a study have found four novel genes by transcriptome analysis using PBMC sample, as shown in this study. (Wang et al., 2017). There is a difference between this study that we used only sorted CD8+ T cell, which showed the difference in the frequency of senescent cells in the previous study for sequencing (Yang et al., 2018).
Of the DEGs with more than twice the log2 fold change for each groups, 191 genes were common, and
the remaining 1373 were not overlapped. Top-ranked up/downregulated genes were found differently between CD8+CD27-CD28- and CD8+CD27+CD28+ T cell groups. RNA sequencing, as shown in Table 3 and 4, found novel genes that exhibit large fold changes compared to HCs.
After listing the top ten DEGs with the largest absolute value of the fold changes in the senescent and non-senescent CD8+ T cell groups respectively, we noted five genes: RGS11, LOC102724428,
ARHGEF10, SHANK1 and UBE2F-SCLY.
First of all, RGS11 (Regulator of G-protein Signaling 11) is a G-protein signaling regulator, and proteins coded by this gene belong to the G protein signaling modulator (RSS) family. Members of the RGS family act as GTP hydrolase-activated protein in alpha subunit of heterotrimeric G-protein signaling protein. By increasing the GTPase activity of the G-protein alpha subunits, they inhibit the signaling and lead to the form of inactive GDP combination status. In addition, RGS11 is a protein in the R7 RGS family that is overexpressed in metastatic tumor cells and induces an increase in Rac-1-dependent tumor cell movement through c-Raf/ERK/FAK signaling activation (Yang et al., 2016). Collapse of endothelial barrier which induced by inflammatory stimuli like as bacterial LPS, thrombin , or TNF-α is suggested to be at least in part caused by loss of intracellular cAMP and Rac1 inactivation (Schlegel and Waschke, 2014). Therefore, it is possible that the RGS11 gene expression, which was upregulated in the both cell group, activated cAMP/Rac1signaling and induced the endothelial barrier disruption of BD patients. It is known that 12.8% of BD patients have vascular involvement and can affect blood vessels of all types and sizes (Fei et al., 2013). The endothelial barrier disruption by RGS11 gene expression which activates cAMP/Rac1 signaling may be related to the vascular involvement of
- 18 -
BD.
LOC102724428 is a serine/threonine-protein kinase salt-inducible kinase (SIK) 1. The gene encodes serine/threonine-protein kinase including ubiquitin-related domains. It is also expected to affect ATP bonds, magnesium ion bonds, and protein phosphorylation. Protein kinase A phosphorylates a number of substances participating in T-cell activation as a serine/threonine kinase (Arumugham and Baldari, 2017). Although mRNA expression is observed in most tissues, including lymph nodes and spleen, there is still not much research on its function. Although it was not a study of T cells, there were several papers about how the SIKs work on the macrophage. It is demonstrated that SIKs inhibit the formation of regulatory macrophages and consequentially induce the rise of many characteristic markers of regulatory macrophages such as IL-10 and other anti-inflammatory molecules (Clark K, 2012). Therapeutic aiming on this macrophage population at inflammatory site may help long-term disease modification for patients who have been suffered from chronic inflammatory disease such as rheumatoid arthritis (Darling et al., 2017). According to Darling et al., when three isoforms of SIK, SIK1,SIK2 or SIK3 are suppressed, a macrophage phenotype is triggered, which secretes high levels of
anti-inflammatory cytokines such as interleukin (IL)-10, and very low levels of pro-anti-inflammatory cytokines, including tumor necrosis factor (TNF) α (Darling et al., 2017). In this study, increased expression of LOC102724428 was observed showing high RNA sequencing fold change in BD patient of senescent cell group. SIK expression reduces the secretion of an anti-inflammatory cytokine, IL-10, in
macrophage, which is in line with the phenomenon of showing IL-10 polymorphisms in BD patients (Afkari et al., 2018). However, there was no significant association between BD and SNPs in IL-10 mediated intracellular signaling in Korea patients in one study (Kang EH, 2014).
ARHGEF10 (Rho Guanine nucleotide exchange factor 10) codes the Rho Guanine nucleotide exchange factor (hereafter, Rho GEF). The Rho GEF controls the activity of small Rho GTPases by stimulating the exchange of guanine diphosphate (GDP) to guanine triphosphate (GTP). ARHFEF10 is related with invasion of breast cancer cells and function of platelet by controlling the membrane trafficking through combining with RAB6A and RAB8A in exocytic vesicles. And the variant of this gene is also associated with the genetic neurological disorder, “Charcot-Marie-Toot disease” which affects the activity of delta 6 unsaturase. In addition, it is revealed by single-nucleotide polymorphism analysis that atherothrombotic stroke is highly associated with the genes encoding ARHGEF10 (Matsushita et al., 2010), indicating their relationship with blood vessel homeostasis and function. However, specific mechanisms and functional roles of Rho-GEFs in mechanotransduction of vascular endothelial cells are uncertain (Abiko et al., 2015). RNA sequencing fold change of ARHGEF10 in the both senescent and non-senescent cell group was decreased in this study. When ARHGEF10 is said to play a role in blood vessel homeostasis and function, it is likely to be related to the vascular involvement in BD, as in RGS11.
SHANK1 (SH3 and multiple ankyrin repeat domains 1) codes SH3 domain and the SHANK family of proteins that contain the ankyrin repetition (Monteiro and Feng, 2017). This family of proteins serves as the scaffold protein needed for the development and functioning of connected synapses. The expression of SHANK1 is highest in the cerebellum, medium in spleen and bone marrow, and low in cerebral lymph nodes. It is also present in the human monocyte THP-1 cells and the human leukemia U937 cells and shows increase of expression in ovarian cancer. Protein in this family is involved in signaling by combining various membrane receptors and cytoplasm proteins using PDZ domains.
UBE2F-SCLY is located between the neighboring Ubiquitin-conjugating enzyme E2F (UBE2F) and Selenocysteine lyase (SCLY) and expressed by read-through-transcription. UBE2F-SCLY converged protein is expected to control cholesterol metabolism through selenium metabolism. Our target patients showed decreased RNA sequencing fold change in both senescent and non-senescent cell group. This can be associated with occurrence of metabolic syndrome in addition to other autoimmune disorders in BD patients (Yalcin et al., 2013).
Among top-ranked up/downregulated genes, we conducted real-time PCR for validating the result of the RNA sequencing on eight genes except miRNA and uncharacterized genes whose function was not revealed. Unfortunately, not all PCR results matched the results of sequencing. This may be due to the PCR and transcriptome analysis were conducted using samples from other patient population,
respectively. Genes with propensity to match transcriptome analysis results were COL5A1 in
CD8+CD27-CD28- T cells, and TRPV3 and ARHGEF10 in CD8+CD27+CD28+ T cells, with the result of PCR having statistically significant differences in the expression level between the BD patients and HCs. ARHGEF10 in the CD8+CD27-CD28- T cells, and UBE2F-SCLY, CD302, SHANK1 in
CD8+CD27+CD28+ T cells also showed same tendency with the transcriptome analysis, although they did not show statistically significant difference in the expression level between BD patients and HCs. There have been no reports of the above genes being linked to other autoimmune or autoinflammatory diseases including BD, but it seems likely that they could be used as biomarkers for BD. There are several reasons why there was differences between transcriptome analysis and PCR results. Individuals with a very large variation of expression level, may fall into the pool and have a tendency error, which is also the limit of implementing pooling methods. In addition, the limitation of not dividing BD patient by their disease activity was active or inactive, may have affected the results. It is also possible that
differences in results between PCR and sequencing occurred due to the differences in activity that controls expression within the pathways.
In this study, we identified 1,564 DEGs with variable size of fold change. All the 1,564 DEGs were subjected to Ingenuity Pathway Analysis. A total of eleven major pathways of DEGs analyzed through IPA were found to show opposite activity pattern in CD8+CD27-CD28- T cells and CD8+CD27+CD28+ T cells. These pathways include cAMP-mediated signaling, RhoA signaling, and
- 20 -
ILK signaling, which are recently assumed to be related to immune-regulating actions (Chen et al., 2013; Motoyoshi et al., 2014; Schlegel and Waschke, 2014; Abiko et al., 2015; Arumugham and Baldari, 2017; Hauck et al., 2017). This result demonstrates that these eleven pathways may be a key of the senescent CD8+ T cells to affect in the pathogenesis of BD.
The most significant canonical pathway resulted from the IPA was mediated signaling. cAMP-mediated signaling is a field of increasing interest recently, with close association with the functioning of immune cells. There are no reports of link between the cAMP-mediated signaling and BD, but we thought it was important because it suggests a connection of BD with autoimmune and
autoinflammatory. cAMP has an overall inhibitory effect on both innate and adaptive immune cell types, including neutrophils, macrophages, and T and B lymphocytes. cAMP is necessary for T cell activation and optimal immune function. Contrary to the result of the transient rise in cAMP triggered by TCR engagement, a continuous elevation of cAMP leads to suppression of T cell activation, proliferation, and chemotaxis (Arumugham and Baldari, 2017). Also, therapeutic strategies to interrupt or enhance cAMP generation may have immunoregulatory potential in autoimmune and inflammatory disorders (Raker et al., 2016). For example, a study has shown that Apremilast, a PDE-4 inhibitor which suppresses
inflammation through cAMP-dependent mechanism has shown efficacy in treating oral ulcers of BD patients (Hatemi et al., 2015; Perez-Aso et al., 2015). Therefore, we could consider the possibility that the activation of the cAMP-mediated signaling pathways sustained by senescent T cells may cause dysfunction of the cells. As Figure 5 shows, there are differential expressions in our data on cAMP-mediated signal pathways through IPA, including G-coupled receiver, Rap1GAP, Gay/o, AO, PDE, EPAC, AKAP, MKP1/2/2/4PKI. Validation of these molecules would help to ensure the relevance with BD, but verification at the protein level could not take place because it required a large amount of cells. Validating experiments in protein level and conducting PCR for cAMP-signaling pathway-related genes for a large sample size, will help to find more clues.
The limitation of this study is a small sample size. BD is a very rare disease, so it was difficult to enroll patients to study. Moreover, RNA sequencing required relatively large amount of blood samples, so we were forced to pool the RNA samples of patients to secure the amount of RNA for sequencing. RNA sequencing could not be performed repeatedly in the same manner. However, we tried to validate the transcriptome analysis result by performing real-time PCR on several genes instead of repeating the RNA sequencing. In one RA study, they analyzed two kinds of sequencing in the initial experimental phase. After making comparisons of results of single-sample tests with the same control, they found that the number of up-/downregulated differentially expressed genes and reported differentially expressed genes were not significantly different among the comparisons. And they also mentioned that there are some benefits of pooling the samples to minimize the variability associated with differences between
individual samples (Wang et al., 2017). In addition, it is difficult to completely rule out the possibility that the anti-CD3 stimulus may have affected the gene expression.
- 22 -
CONCLUSION
This is the first study to investigate the differential gene expression through RNA sequencing of CD8+ T cells of BD patients compared to HCs. This study was also the first to separate senescent CD8+ T cells and nonsenescent CD8+ T cells to perform RNA sequencing, respectively. As a result of
sequencing in each group, only 191 of the total 1,564 DEGs were common, and only one gene (RGS11) was in common among the top-ranked up/down regulated genes, so the difference between the senescent and non-senescent CD8+ T cells is thought to be significant. Therefore, there is likely to be having masking effect if using total CD8+ T cells without subgrouping. The strength of this study is that it is possible to exclude the possible effects of senescent CD8+ T cells on BD patients because prior studies have shown an increased frequency of senescent CD8+ T cells in BD patients (Yang et al., 2018). Also, we analyzed top 10 DEGs with a large-fold change values. Using these differentially expressed genes may be useful for developing biomarkers in BD that can predict the treatment response as well as help diagnosing easily. Consequently, we hope that the genetic profiling we have conducted can be used as a key to research approaching BD pathogenesis.
REFERENCES
1. Ingenuity Systems [http://www.ingenuity.com]
2. Abiko H, Fujiwara S, Ohashi K, Hiatari R, Mashiko T, Sakamoto N, Sato M, Mizuno K: Rho guanine nucleotide exchange factors involved in cyclic-stretch-induced reorientation of vascular endothelial cells. J Cell Sci 128: 1683-1695, 2015
3. Afkari B, Babaloo Z, Dolati S, Khabazi A, Jadidi-Niaragh F, Talei M, Shanehbandi D, Mahmoudi S, Hazhirkarzar B, Sakhinia E: Molecular analysis of interleukin-10 gene polymorphisms in patients with Behcet's disease. Immunol Lett 194: 56-61, 2018
4. Arumugham VB, Baldari CT: cAMP: a multifaceted modulator of immune synapse assembly and T cell activation. J Leukoc Biol 101: 1301-1316, 2017
5. Ben Ahmed M, Houman H, Miled M, Dellagi K, Louzir H: Involvement of chemokines and Th1 cytokines in the pathogenesis of mucocutaneous lesions of Behcet's disease. Arthritis
Rheum 50: 2291-2295, 2004
6. Chen F, Xu L, Zhao T, Xiao X, Pan Y, Hou S: Genetic Variation in the REL Gene Increases Risk of Behcet's Disease in a Chinese Han Population but That of PRKCQ Does Not. PLoS
One 11: e0147350, 2016
7. Chen ML, Yi L, Jin X, Liang XY, Zhou Y, Zhang T, Xie Q, Zhou X, Chang H, Fu YJ, Zhu JD, Zhang QY, Mi MT: Resveratrol attenuates vascular endothelial inflammation by inducing autophagy through the cAMP signaling pathway. Autophagy 9: 2033-2045, 2013
8. Clark K MK, Petkevicius K, Kristariyanto Y, Zhang J, Choi HG, Peggie M, Plater L, Pedrioli PG, McIver E, Gray NS, Arthur JS, Cohen P.: Phosphorylation of CRTC3 by the salt-inducible kinases controls the interconversion of classically activated and regulatory macrophages. Proc
Natl Acad Sci U S A. 109(42): 16986-16991, 2012
9. Darling NJ, Toth R, Arthur JS, Clark K: Inhibition of SIK2 and SIK3 during differentiation enhances the anti-inflammatory phenotype of macrophages. Biochem J 474: 521-537, 2017 10. Direskeneli H, Saruhan-Direskeneli G: The role of heat shock proteins in Behcet's disease. Clin
Exp Rheumatol 21: S44-48, 2003
11. Ekinci NS, Alpsoy E, Karakas AA, Yilmaz SB, Yegin O: IL-17A has an important role in the acute attacks of Behcet's disease. J Invest Dermatol 130: 2136-2138, 2010
12. Fei Y, Li X, Lin S, Song X, Wu Q, Zhu Y, Gao X, Zhang W, Zhao Y, Zeng X, Zhang F: Major vascular involvement in Behcet's disease: a retrospective study of 796 patients. Clin Rheumatol 32: 845-852, 2013
13. Goronzy JJ, Lee WW, Weyand CM: Aging and T-cell diversity. Exp Gerontol 42: 400-406, 2007
14. Hatemi G, Melikoglu M, Tunc R, Korkmaz C, Turgut Ozturk B, Mat C, Merkel PA, Calamia KT, Liu Z, Pineda L, Stevens RM, Yazici H, Yazici Y: Apremilast for Behcet's syndrome--a phase 2, placebo-controlled study. N Engl J Med 372: 1510-1518, 2015
15. Hatemi G, Silman A, Bang D, Bodaghi B, Chamberlain AM, Gul A, Houman MH, Kotter I, Olivieri I, Salvarani C, Sfikakis PP, Siva A, Stanford MR, Stubiger N, Yurdakul S, Yazici H, EULAR Expert Committee: EULAR recommendations for the management of Behcet disease.
Ann Rheum Dis 67: 1656-1662, 2008
16. Hauck SM, Lepper MF, Hertl M, Sekundo W, Deeg CA: Proteome Dynamics in Biobanked Horse Peripheral Blood Derived Lymphocytes (PBL) with Induced Autoimmune Uveitis.
Proteomics 17, 2017
17. Imirzalioglu N, Dursun A, Tastan B, Soysal Y, Yakicier MC: MEFV gene is a probable susceptibility gene for Behcet's disease. Scand J Rheumatol 34: 56-58, 2005
18. International Study Group for Behcet's Disease: Criteria for diagnosis of Behcet's disease.
Lancet 335: 1078-1080, 1990
- 24 -
mediated signalling pathways in Korea patients with Behçet's disease. Clin Exp Rheumatol. 32 (Suppl. 84): S27-S32, 2014
20. Kang EH, Kim S, Park MY, Choi JY, Choi IA, Kim MJ, Ha YJ, Lee EY, Lee YJ, Lee EB, Kang C, Song YW: Behcet's disease risk association fine-mapped on the IL23R-IL12RB2 intergenic region in Koreans. Arthritis Res Ther 19: 227, 2017
21. Kappen JH, Medina-Gomez C, van Hagen PM, Stolk L, Estrada K, Rivadeneira F, Uitterlinden AG, Stanford MR, Ben-Chetrit E, Wallace GR, Soylu M, van Laar JA: Genome-wide association study in an admixed case series reveals IL12A as a new candidate in Behcet disease.
PLoS One 10: e0119085, 2015
22. Kirino Y, Bertsias G, Ishigatsubo Y, Mizuki N, Tugal-Tutkun I, Seyahi E, Ozyazgan Y, Sacli FS, Erer B, Inoko H, Emrence Z, Cakar A, Abaci N, Ustek D, Satorius C, Ueda A, Takeno M, Kim Y, Wood GM, Ombrello MJ, Meguro A, Gul A, Remmers EF, Kastner DL: Genome-wide association analysis identifies new susceptibility loci for Behcet's disease and epistasis between HLA-B*51 and ERAP1. Nat Genet 45: 202-207, 2013
23. Liang L, Tan X, Zhou Q, Zhu Y, Tian Y, Yu H, Kijlstra A, Yang P: IL-1beta triggered by peptidoglycan and lipopolysaccharide through TLR2/4 and ROS-NLRP3 inflammasome-dependent pathways is involved in ocular Behcet's disease. Invest Ophthalmol Vis Sci 54: 402-414, 2013
24. Markovic-Plese S, Cortese I, Wandinger KP, McFarland HF, Martin R: CD4+CD28- costimulation-independent T cells in multiple sclerosis. J Clin Invest 108: 1185-1194, 2001 25. Matsushita T, Ashikawa K, Yonemoto K, Hirakawa Y, Hata J, Amitani H, Doi Y, Ninomiya T,
Kitazono T, Ibayashi S, Iida M, Nakamura Y, Kiyohara Y, Kubo M: Functional SNP of ARHGEF10 confers risk of atherothrombotic stroke. Hum Mol Genet 19: 1137-1146, 2010 26. Mizuki N, Meguro A, Ota M, Ohno S, Shiota T, Kawagoe T, Ito N, Kera J, Okada E, Yatsu K,
Song YW, Lee EB, Kitaichi N, Namba K, Horie Y, Takeno M, Sugita S, Mochizuki M, Bahram S, Ishigatsubo Y, Inoko H: Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behcet's disease susceptibility loci. Nat Genet 42: 703-706, 2010
27. Mizushima Y IG, Mimura Y: Guide for the diagnosis of Behçet’s disease. Japan: Report of Behçet’s Disease Research, 1987
28. Monteiro P, Feng G: SHANK proteins: roles at the synapse and in autism spectrum disorder.
Nat Rev Neurosci 18: 147-157, 2017
29. Motoyoshi S, Yamamoto Y, Munesue S, Igawa H, Harashima A, Saito H, Han D, Watanabe T, Sato H, Yamamoto H: cAMP ameliorates inflammation by modulation of macrophage receptor for advanced glycation end-products. Biochem J 463: 75-82, 2014
30. Na SY, Park MJ, Park S, Lee ES: Up-regulation of Th17 and related cytokines in Behcet's disease corresponding to disease activity. Clin Exp Rheumatol 31: S32-40, 2013
31. Oguz AK, Yilmaz ST, Oygur CS, Candar T, Sayin I, Kilicoglu SS, Ergun I, Ates A, Ozdag H, Akar N: Behcet's: A Disease or a Syndrome? Answer from an Expression Profiling Study. PLoS
One 11: e0149052, 2016
32. Okuzaki D, Yoshizaki K, Tanaka T, Hirano T, Fukushima K, Washio T, Nojima H: Microarray and whole-exome sequencing analysis of familial Behcet's disease patients. Sci Rep 6: 19456, 2016
33. Pawelec G: Immunosenescence: impact in the young as well as the old? Mech Ageing Dev 108: 1-7, 1999
34. Perez-Aso M, Montesinos MC, Mediero A, Wilder T, Schafer PH, Cronstein B: Apremilast, a novel phosphodiesterase 4 (PDE4) inhibitor, regulates inflammation through multiple cAMP downstream effectors. Arthritis Res Ther 17: 249, 2015
35. Puccetti A, Fiore PF, Pelosi A, Tinazzi E, Patuzzo G, Argentino G, Moretta F, Lunardi C, Dolcino M: Gene Expression Profiling in Behcet's Disease Indicates an Autoimmune Component in the Pathogenesis of the Disease and Opens New Avenues for Targeted Therapy.
J Immunol Res 2018: 4246965, 2018
36. Qin X, Xu J, Wu Z, Sun F, Chen H, Zheng W, Li S, Li P, Chen S, Shen M, Zhang W, You X, Wu Q, Zhang F, Li YZ: Association study of rs924080 and rs11209032 polymorphisms of IL23R-IL12RB2 in a Northern Chinese Han population with Behcet's disease. Hum Immunol 77: 1284-1290, 2016
37. Raker VK, Becker C, Steinbrink K: The cAMP Pathway as Therapeutic Target in Autoimmune and Inflammatory Diseases. Front Immunol 7: 123, 2016
38. Sakane T, Takeno M, Suzuki N, Inaba G: Behcet's disease. N Engl J Med 341: 1284-1291, 1999 39. Saurwein-Teissl M, Lung TL, Marx F, Gschosser C, Asch E, Blasko I, Parson W, Bock G, Schonitzer D, Trannoy E, Grubeck-Loebenstein B: Lack of Antibody Production Following Immunization in Old Age: Association with CD8+CD28- T Cell Clonal Expansions and an Imbalance in the Production of Th1 and Th2 Cytokines. The Journal of Immunology 168: 5893-5899, 2002
40. Schlegel N, Waschke J: cAMP with other signaling cues converges on Rac1 to stabilize the endothelial barrier- a signaling pathway compromised in inflammation. Cell Tissue Res 355: 587-596, 2014
41. Sousa I SF, Francisco D, Davatchi F, Abdollahi BS, Ghaderibarmi F, Nadji A, Mojarad Shafiee N, Xavier JM, Oliveira SA: Brief report: association of CCR1, KLRC4, IL12A-AS1, STAT4, and ERAP1 With Behçet's disease in Iranians. Arthritis Rheumatol. 67(10): 2742-2748., 2015 42. Takeuchi M, Kastner DL, Remmers EF: The immunogenetics of Behcet's disease: a
comprehensive review. J Autoimmun 64: 137-148, 2015
43. Takeuchi M, Mizuki N, Meguro A, Ombrello MJ, Kirino Y, Satorius C, Le J, Blake M, Erer B, Kawagoe T, Ustek D, Tugal-Tutkun I, Seyahi E, Ozyazgan Y, Sousa I, Davatchi F, Francisco V, Shahram F, Abdollahi BS, Nadji A, Shafiee NM, Ghaderibarmi F, Ohno S, Ueda A, Ishigatsubo Y, Gadina M, Oliveira SA, Gul A, Kastner DL, Remmers EF: Dense genotyping of immune-related loci implicates host responses to microbial exposure in Behcet's disease susceptibility.
Nat Genet 49: 438-443, 2017
44. Targonski PV, Jacobson RM, Poland GA: Immunosenescence: role and measurement in influenza vaccine response among the elderly. Vaccine 25: 3066-3069, 2007
45. Vallejo AN, Weyand CM, Goronzy JJ: T-cell senescence: a culprit of immune abnormalities in chronic inflammation and persistent infection. Trends Mol Med 10: 119-124, 2004
46. Wang H, Guo J, Jiang J, Wu W, Chang X, Zhou H, Li Z, Zhao J: New genes associated with rheumatoid arthritis identified by gene expression profiling. Int J Immunogenet 44: 107-113, 2017
47. Weiskopf D, Weinberger B, Grubeck-Loebenstein B: The aging of the immune system. Transpl
Int 22: 1041-1050, 2009
48. Weng NP, Akbar AN, Goronzy J: CD28(-) T cells: their role in the age-associated decline of immune function. Trends Immunol 30: 306-312, 2009
49. Yalcin B, Gur G, Artuz F, Alli N: Prevalence of metabolic syndrome in Behcet disease: a case-control study in Turkey. Am J Clin Dermatol 14: 421-425, 2013
50. Yang JY, Park MJ, Park S, Lee ES: Increased senescent CD8+ T cells in the peripheral blood mononuclear cells of Behcet's disease patients. Arch Dermatol Res 310: 127-138, 2018
51. Yang SH, Li CF, Chu PY, Ko HH, Chen LT, Chen WW, Han CH, Lung JH, Shih NY: Overexpression of regulator of G protein signaling 11 promotes cell migration and associates with advanced stages and aggressiveness of lung adenocarcinoma. Oncotarget 7: 31122-31136, 2016
52. Yu H, Zheng M, Zhang L, Li H, Zhu Y, Cheng L, Li L, Deng B, Kijlstra A, Yang P: Identification of susceptibility SNPs in IL10 and IL23R-IL12RB2 for Behcet's disease in Han Chinese. J Allergy Clin Immunol 139: 621-627, 2017
- 26 -
Ozoren N: Novel NLRP3/cryopyrin mutations and pro-inflammatory cytokine profiles in Behcet's syndrome patients. Int Immunol 26: 71-81, 2014
- 국문요약 -
CD8+CD27-CD28-와 CD8+CD27+CD28+ T세포 전사체 분석을 통한 베체트
병의 새로운 차별발현유전자와 신호전달체계 발견
아주대학교 대학원 의학과 김 소 민 (지도교수: 이 은 소) 연구배경: 베체트병은 점막과 피부, 혈관, 눈, 신경 및 기타 여러 주요 장기들을 침범할 수 있 는 임상증상을 가지는 만성 재발성 전신 질환이다. 면역노화는 면역계에 노화현상이 발생하며 나타나는 변화로, 감염성 질환에 취약해지고 백신 접종 시 면역획득에 실패하거나, 지속적인 만성 염증 증세를 보이는 것 등과 관련이 있다. 본 연구진의 선행 연구에서 베체트병 환자의 말초혈액에서 노화 CD8+ T세포의 빈도 증가를 확인한 바 있다. 연구목적: 따라서 본 연구에서는 베체트병 환자 군과 정상 대조군의 말초혈액 내 단핵세포를 노화(CD27-CD28-) 그리고 비노화(CD27+CD28+) CD8+ T세포 그룹으로 나눈 뒤, 전사 체 분석을 시행하여 이들의 유전자 발현 특성을 확인하고자 하였다. 연구방법: 18명의 베체트병 환자 군과 연령 일치된 정상대조군 18명을 대상으로 말초혈 단핵 세포를 분리하였다. CD8 마이크로비드(microbead)로 CD8+ T세포만 분리한 후 면역결합 단 세포 항체를 이용하여 CD8, CD27, CD28을 표지 하였고, Fluorescence-activated cell sorting(FACS)를 이용하여 노화 CD8+ T세포(CD8+CD27-CD28-)와 비노화 CD8+ T세 포(CD8+CD27+CD28+)로 분리하였다. 환자들로부터 분리된 세포들은 함께 모아 RPMI 1640 배지에 배양하였고 이후 anti-CD3으로 자극하였다. 자극된 세포들로부터 RNA isolation kit를 사용하여 RNA들을 추출하였고, 이를 Macrogen 사에 의뢰하여 전사체 분석을 시행하였다. 베체트병 환자와 정상대조군, 노화세포와 비노화세포 여부에 따른 네 가지 그룹으 로 나누어 차별발현유전자(Differentially expressed genes)를 분석하였다. 차별발현유전자들 은 Ingenuity Pathway Analysis(IPA)를 통해 분석하여 기능적 주석과 주요 생물학적 경로 및 상위조절인자를 확인하였다. 또한 본 전사체 분석의 유효성을 확인하기 위하여, 환자와 정 상인 간에 큰 폭의 차이를 보인 유전자들을 대상으로 환자와 정상대조군을 각 5명씩 추가 등 록하여 실시간 Polymerase chain reaction(PCR)을 시행하였다.연구결과: 노화세포 군에서 베체트병 환자의 차별발현유전자는 1103개였으며, 비노화세포 군 에서는 652개였다. 이 결과에 대한 유효성 평가를 위하여 시행한 상위 차별발현유전자들 중에
- 28 -
서 노화세포 군에서는 COL5A1, ARHGEF10, 그리고 LOC102724428이 전사체 분석의 결과 와 같은 경향성을 보였고 그 중 COL5A1에서는 환자와 정상대조군 사이에 통계적으로 유의한 차이를 확인할 수 있었다. 한편 비노화세포 군에서는 TRPV3, ARHGEF10, UBE2F-SCLY, CD302, SHANK1이 전사체 분석 결과와 같은 경향성을 보였고, 그 중 TRPV3과 ARHGEF10은 환자와 정상대조군 사이에 통계적으로 유의한 차이를 보였다. IPA를 통해 확인 된 주요 canonical 경로에서, 11개의 경로가 서로 반대의 활성을 보였으며 2개의 경로에서는 같은 방향의 활성을 보였다. 가장 주요한 canonical 경로로 확인된 것은 cAMP-매개 신호전 달 경로였다. 결론: 본 연구는 베체트병과 환자간의 차별발현유전자를 확인 시켜 주었다. 또한 IPA를 이용한 접근법은 베체트병의 병인기전과 관련되어 있을 가능성이 있는 경로들을 제시하였다. 본 연구 에서 확인된 차별발현유전자들을 이용하여 베체트병 환자 진단에 유용한 바이오마커(biomarker) 개발을 시행한다면, 아직 객관화 되지 않은 베체트병 진단을 보다 쉽고 간단하게 하며, 치료 반응성을 예측하는 데에도 도움이 될 것으로 기대한다. 핵심어: 베체트병, 전사체 분석, RNA 서열분석, CD8+ T 세포, 면역노화