저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학 석사학위 논문
미토콘드리아 형태 조절 단백질
OPA1과 연관된
새로운 비만 관련 유전자 발굴
아 주 대 학 교 대 학 원
의생명과학과 / 분자의학전공
박 슬 비
미토콘드리아 형태 조절 단백질
OPA1과 연관된
새로운 비만 관련 유전자 발굴
지도교수 정 선 용
이 논문을 이학 석사학위 논문으로 제출함.
2019년 8월
아 주 대 학 교 대 학 원
의생명과학과 / 분자의학전공
박 슬 비
i
– 국문요약 –
미토콘드리아 형태 조절 단백질 OPA1 과 연관된 새로운 비만
관련 유전자 발굴
미토콘드리아는 세포의 정상적인 생리적 기능의 유지에 필수적인 세포 소기관이다. 미토콘드리아는 세포의 생존, 사멸 및 대사 환경 변화에 대한 적응과 관련된 제어를 위해 융합 (fusion) 및 분열 (fission)을 통한 역동적인 형태 조절이 이루어진다. 이러한 미토콘드리아의 형태 조절은 Drp1, Fis1, Mfn1, Mfn2 그리고 Opa1 등의 단백질에 의해 이뤄지는 것으로 알려져 있으며, 최근 미토콘드리아 다이나믹스 (dynamics)가 비만, 인슐린 저항성, 제 2 형 당뇨병의 발병과의 관련이 점차 밝혀지고 있다. 본 연구는 비만 관련 표현형과 연관성이 있는 미토콘드리아 형태조절 단백질을 발굴하고 이와 관련된 새로운 유전자 발굴을 목적으로 하였다. 미토콘드리아 형태조절 관련 유전자 49 종을 선별하여 이들 유전자와 연관된 647 개의 SNP 을 선정하였다. Korean Association REsourece (KARE) 코호트의 임상자료, 검사 자료, 역학자료, SNP 유전형 자료를 이용하여 비만 관련 형질에 대한 연관성 분석 (association study)을 실시하였으며, 남성 복부지방 및 체지방에서 Bonferroni-correction threshold 유의수준을 통과하는 OPA1 유전자의 SNP rs7646539 (p = 0.00034)를 밝혔다. OPA1 유전자 상의 다수의 SNP 들도 남성의 복부지방 및 체지방과 유의한 연관성이 있었다.ii
3T3-L1 전지방 세포에서 Opa1 의 발현 억제 시에 대조군 대비 Oil Red O 염색 세포수가 감소하였으며, 지방세포 분화 마커인 Adipoq, Cebpa, Fabp4,
Slc2a4, Pparg 의 발현이 유의하게 감소되어 지방세포 분화가 현저하게
억제되었음을 증명하였다. Whole-genome RNA-sequencing 을 통해
Opa1 의 발현 억제에 따라 mRNA 발현에 현저한 차이를 보이는 유전자를 탐색 (screening)하여 기존에 비만 관련 형질과의 연관성이 보고되지 않은
Fmo2유전자 (flavin-containing monooxygenase 2)를 발굴하였다. Fmo2의 발현 억제 시에 지방세포 분화를 유도하였음에도 불구하고 대조군 대비 Oil Red O 염색 세포수가 감소하였으며, 지방세포 분화 마커인 Adipoq, Cebpa,
Fabp4, Slc2a4, Pparg 의 발현이 유의하게 감소되어 지방세포 분화가
현저하게 억제되었음을 증명하였다.
이러한 결과는 Fmo2 가 미토콘드리아 융합 조절 단백질인 Opa1 에 의해 매개되는 새로운 비만 관련 유전자일 가능성이 있음을 시사한다.
_________________________________________________________________
핵심어:
비만, 지방 분화, 미토콘드리아 형태 조절 단일염기 다형성(SNP), 연관성 분석, Optic atrophy protein 1 (Opa1), Flavin-containing monooxygenases 2 (Fmo2)iii
차 례
국문요약··· i
차례··· ⅲ
그림 차례··· vi
표 차례··· vii
I.
서론··· 1
II. 실험 재료 및 방법 ··· 6
A.
연관성 분석 (association study)··· 6
B.
세포주 및 세포 배양··· 7
C. Short hairpin RNA (shRNA) construct ··· 7
D.
지방세포 분화 유도··· 8
E. Oil Red O
염색··· 8
F. Whole-genome RNA-sequencing ··· 9
G. RNA
추출, cDNA
합성··· 9
iv
I. Western blot
분석··· 10
J.
통계분석··· 11
III.
결과··· 12
A.
한국인 코호트를 사용한 비만 관련 형질에 대한 연관성 분석 결과···12
B. Opa1
발현 억제 시의 미토콘드리아 형태 변화와 3T3-L1 세포의 지방세포 분화 변화··· 19
C.
전장 유전체RNA-sequencing
을 이용한 Opa1 매개 비만 관련 유전자 발굴···22
D. Opa1
발현 억제 세포에서의 지방세포 분화 유도에 따른Fmo2
의 발현 변화··· 25
E.
Fmo2 발현 억제에 따른 지방세포 분화 변화··· 27
IV.
고찰··· 30
V.
결론··· 33
참고문헌··· 34
v
vi
그림 차례
Figure 1. Schematic diagram of adipocyte differentiation ··· 4
Figure 2. Schematic diagram of mitochondrial fusion and fission machinery · 5
Figure 3. The association between OPA1 and obesity ··· 17
Figure 4. Alterations in adipogenesis in Opa1 knock-downed 3T3-L1 cells
··· 20
Figure 5. The expression levels of adipogenesis related genes and Fmo2 gene
obtained by whole-genome RNA-sequencing analysis in Opa1
knock-downed 3T3-L1 cells ··· 24
Figure 6. The expression levels of Fmo2 in Opa1 knock-downed 3T3-L1 and
C3H10T1/2 cells ··· 26
vii
표 차례
Table 1. Oligonucleotide primers used for quantitative RT-PCR ··· 10
Table 2. Basic characteristics of the study subjects ··· 13
Table 3. Information in the SNPs in the mitochondrial shaping genes used for
the association study ··· 14
Table 4. The SNPs in the OPA1 gene most significantly associated with obesity
related phenotypes in the Korean men ··· 16
Table 5. The most differentially expressed genes in 3T3-L1 cells after Opa1
- 1 -
I. 서론
세계보건기구 (WHO)는 인종이나 성별과 관계없이 체질량지수 25 kg/m² 이상을 과체중으로, 30 kg/m² 이상을 비만으로 정의하며 (2000), 국내외 연구 결과를 바탕으로, 대한비만학회와 세계보건기구 아시아태평양지역에서는 과체중의 기준을 체질량지수 23 kg/m² 이상, 비만의 기준은 체질량지수 25 kg/m² 이상으로 정의하고, 이때 체질량지수는 체중 (kg)을 신장 (meter)의 제곱으로 나눈 값으로, 비만도 판정의 기준이며, 체지방량과 상관관계가 높아 체중 및 신장을 이용한 지수 중 가장 널리 사용되는 방법이다. 비만은 비정상적이거나 과도한 체지방 축적을 초래하는 만성 질환으로 동맥경화, 제 2 형 당뇨병의 발병 위험이 높으며 심혈관 질환 등 다양한 대사증후군과 연관이 있다 (Kang et al., 2011). 비만은 과도한 음식물 섭취, 운동 부족 및 유전적 감수성으로 인해 주로 발생하며 (Yazdi et al., 2015), 약물, 내분비 장애 또는 정신 질환이 원인이 되기도 한다 (Bleich et al., 2008). 이러한 비만은 2016 년 국민건강영양조사 결과, 국내 만 30 세 이상의 인구 중 체질량지수 25 이상인 유병률이 37.0% (남자 50.0%, 여자 30.0%)이며, 유병자의 5 명 중 3 명은 체중감소를 시도한 적이 있는 것으로 나타났고, 우리나라 평균수명의 증가와 서구화된 식습관, 생활습관의 변화, 운동 부족, 스트레스 등의 환경적 위험요인들로 인해 향후 비만 유병률은 지속적으로 늘어날 것으로 예상된다. 특히, 가족력 등의 고위험군 유전적 소인을 가진 사람의 경우는 30 세 이전의 젊은 나이에 적절한 관리가 비만 예방을 위해 매우 중요하다. 이와 관련하여 비만 연관 유전자 (gene)/유전자 자리 (locus)가 많이 보고 되어있다. 하지만, 확실한 유전 소인으로 밝혀진- 2 - 유전자는 많지 않으며 아직 밝혀지지 않은 핵심 유전소인들이 많이 남아있고, 질환의 발병기전의 이해와 더불어, 위험도 예측 유전자 마커 및 새로운 치료 타겟의 발굴을 위해서는 체계적인 유전학적, 유전체학적 접근이 필요하다. 비만은 지방 생성 (adipogenesis)과 밀접한 관계가 있다. 지방 생성은 전지방세포가 성숙한 지방세포로 분화하는 것과 관련된 과정을 가리킨다 (Kersten, 2001). 지방 생성은 전사인자인 Peroxisome proliferator activated receptor gamma (Pparg), CCAAT/enhancer-binding protein α
(Cebpa)에 의해 시작되며 (Tontonoz et al., 1994), Solute carrier family 2
member 4 (Slc2a4), Adiponectin (Adipoq), Fatty acid binding protein 4
(Fabp4) 등의 지방세포 분화 마커를 이용하여 분화를 확인할 수 있다
(Figure 1)(Tontonoz and Spiegelman, 2008).
미토콘드리아는 세포 내에서 에너지 생산뿐만 아니라 다양한 역할을 하며 미토콘드리아의 형태와 관계가 밝혀지고 있다 (Chen et al., 2005; McBride et al., 2006; Detmer and Chan, 2007). 미토콘드리아의 형태 조절은 세포의 항상성 유지에 필수적이며, 특정 단백질들에 의해 이뤄지고 있는 것으로 알려져 있다 (Chan, 2006b; Cerveny et al., 2007; Hoppins et al., 2007).
미토콘드리아의 형태를 조절하는 단백질로는 융합을 조절하는 Mitofusin 1 (Mfn1), Mitofusin 2 (Mfn2), Optic atrophy protein 1 (Opa1)과 분열을 조절하는 Dynamin related protein 1 (Drp1), Fission 1 (Fis1) 등이 알려져 있다. Mfn1 과 Mfn2 는 미토콘드리아의 외막에 존재하며, Opa1 은 미토콘드리아의 내막에 위치하는 단백질 (Griparic et al., 2004)로 Mfn1 에 의존적이다(Chan, 2006b). Drp1 은 대부분 세포질에 존재하며, Fis1 과 같은, 미토콘드리아의 외막에 위치하는 수용체 단백질과의 상호작용을 통해
- 3 -
미토콘드리아 외막으로 이동하여 미토콘드리아의 분열을 유도한다 (Figure 2) (Chan, 2006b).
최근의 연구결과에 따르면 미토콘드리아의 형태 조절은 apoptosis (Perfettini et al., 2005; Youle and Karbowski, 2005), autophagy (Twig et al., 2008), calcium signaling (Szabadkai et al., 2006), neurodegeneration (Chen et al., 2007; Cheung et al., 2007; Baloh, 2008), ageing (Lee et al., 2007; Scheckhuber et al., 2007; Tatsuta and Langer, 2008), development (Chen et al., 2003; Chan, 2006a), reactive oxygen species (Yu et al., 2006), oncogenesis (Alirol and Martinou, 2006), adipogenesis (Li et al., 2017; Lopez-Lluch, 2017) 등의 다양한 생명활동에 관여함을 알 수 있다. 특히, 12 주간 고지방 식이요법으로 비만을 유도한 rat 의 백색 지방 조직에서
Opa1의 발현이 증가했다는 연구 결과는 (Lahera et al., 2017) 미토콘드리아 역학과 비만의 연관성을 증명한다. 따라서, 미토콘드리아 형태 조절의 기전에 대한 연구와 세포 기능과의 상관관계에 대한 연구는 새로운 연구영역으로 주목받고 있다.
Fmo2 의 정확한 역할은 잘 알려져 있지 않지만, 다양한 산화 환원 스트레스 인자에 의해 유도되며 소포체의 혈장 항상성에 영향을 주는 환원 스트레스의 저항성에 중요하며, Hypoxia induce factor 1 (HIF-1)의 표적이라는 연구결과가 있다 (Suh et al., 1999; Suh and Robertus, 2000; Leiser et al., 2015).
본 연구에서는 OPA1 유전자의 발현 변화에 따른 비만 관련 형질과의 연관성을 조사하고, OPA1 매개의 새로운 비만 관련 인자를 발굴하는 것을 목표로 하였다.
- 4 -
(Ghaben and Scherer, 2019)
Fi
g
u
r
e
1
. S
c
h
e
ma
ti
c
d
ia
g
r
a
m
o
f
a
d
ip
o
cy
te
d
if
fe
re
n
ti
a
ti
o
n
.
M
ul
ti
p
o
te
nt
f
ib
ro
b
la
st
-l
ik
e
m
es
en
ch
y
m
al
p
re
cu
rs
o
rs
c
an
d
if
fe
re
n
ti
at
e
to
m
y
o
b
la
st
,
ch
o
n
d
ro
c
y
te
,
o
st
eo
b
la
st
a
n
d
a
d
ip
o
c
y
te
.
B
o
n
e
mo
rp
h
o
ge
n
et
ic
p
ro
te
in
(B
M
P
)
si
gn
al
in
g
is
a
co
mm
itm
en
t
to
m
ed
ia
te
th
es
e
m
e
se
n
c
h
y
m
al
p
re
c
u
rs
o
rs
to
t
h
e
ad
ipo
c
y
te
.
W
h
e
n
th
e
c
o
m
m
it
te
d
p
re
ad
ip
oc
yt
e’
s
gr
ow
th
a
rr
es
t,
th
e
m
ast
er
re
gu
la
to
r
P
pa
rγ
a
n
d
t
ra
n
sc
ri
pt
ion
c
o
-a
ct
iv
at
o
rs
C
eb
pα
a
n
d
C
eb
pβ
i
s
ac
ti
v
at
ed
.
L
ip
id
a
cc
u
m
ul
at
io
n
d
er
iv
es
th
e
e
x
p
re
ss
io
n
i
ns
u
li
n
-s
en
si
ti
v
e
tr
an
sp
o
rt
er
S
lc
2
a
4
(
G
lut
4
),
i
t
is
m
ar
k
er
o
f
ea
rl
y
s
ta
ge
s
ad
ip
o
c
y
te
s.
A
n
d
w
h
e
n
di
ff
e
re
n
ti
a
ti
o
n
e
n
d
,
m
at
u
re
a
di
p
o
c
y
te
r
e
le
a
se
t
h
e
p
e
p
ti
d
e
h
o
rm
o
n
e
A
d
ipo
q
,
as
so
ci
at
ed
w
it
h
ac
cu
m
u
la
te
f
at
ty
a
c
id
bi
n
d
in
g
p
ro
te
in
F
a
b
p
4
a
nd
li
p
id
-d
ro
pl
et
-a
ss
o
ci
at
ed
p
ro
te
in
P
lin
2
(
G
h
ab
en
a
n
d
S
ch
er
er
,
2
0
1
9
).
- 5 -
Figure 2. Schematic diagram of mitochondrial fusion and fission machinery.
Fusion of mitochondrial outer membrane is occurred by Mfn1 and Mfn2. Fusion
of inner membrane is regulated by interaction of Opa1 and cardiolipin. For
mitochondrial fission, Drp1 is recruited to mitochondria through interaction with
receptor proteins that located in the outer membrane, like Fis1. And then Drp1 is
oligomerized to filaments that wrap around the mitochondria. Finally, constriction
of Drp1 spirals derived mitochondrial fission (Kameoka et al., 2018).
- 6 -
II. 실험 재료 및 방법
A. 연관성 분석 (association study)
Korean Association REsourece (KARE) study 연구의 대상자는 다른 연구 (Cho et al., 2009)에 의해 보다 자세히 기술되었다. Genotyping 결과가 불량하거나 약물 치료를 받은 피험자는 제외되었고, 결과적으로 3,027 명의 한국 남성들을 조사하였다.
피험자의 체지방률과 골밀도를 Table 3 에 나타내었으며, 피험자 중 체지방률과 골다공증을 측정한 연관성을 분석하였다. 체지방률은 4 극 생체 전기 임피던스 분석 (tetrapolar bioelectrical impedance analysis) (Inbody 3.0, Biospace, Korea)으로 측정하였다. 생체 전기 임피던스 분석은 이전에 검증된 경험적으로 유도된 공식을 사용하여 두가지 매개 변수인 지방과 마른 조직을 측정한다. 골밀도는 골다공증을 선별하는데 널리 사용되는 골절 저항인 뼈 강도의 프록시 측정으로, 골밀도는 건강한 젊은 성인 인구의 평균 SOS (speed of sound)와 측정된 SOS 의 차이를 젊은 성인 인구의 SOS 표준 편차로 나누어 T score 로 추정하였으며, SOS 는 피험자의 요골 원위부와 중축 경골에서 정량적 초음파로 측정하였다. 골다공증은 골밀도 T score -2.5 SD 이하, 정상은 골밀도 T score -1 SD 이상으로 정의하였다.
자세한 genotyping 과 품질 관리 과정은 Cho et al. (Cho et al., 2009)에 보고되어 있다. 대부분의 DNA 샘플을 참가자의 말초 혈액으로부터 분리하고 Affymetix Genome-Wide Human SNP array 5.0 (Affymetrix, Inc., Santa Clara, CA, USA)을 사용하여 유전자형을 결정하였으며, Genotyping 의 정확도는 Bayesian Robust Linear Model with Mahalanobis distance
- 7 -
classifier (BRLMM) 알고리즘 (Rabbee and Speed, 2006)을 사용하여 계산하였다. 샘플의 genotyping 정확도는 98% 미만이었으며, 소실된 genotype 호출률 (missing genotype call rate)이 높거나 (≥4%), 이형 접합률 (heterozygosity)이 높은 (>30%) 표본은 제외되었다.
NCBI human genome build 36 에 따라 OPA1 유전자의 4 가지 SNP (Table 4)을 선별하였다. 또한, 이 연구는 국립보건연구원의 기관 검토위원회의 승인을 받았고, 모든 피험자로부터 서면동의서를 받았으며, 사용한 임상 정보 및 유전자형 데이터는 국립보건연구원에서 제공하였다.
B. 세포주 및 세포 배양
전지방세포 3T3-L1 을 10% bovine calf serum (BCS) (Gibco, NZ), 1% Antibiotic-Antimycotic (Gibco Life Sciences, Grand Island, NY, USA)을 첨가한 high-glucose Dulbecco’s Modified Eagle’s medium (DMEM) (Gibco)으로, 마우스 중간엽 세포 C3H10T1/2 를 10% fetal bovine serum (FBS) (Gibco, NY, USA), 1% Antibiotic-Antimycotic (Gibco)을 첨가한 low-glucose DMEM (Gibco)으로, 인간 배아 신장 세포 HEK293TN 은 10% FBS (PANBIOTECH), 1% Antibiotic-Antimycotic (Gibco)을 첨가한 high-glucose DMEM 으로 5% CO2, 37℃의 환경에서 배양하였다.
C. Short hairpin RNA (shRNA) construct
Opa1 유전자에 대한 shRNA 와 Fmo2 유전자에 대한 shRNA 가 삽입되어 있는 pLKO1.0 (Sigma Aldrich, USA)들을 구매한 후, HEK293TN 세포에 Lipofectamine™ 3000 Transfection Kit (Invitrogen, CA, USA)와
- 8 -
pPACKH1 HIV Lentivector Packaging Kit (SBI, USA)를 이용하여 Lentiviral particle 제작한 후 3T3-L1, C3H10T1/2 에 infection 하고, selection 을 위해 puromycin (3T3-L1: 2 ㎍/mL, C3H10T1/2: 4 ㎍/mL) 처리하였다.
D. 지방세포 분화 유도
3T3-L1 전지방 세포의 지방세포로의 분화는 0.5 mM 3-isobutyl-1-methylxanthine, 1 mM dexamethasone 및 1 ㎍/mL insulin 으로 처리함으로써 유도하였으며 3 일 후, 1 ㎍/mL insulin 을 포함하는 media 로 배양하였다. 이때 insulin 만 들어있는 배지는 3 일마다 교체하였고, 총 8 일간 배양하였다.
C3H10T1/2 중간엽 세포의 지방세포로의 분화는 MesenCult™ Adipogenic Differentiation Kit (Mouse) (STEMCELL, Vancouver, Canada)를 이용하여 14 일동안 배양하였으며, 배지는 3 일마다 교체하였다.
E. Oil Red O 염색
지방 생성 유도 후 지질의 축적은 Oil Red O 염색으로 측정하였다. 분화된 지방세포를 PBS 로 세척하고, 차가운 4% paraformaldehyde 를 분주한 후, 10 분간 방치하여 고정한 뒤 증류수로 3 번 헹구어 실온에서 air dry 하고, 100% propylene glycol 을 분주하여 2~5 분 방치한 뒤, 60℃로 예열된 Oil Red O 용액을 분주하여 20 분동안 heat block 위에 둔다. 85% propylene glycol 용액을 첨가하여 3 분 뒤 증류수로 2 번 헹군 뒤 대조염색으로 hamatoxylin 을 30 초 염색한다. 증류수로 워싱 후 현미경으로 확인하였다.- 9 -
F. Whole-genome RNA sequencing
라이브러리는 TruSeq RNA Sample Preparation Kit (Illumina, CA, USA)를 사용하여 100 bp 의 paired-end sequencing 을 위해 준비되었다. mRNA 는 oligo (dT) 자성 비드를 사용하여 RNA 2 ㎍에서 정제 및 단편화하였으며, mRNA 는 random hexamer priming 을 통해 single-stranded cDNA 로 합성되어 double-single-stranded cDNA 를 제조하기 위한 두 번째 가닥 합성을 위한 주형으로 사용되었다. End repair, A-tailing, adapt ligation 의 순차적 과정 후에, cDNA 라이브러리는 PCR (중합 효소 연쇄 반응)을 사용하여 증폭되었다. 이 cDNA 라이브러리의 quality 는 Agilent 2100 BioAnalyzer (Agilent, CA, USA)로 평가했다. KAPA 라이브러리 정량 키트 (Kapa Biosystems, MA, USA)를 사용하여 제조업체의 라이브러리 정량화 프로토콜에 따라 정량화하였다. 변성된 템플릿의 클러스터 증폭 후, 시퀀싱은 Illumina HiSeq2500 (Illumina, CA, USA)을 사용하여 쌍방향 (2 × 100 bp)으로 진행되었다.
G. RNA 추출 및 cDNA 합성
배양한 세포에서 Trizol™ reagent (Invitrogen, USA)와 RNeasy mini kit (Qiagen, Germany)를 이용하여 RNA 를 추출하고, 260 nm, 280 nm 에서 흡광도의 비율을 사용하여 추출된 RNA 의 quality 를 측정하였다. 그 후, 1000 ng 의 RNA 를 2 U RNase-free DNase I (Invitrogen, USA)으로 처리한 후 oligo (dT) 18 primer, random primer 와 RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA)를 사용하여 cDNA 를 합성하였다.
- 10 -
H. 정량 real-time PCR (qRT-PCR)
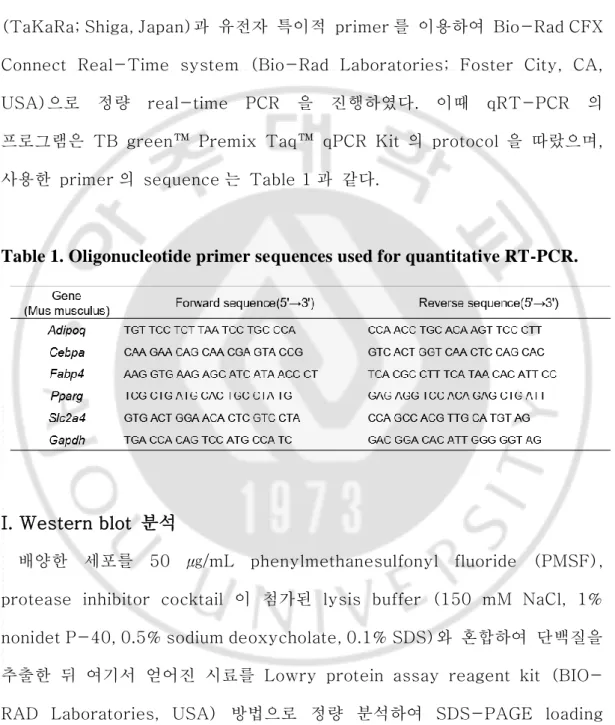
100 ng 의 cDNA 를 주형으로, TB green™ Premix Taq™ qPCR Kit (TaKaRa; Shiga, Japan)과 유전자 특이적 primer 를 이용하여 Bio-Rad CFX Connect Real-Time system (Bio-Rad Laboratories; Foster City, CA, USA)으로 정량 real-time PCR 을 진행하였다. 이때 qRT-PCR 의 프로그램은 TB green™ Premix Taq™ qPCR Kit 의 protocol 을 따랐으며, 사용한 primer 의 sequence 는 Table 1 과 같다.
Table 1. Oligonucleotide primer sequences used for quantitative RT-PCR.
I. Western blot 분석
배양한 세포를 50 ㎍/mL phenylmethanesulfonyl fluoride (PMSF), protease inhibitor cocktail 이 첨가된 lysis buffer (150 mM NaCl, 1% nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS)와 혼합하여 단백질을 추출한 뒤 여기서 얻어진 시료를 Lowry protein assay reagent kit (BIO-RAD Laboratories, USA) 방법으로 정량 분석하여 SDS-PAGE loading buffer (5X)와 섞은 후, 100℃에서 10 분간 가열하였다. 이를 8~10% SDS-PAGE gel 로 전기영동하여 분리하고, polyvinylidene fluoride (PVDF,
- 11 -
Immobilon-P membrane (0.45 μm), Millipore Corporation, USA)에 흡착 이동시켜, 5% BSA 혹은 skim milk 에 희석한 각 유전자의 특이적 항체로 Western blot 을 하여 분석하였다.
사용한 1 차 항체는 Opa1 (BD Transduction Laboratories, USA), Fmo2 (Proteintech), Actin (SANTA CRUZ BIOTECHNOLOGY), 2 차 항체는 peroxidase 가 conjugation 된 goat anti-rabbit, goat anti-mouse, rabbit anti-goat (Bethyl)를 이용하여 반응시킨 뒤, West-Q pico-dura ECL solution (GenDEPOT)을 처리하여 Agfa X-ray film (Agfa)에 현상하였다
J. 통계분석
Association study 의 대부분의 통계 분석은 PLINK version 1.07 (http://pngu.mgh.harvard.edu/~purcell/plink) and PASW Statistics version 17.0 (SPSS Inc.; Chicago, IL, USA)를 사용하였다. Haploview Version 4.2 program (Whitehead Institute for Biomedical Research; Cambridge, MA, USA)를 이용하였고, 대부분의 연관성 검사는 additive genetic model 을 기반으로 하였으며, p-value threshold 는 p = 0.0004 로 조정되었다. Plot 의 경우, 웹 기반 plotting 도구인 LocusZoom 버전 1.1 (http://csg.sph.umich. edu/locuszoom)을 사용하였다 (Pruim et al., 2010).
실험적 연구에서, 결과는 평균 ± 표준 편차로 표현되었으며 모든 실험은 최소 3 번 반복하였다. 통계 분석은 PASW Statistics 버전 17.0 (SPSS Inc.)을 사용하였고, 그룹 간의 통계적 유의성은 Student 's t-test 에 의해 계산하였다. p < 0.05 미만의 확률 값은 통계적으로 유의하다고 간주하였다.
- 12 -
III. 결과
A. 한국인 코호트를 사용한 비만 관련 형질에 대한 연관성 분석 결과
본 연구에서는 안성과 안산 지역의 주민을 대상으로 구축된 총 3,747 명의 남성 유전체 역학 코호트 (KARE cohort)을 사용하였다 (Table 2). 지금까지 알려진 49 종의 미토콘드리아 형태 조절 유전자들에 존재하는 647 개의 SNP 들을 분석 대상으로 하여 (Table 3), 비만 연관 표현형들에 대한 연관성 분석 (association study)을 시행하였다. 분석한 전체 SNP 숫자를 계산한 Bonferroni-correction threshold 유의수준 (p < 0.0004)을 통과하는 SNP 는 남성 복부비만율과 연관성이 있는 OPA1 유전자의 rs7646539 SNP 은 1 개였다 (Table 4). OPA1 유전자에 있는 SNP 들에 대한 signal plot 결과에서 rs7646539 SNP 과 연관되어 있는 다수의 SNP 들이 남성 복부지방율과 남성 체지방율에서 유의한 연관성이 있음을 알 수 있었다 (Figure 3).
- 13 -
- 14 -
Table 3. Information in the SNPs in the mitochondrial shaping genes used
for the association study.
- 15 -
Abbreviation: Chr., chromosome. *The SNPs were selected from the KARE data based on their locations within the gene boundary (20 kb upstream and downstream of the first and last exons, respectively) according to NCBI human genome build 36.
- 16 -
T
a
b
le
4
.
T
h
e
S
N
Ps
in
t
h
e
O
P
A
1
g
en
e
m
o
st
si
g
n
if
ic
an
tl
y
a
ss
o
ci
a
te
d
w
it
h
ob
es
it
y
r
ela
te
d
p
h
en
o
typ
es
in
th
e
Ko
re
a
n
m
en
.
*R equi red sa m pl e s iz e for a ss oc iat ion bel ow t he st udi ed sub jec t s iz e a re ind ica ted in i ta li c a nd b ol d. The p -v al ues be low t he B onf er ron i-cor rec ted si g ni fi ca n ce l ev el ( p < 0.0 004) i s indi ca ted in bo ld an d unde rl in ed, a nd t h e p -v al ues b el o w t he s tand ar d si g ni fi ca n ce l ev el ( p < 0. 05 ) a re in di ca te d i n b ol d. T he sa m pl e si ze f or 80 % p owe r at α= 0. 05 is b ase d o n K AR E p ar am et er s, inc ludi ng m inor a ll el e f req uenc y , e ff ec t s iz e a nd m ea n v al ue of t he pheno ty pes . A bbr ev ia ti on s: A 1, m inor a ll el e; A 2, m aj or al lel e; B MI , B ody m as s i nd ex; WHR , Wa is t hi p r at io; B FR , B ody f at r at e; A bFR , A bdom inal f at r at e.- 18 -
Figure 3. The association between OPA1 and obesity.
The p-values of SNPs in the OPA1 gene determined by association analysis with
abdominal fat rate (A) and body fat rate (B) in Korean men. The p-value (Y-axis)
and chromosome position (X-axis) of SNPs are plotted. The selected SNPs are
labeled with arrows. The other SNPs are colored according to the heat scale, based
on the r
2values, to reflect their linkage disequilibrium (LD) with the top above
SNP. Lower panel; gene annotations are described. The direction of the transcripts
is shown using arrows.
- 19 -
B. Opa1 발현 억제 시의 미토콘드리아 형태 변화와
3T3-L1 세포의지방세포 분화 변화
3T3-L1 전지방 세포주에서 Opa1 유전자 발현 억제 후 지방세포 분화를 유도한 뒤, Western blot 과 qRT-PCR 로 Opa1의 발현을 확인한 결과, Opa1
발현을 억제한 shOpa1_#1 과 shOpa1_#2 세포에서 Opa1 의 발현이 현저히 감소하여 Opa1 유전자가 효과적으로 억제된 것을 확인하였다 (Figure 4A).
Mito-tracker 를 이용하여 미토콘드리아의 형태를 관찰하였을 때, shOpa1_#1 과 shOpa1_#2 세포는 Opa1 발현 억제로 미토콘드리아의 길이가 현저히 짧아진 것을 확인하였으며, 지방세포 분화 시 생성되는 지질 방울을 붉게 염색하는 Oil Red O 염색 결과에서 shOpa1_#1 과 shOpa1_#2 는 대조군 (shCon) 세포와 비교하였을 때 염색정도가 현저히 감소된 것이 관찰되었다 (Figure 4B).
지방세포 분화 유도 시에 양성 대조군 (shCon)에 비해 shOpa1_#1 과 shOpa1_#2 세포에서 지방세포 분화 마커인 Adipoq (p < 0.05), Cebpa (p < 0.05), Fabp4 (p < 0.05), Slc2a4 (p < 0.05), Pparg (p < 0.05)의 발현이 유의하게 감소되었다 (Figure 4C).
- 21 -
Figure 4. Alterations of adipogenesis in Opa1 knock-downed 3T3-L1 cells.
(A) Evaluation of Opa1 short hairpin RNA interference constructs. 3T3-L1 cells
were transfected with the control vector (shCon) or 3 kinds of Opa1 short hairpin
constructs (shOpa1). After culturing for 3 days, Opa1 protein and mRNA levels
were assessed by Western blot analysis and quantitative RT-PCR, respectively.
(B, C) 3T3-L1 cells were treated with 0.5 mM of 3-isobutyl-1-methylxanthine, 1
mM of dexamethasone, and 1 µg/mL of insulin for 8 days. Mitochondria stained
with MitoTracker and nuclei stained with DAPI were imaged by confocal
microscopy (B). Cells stained with Oil Red O were imaged by inverted
microscopy (B). The mRNA levels of Adipoq, Cebpa, Fabp4, Slc2a4, and Pparg
genes were measured by quantitative RT-PCR (C). ***p < 0.001 vs. shCon.
- 22 -
C.
전장 유전체 RNA-sequencing 을 이용한Opa1
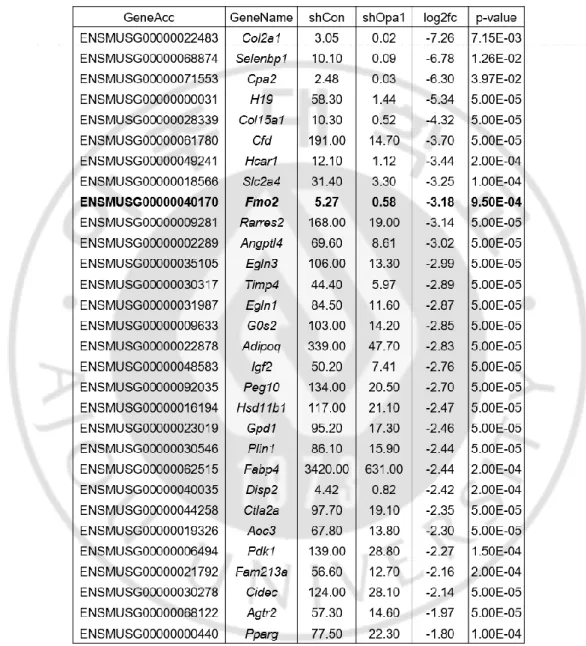
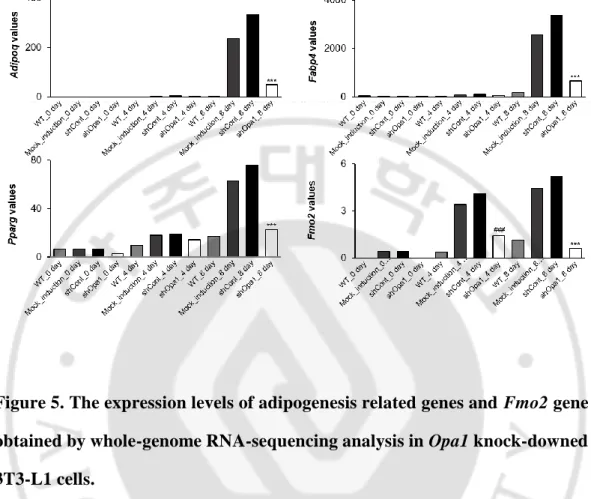
매개 비만 관련 유전자 발굴다음으로, 미토콘드리아 형태조절 단백질 OPA1 이 매개하는 새로운 비만인자를 밝히고자 하였다. RNA 간섭 (RNA interference)에 의해
Opa1 의 발현이 억제된 3T3-L1 전지방 세포에 지방세포 분화를 유도한 후 Opa1 의 발현과 지방세포 분화로 인해 변화되는 유전자를 확인하기 위해 whole-genome RNA-sequencing 을 시행하였다. Opa1 유전자 발현 억제 시에 유전자 발현에 큰 차이가 있는 (log2fold 값이 큰) 대표적 30 개의 유전자들 중 이미 비만과의 관련성이 보고된 유전자들을 제외하였을 때, 지방세포 분화 유도 시 증가하고 Opa1 발현 억제된 분화 세포에서는 감소하는 패턴을 보인 Fmo2 유전자를 비만 관련 후보유전자로 선정하였다 (Table 5). 3T3-L1 세포에 지방세포 분화 유도 시, 지방세포 분화 마커로 알려진
Slc2a4, Adipoq, Fabp4, Pparg 유전자가 유의하게 증가하였고, Opa1 발현
억제 세포에서는 감소하였기 때문에, 실험 모델로서 적합함을 확인하였다 (Figure 5).
Opa1 유전자 발현이 억제된 세포의 지방 분화 4 일차, 8 일차에 Fmo2
유전자의 발현이 양성 대조군 세포 (shCon)에 비해 현저하게 감소되었음을 밝혔다 (Figure 5).
- 23 -
Table 5. The most differentially expressed genes in 3T3-L1 cells after Opa1
gene knock down.
Abbreviations: GeneAcc, gene accession number; shCon, short hairpin control (RNA interference); shOpa1, short hairpin Opa1 gene (RNA interference); and Log2fc, logarithm of fold change.
- 24 -
Figure 5. The expression levels of adipogenesis related genes and Fmo2 gene
obtained by whole-genome RNA-sequencing analysis in Opa1 knock-downed
3T3-L1 cells.
Untransfected 3T3-L1 cells, and 3T3-L1 cells transfected with the control vector (shCon), or transfected with Opa1 short hairpin constructs (shOpa1) were treated with 0.5 mM of 3-isobutyl-1-methylxanthine, 1 mM of dexamethasone, and 1 µg/mL of insulin for 8 days. Total RNA was extracted, the cDNA was synthesized, and then whole-genome RNA-sequencing analysis was conducted. The mRNA levels of Adipoq, Cebpa, Fabp4, Slc2a4, and Pparg genes were analyzed from the RNA-sequencing data. ***p < 0.001 vs. shCont on the same day.
- 25 -
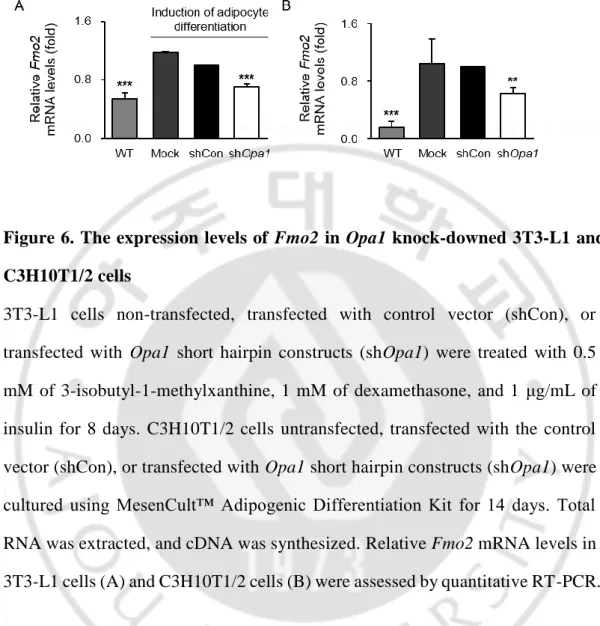
D. Opa1 발현 억제세포에서의 지방세포 분화 유도에 따른 Fmo2 의
발현 변화
Opa1 유전자를 발현 억제한 후, 지방세포로 분화를 유도한 3T3-L1 전지방 세포와 C3H10T1/2 중간엽 세포에서 Fmo2 의 발현을 확인하였다. 음성 대조군 (WT)에 지방세포 분화를 유도하였을 때 Fmo2 의 발현이 증가하고, 양성 대조군 (shCon)에 비해 Opa1 발현 억제군 (shOpa1)에서- 26 -
Figure 6. The expression levels of Fmo2 in Opa1 knock-downed 3T3-L1 and
C3H10T1/2 cells
3T3-L1 cells non-transfected, transfected with control vector (shCon), or
transfected with Opa1 short hairpin constructs (shOpa1) were treated with 0.5
mM of 3-isobutyl-1-methylxanthine, 1 mM of dexamethasone, and 1 μg/mL of
insulin for 8 days. C3H10T1/2 cells untransfected, transfected with the control
vector (shCon), or transfected with Opa1 short hairpin constructs (shOpa1) were
cultured using MesenCult™ Adipogenic Differentiation Kit for 14 days. Total
RNA was extracted, and cDNA was synthesized. Relative Fmo2 mRNA levels in
3T3-L1 cells (A) and C3H10T1/2 cells (B) were assessed by quantitative RT-PCR.
- 27 -
E. Fmo2 발현 저해에 따른 지방세포 분화 변화
Fmo2 발현량에 따른 지방세포 분화의 변화를 확인하기 위해, Fmo2
유전자의 발현을 억제한 후, 지방세포로 분화를 유도하였다. Western blot 과 qRT-PCR 을 이용하여 Fmo2 의 발현이 확실하게 억제되었음을 확인하였다 (Figure 7A). Oil Red O 염색 결과를 통해 Fmo2 발현 억제 후 지방세포 분화를 유도했을 시 지방세포의 분화가 현저히 감소하였음을 증명하였다 (Figure 7B). 또한, Fmo2 발현 저해 세포에서 지방세포 분화 관련 마커인Adipoq (p < 0.05), Cebpa (p < 0.05), Fabp4 (p < 0.05), Pparg (p < 0.05)의 발현이 감소하였다 (Figure 7C).
- 29 -
Figure 7. Alterations in adipogenesis in Fmo2 knock-downed 3T3-L1 cells.
(A) Evaluation of Fmo2 short hairpin RNA interference constructs. 3T3-L1 cells were transfected with the control vector (shCon) or Fmo2 short hairpin construct (shFmo2). After culturing for 3 days, Fmo2 protein and mRNA levels were assessed by Western blot analysis and quantitative RT-PCR, respectively. (B, C) 3T3-L1 cells were treated with 0.5 mM of 3-isobutyl-1-methylxanthine, 1 mM of dexamethasone, and 1 µg/mL of insulin for 8 days. Cells stained with Oil Red O were imaged by inverted microscopy (B). The mRNA levels of Adipoq, Cebpa, Fabp4, Slc2a4, and Pparg genes were measured by quantitative RT-PCR (C). **p <0.005, ***p <0.01 vs. shCon.
- 30 -
IV. 고찰
비만은 전세계적으로 유병률이 증가하고 있는 위험한 질병 중 하나임에도 불구하고 확실한 유전적 소인으로 발견된 유전자는 많지 않은 실정이다. PubMed 에서 obesity 관련 논문 검색 시 총 278,324 편이 검색되며, 10 년 단위의 조사에 의하면 2000 년대 이후부터 현재까지 급속적으로 논문편수가 증가되었음을 알 수 있다. Mitochondrial dynamics 를 동시에 입력하여 검색된 논문은 98 편에 불과하지만, 이중 94 편이 2010 년 이후에 publish 된 논문이었으며, 이러한 통계는 비만과 미토콘드리아 역학에 대한 관련성이 최근 입증되고 주목받고 있음을 나타낸다. 그럼에도 불구하고 아직까지 확실한 유전적 소인으로 발견된 유전자는 많지 않아 체계적인 유전학적, 유전체학적 접근이 필요하다. 미토콘드리아의 생합성은 지방세포 분화 과정에서 상향 조절되며, 지방 생성을 촉진한다 (Tormos et al., 2011; Zhang et al., 2013). 이때, 미토콘드리아의 밀도는 증가하고, 구조 및 생화학적 변화가 나타난다 (Hofmann et al., 2012). 당뇨, 비만 및 심혈관 질환 등의 대사질환에서 지방세포의 미토콘드리아 기능이 저하되는 것이 밝혀졌으며 (De Pauw et al., 2009; Vernochet et al., 2014), 지방세포의 분화 및 항상성에 역할을 하는 미토콘드리아의 기능 저하는 심각한 결과를 초래한다 (Tormos et al., 2011; Zhang et al., 2013). 이러한 연구 결과는 지방세포의 미토콘드리아 기능과 관련된 단백질이 대사 질환의 잠재적인 치료 표적임을 시사한다 (Cho et al., 2019).- 31 -
미토콘드리아의 융합을 조절하는 것으로 알려진 OPA1 은 2011 년의 연구 결과에 따르면 지방세포의 지질 방울(lipid droplets)에 위치하는 것으로 밝혀졌다. OPA1은 백색 지방세포의 분화 과정에서 증가하는데 (Pidoux et al., 2011), 이것은 지질의 축적과 OPA1 의 발현이 동시에 일어남을 의미한다 (Chu et al., 2017).
본 연구에서는 Candidate gene association study 로 미토콘드리아 형태 조절 단백질 OPA1 과 비만의 연관성을 증명하였다. 실제 KARE cohort 는 여성과 남성 모두를 대상으로 피험자를 모집했지만, 여성을 포함한 결과는 유의적인 결과가 없었다. 이것은 모집된 여성의 연령이 갱년기에 가깝고, 갱년기의 여성은 호르몬의 급격한 변화가 일어나는 시기이기 때문일 것으로 예상된다.
지방세포 분화 시에 발현이 증가하는 것으로 알려진, 지방세포 분화의 마커
Pparg, Cebpa, Slc2a4, Adipoq, Fabp4, Plin2 (Takahashi et al., 2016; Smith
et al., 2018; Ando et al., 2019) 모두 Opa1 의 발현을 억제시킨 경우와
Fmo2의 발현을 억제시킨 경우에 유의적으로 감소하였다. Oil Red O 의 결과 또한 지방세포의 분화가 감소하는 것으로 보아 Opa1과 Fmo2 모두 지방세포 분화시에 중요한 역할을 하는 것을 알 수 있다.
Fmo2 는 열량 제한적 식이요법을 진행한 마우스 모델에서의 수명 연장에 유의미하게 높은 수준을 보인 유전자로 (Swindell, 2009), 기아와 유사한 반응과 연관되어 유전자의 발현이 증가하고, 미토콘드리아의 항상성과 수명 연장에 역할을 한다는 연구 결과가 있다 (Bennett et al., 2017). 또한, family 유전자인 Flavin-containing monooxygenase 3 (Fmo3)는 지질 및 포도당 내성의 주요 조절 인자로 알려져 있으며 (Morandi et al., 2018), BMI 지수
- 32 -
및 비만과 양의 상관관계 (Schugar et al., 2017)라는 연구 결과가 있다. Flavin-containing monooxygenase 5 (Fmo5) 또한 체중 조절과 포도당 처리 및 인슐린 감수성과 연관되어, 비만 및 인슐린 저항성의 치료제로서의 가능성 (Scott et al., 2017)이 밝혀져 있다. 본 연구에서는 Opa1 과 Fmo2 의 발현량 변화가 지방세포 분화에 관여한다는 것을 밝혔다. 향후 Opa1 이외의 다양한 미토콘드리아 형태조절 단백질에 대한 연구를 통해 미토콘드리아 형태 다이나믹스에 따른 Fmo2 유전자의 발현 및 활성의 변화가 비만 관련 표현형에 미치는 영향을 밝힐 수 있을 것으로 기대된다.
- 33 -
V. 결론
Korean Association REsourece (KARE) 코호트를 이용하여 49 종의 미토콘드리아 형태조절 관련 유전자의 647 개 SNP 에 대한 연관성 분석 (association study)을 통해 남성 복부비만율과 Bonferroni-correction threshold 유의수준을 통과하는 OPA1 유전자의 rs7646539 SNP (p < 0.00034)를 찾았다. 3T3-L1 전지방 세포에서 Opa1 의 발현 억제 시에 지방세포 분화가 유의하게 억제되었으며, whole-genome RNA-sequencing 을 통해 Opa1의 발현 억제에 따라 발현에 현저한 차이를 보이고 기존에 비만 관련 형질과의 연관성이 보고되지 않은 Fmo2 유전자를 발굴하였다. Fmo2 의 발현을 억제할 경우 지방세포 분화가 유의하게 억제되었다. 이러한 결과는 Fmo2 가 미토콘드리아 형태 조절 단백질에 의해 매개되는 새로운 비만 관련 유전자일 가능성이 있음을 시사한다.
- 34 -
참고 문헌
1.
Obesity: preventing and managing the global epidemic. Report of a WHO
consultation. World Health Organ Tech Rep Ser 894: i-xii, 1-253, 2000
2.
Alirol E, Martinou JC: Mitochondria and cancer: is there a morphological
connection? Oncogene 25: 4706-4716, 2006
3.
Ando Y, Sato F, Fukunaga H, Iwasaki Y, Chiba Y, Tebakari M, Daigo Y,
Kawashima J, Kamei J: Placental extract suppresses differentiation of
3T3-L1 preadipocytes to mature adipocytes via accelerated activation of
p38 MAPK during the early phase of adipogenesis. Nutr Metab (Lond) 16:
32, 2019
4.
Baloh RH: Mitochondrial dynamics and peripheral neuropathy.
Neuroscientist 14: 12-18, 2008
5.
Bennett CF, Kwon JJ, Chen C, Russell J, Acosta K, Burnaevskiy N, Crane
MM, Bitto A, Vander Wende H, Simko M, Pineda V, Rossner R, Wasko
BM, Choi H, Chen S, Park S, Jafari G, Sands B, Perez Olsen C,
Mendenhall AR, Morgan PG, Kaeberlein M: Transaldolase inhibition
impairs mitochondrial respiration and induces a starvation-like longevity
response in Caenorhabditis elegans. PLoS Genet 13: e1006695, 2017
- 35 -
6.
Bleich S, Cutler D, Murray C, Adams A: Why is the developed world
obese? Annu Rev Public Health 29: 273-295, 2008
7.
Cerveny KL, Tamura Y, Zhang Z, Jensen RE, Sesaki H: Regulation of
mitochondrial fusion and division. Trends Cell Biol 17: 563-569, 2007
8.
Chan DC: Mitochondria: dynamic organelles in disease, aging, and
development. Cell 125: 1241-1252, 2006a
9.
Chan DC: Mitochondrial fusion and fission in mammals. Annu Rev Cell
Dev Biol 22: 79-99, 2006b
10.
Chen H, Chomyn A, Chan DC: Disruption of fusion results in
mitochondrial heterogeneity and dysfunction. J Biol Chem 280:
26185-26192, 2005
11.
Chen H, Detmer SA, Ewald AJ, Griffin EE, Fraser SE, Chan DC:
Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion
and are essential for embryonic development. J Cell Biol 160: 189-200,
2003
12.
Chen H, McCaffery JM, Chan DC: Mitochondrial fusion protects against
neurodegeneration in the cerebellum. Cell 130: 548-562, 2007
13.
Cheung EC, McBride HM, Slack RS: Mitochondrial dynamics in the
regulation of neuronal cell death. Apoptosis 12: 979-992, 2007
- 36 -
14.
Cho E, Jung W, Joo HY, Park ER, Kim MY, Kim SB, Kim KS, Lim YB,
Lee KH, Shin HJ: Cluh plays a pivotal role during adipogenesis by
regulating the activity of mitochondria. Sci Rep 9: 6820, 2019
15.
Cho YS, Go MJ, Kim YJ, Heo JY, Oh JH, Ban HJ, Yoon D, Lee MH, Kim
DJ, Park M, Cha SH, Kim JW, Han BG, Min H, Ahn Y, Park MS, Han
HR, Jang HY, Cho EY, Lee JE, Cho NH, Shin C, Park T, Park JW, Lee
JK, Cardon L, Clarke G, McCarthy MI, Lee JY, Lee JK, Oh B, Kim HL:
A large-scale genome-wide association study of Asian populations
uncovers genetic factors influencing eight quantitative traits. Nat Genet
41: 527-534, 2009
16.
Chu DT, Tao Y, Tasken K: OPA1 in Lipid Metabolism: Function of
OPA1 in Lipolysis and Thermogenesis of Adipocytes. Horm Metab Res
49: 276-285, 2017
17.
De Pauw A, Tejerina S, Raes M, Keijer J, Arnould T: Mitochondrial
(dys)function in adipocyte (de)differentiation and systemic metabolic
alterations. Am J Pathol 175: 927-939, 2009
18.
Detmer SA, Chan DC: Functions and dysfunctions of mitochondrial
dynamics. Nat Rev Mol Cell Biol 8: 870-879, 2007
19.
Ghaben AL, Scherer PE: Adipogenesis and metabolic health. Nat Rev Mol
- 37 -
20.
Griparic L, van der Wel NN, Orozco IJ, Peters PJ, van der Bliek AM:
Loss of the intermembrane space protein Mgm1/OPA1 induces swelling
and localized constrictions along the lengths of mitochondria. J Biol Chem
279: 18792-18798, 2004
21.
Hofmann AD, Beyer M, Krause-Buchholz U, Wobus M, Bornhauser M,
Rodel G: OXPHOS supercomplexes as a hallmark of the mitochondrial
phenotype of adipogenic differentiated human MSCs. PLoS One 7:
e35160, 2012
22.
Hoppins S, Lackner L, Nunnari J: The machines that divide and fuse
mitochondria. Annu Rev Biochem 76: 751-780, 2007
23.
Kameoka S, Adachi Y, Okamoto K, Iijima M, Sesaki H: Phosphatidic
Acid and Cardiolipin Coordinate Mitochondrial Dynamics. Trends Cell
Biol 28: 67-76, 2018
24.
Kang SI, Ko HC, Shin HS, Kim HM, Hong YS, Lee NH, Kim SJ:
Fucoxanthin exerts differing effects on 3T3-L1 cells according to
differentiation stage and inhibits glucose uptake in mature adipocytes.
Biochem Biophys Res Commun 409: 769-774, 2011
25.
Kersten S: Mechanisms of nutritional and hormonal regulation of
lipogenesis. EMBO Rep 2: 282-286, 2001
- 38 -
26.
Lahera V, de Las Heras N, Lopez-Farre A, Manucha W, Ferder L: Role of
Mitochondrial Dysfunction in Hypertension and Obesity. Curr Hypertens
Rep 19: 11, 2017
27.
Lee S, Jeong SY, Lim WC, Kim S, Park YY, Sun X, Youle RJ, Cho H:
Mitochondrial fission and fusion mediators, hFis1 and OPA1, modulate
cellular senescence. J Biol Chem 282: 22977-22983, 2007
28.
Leiser SF, Miller H, Rossner R, Fletcher M, Leonard A, Primitivo M,
Rintala N, Ramos FJ, Miller DL, Kaeberlein M: Cell nonautonomous
activation of flavin-containing monooxygenase promotes longevity and
health span. Science 350: 1375-1378, 2015
29.
Li Q, Gao Z, Chen Y, Guan MX: The role of mitochondria in osteogenic,
adipogenic and chondrogenic differentiation of mesenchymal stem cells.
Protein Cell 8: 439-445, 2017
30.
Lopez-Lluch G: Mitochondrial activity and dynamics changes regarding
metabolism in ageing and obesity. Mech Ageing Dev 162: 108-121, 2017
31.
McBride HM, Neuspiel M, Wasiak S: Mitochondria: more than just a
powerhouse. Curr Biol 16: R551-560, 2006
32.
Morandi A, Zusi C, Corradi M, Olivieri F, Piona C, Fornari E, Maffeis C:
Minor diplotypes of FMO3 might protect children and adolescents from
obesity and insulin resistance. Int J Obes (Lond) 42: 1243-1248, 2018
- 39 -
33.
Perfettini JL, Roumier T, Kroemer G: Mitochondrial fusion and fission in
the control of apoptosis. Trends Cell Biol 15: 179-183, 2005
34.
Pidoux G, Witczak O, Jarnaess E, Myrvold L, Urlaub H, Stokka AJ,
Kuntziger T, Tasken K: Optic atrophy 1 is an A-kinase anchoring protein
on lipid droplets that mediates adrenergic control of lipolysis. EMBO J 30:
4371-4386, 2011
35.
Pruim RJ, Welch RP, Sanna S, Teslovich TM, Chines PS, Gliedt TP,
Boehnke M, Abecasis GR, Willer CJ: LocusZoom: regional visualization
of genome-wide association scan results. Bioinformatics 26: 2336-2337,
2010
36.
Rabbee N, Speed TP: A genotype calling algorithm for affymetrix SNP
arrays. Bioinformatics 22: 7-12, 2006
37.
Scheckhuber CQ, Erjavec N, Tinazli A, Hamann A, Nystrom T, Osiewacz
HD: Reducing mitochondrial fission results in increased life span and
fitness of two fungal ageing models. Nat Cell Biol 9: 99-105, 2007
38.
Schugar RC, Shih DM, Warrier M, Helsley RN, Burrows A, Ferguson D,
Brown AL, Gromovsky AD, Heine M, Chatterjee A, Li L, Li XS, Wang
Z, Willard B, Meng Y, Kim H, Che N, Pan C, Lee RG, Crooke RM,
Graham MJ, Morton RE, Langefeld CD, Das SK, Rudel LL, Zein N,
McCullough AJ, Dasarathy S, Tang WHW, Erokwu BO, Flask CA,
- 40 -
Laakso M, Civelek M, Naga Prasad SV, Heeren J, Lusis AJ, Hazen SL,
Brown JM: The TMAO-Producing Enzyme Flavin-Containing
Monooxygenase 3 Regulates Obesity and the Beiging of White Adipose
Tissue. Cell Rep 19: 2451-2461, 2017
39.
Scott F, Gonzalez Malagon SG, O'Brien BA, Fennema D, Veeravalli S,
Coveney CR, Phillips IR, Shephard EA: Identification of
Flavin-Containing Monooxygenase 5 (FMO5) as a Regulator of Glucose
Homeostasis and a Potential Sensor of Gut Bacteria. Drug Metab Dispos
45: 982-989, 2017
40.
Smith A, Yu X, Yin L: Diazinon exposure activated transcriptional factors
CCAAT-enhancer-binding proteins alpha (C/EBPalpha) and peroxisome
proliferator-activated receptor gamma (PPARgamma) and induced
adipogenesis in 3T3-L1 preadipocytes. Pestic Biochem Physiol 150:
48-58, 2018
41.
Suh JK, Poulsen LL, Ziegler DM, Robertus JD: Yeast flavin-containing
monooxygenase generates oxidizing equivalents that control protein
folding in the endoplasmic reticulum. Proc Natl Acad Sci U S A 96:
2687-2691, 1999
- 41 -
42.
Suh JK, Robertus JD: Yeast flavin-containing monooxygenase is induced
by the unfolded protein response. Proc Natl Acad Sci U S A 97: 121-126,
2000
43.
Swindell WR: Genes and gene expression modules associated with caloric
restriction and aging in the laboratory mouse. BMC Genomics 10: 585,
2009
44.
Szabadkai G, Simoni AM, Bianchi K, De Stefani D, Leo S, Wieckowski
MR, Rizzuto R: Mitochondrial dynamics and Ca2+ signaling. Biochim
Biophys Acta 1763: 442-449, 2006
45.
Takahashi Y, Shinoda A, Kamada H, Shimizu M, Inoue J, Sato R:
Perilipin2 plays a positive role in adipocytes during lipolysis by escaping
proteasomal degradation. Sci Rep 6: 20975, 2016
46.
Tatsuta T, Langer T: Quality control of mitochondria: protection against
neurodegeneration and ageing. EMBO J 27: 306-314, 2008
47.
Tontonoz P, Hu E, Spiegelman BM: Stimulation of adipogenesis in
fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell
79: 1147-1156, 1994
48.
Tontonoz P, Spiegelman BM: Fat and beyond: the diverse biology of
PPARgamma. Annu Rev Biochem 77: 289-312, 2008
- 42 -
49.
Tormos KV, Anso E, Hamanaka RB, Eisenbart J, Joseph J, Kalyanaraman
B, Chandel NS: Mitochondrial complex III ROS regulate adipocyte
differentiation. Cell Metab 14: 537-544, 2011
50.
Twig G, Elorza A, Molina AJ, Mohamed H, Wikstrom JD, Walzer G,
Stiles L, Haigh SE, Katz S, Las G, Alroy J, Wu M, Py BF, Yuan J,
Deeney JT, Corkey BE, Shirihai OS: Fission and selective fusion govern
mitochondrial segregation and elimination by autophagy. EMBO J 27:
433-446, 2008
51.
Vernochet C, Damilano F, Mourier A, Bezy O, Mori MA, Smyth G,
Rosenzweig A, Larsson NG, Kahn CR: Adipose tissue mitochondrial
dysfunction triggers a lipodystrophic syndrome with insulin resistance,
hepatosteatosis, and cardiovascular complications. FASEB J 28:
4408-4419, 2014
52.
Yazdi FT, Clee SM, Meyre D: Obesity genetics in mouse and human:
back and forth, and back again. PeerJ 3: e856, 2015
53.
Youle RJ, Karbowski M: Mitochondrial fission in apoptosis. Nat Rev Mol
Cell Biol 6: 657-663, 2005
54.
Yu T, Robotham JL, Yoon Y: Increased production of reactive oxygen
species in hyperglycemic conditions requires dynamic change of
- 43 -
mitochondrial morphology. Proc Natl Acad Sci U S A 103: 2653-2658,
2006
55.
Zhang Y, Marsboom G, Toth PT, Rehman J: Mitochondrial respiration
regulates adipogenic differentiation of human mesenchymal stem cells.
- 44 -
- Abstract -
Identification of novel obesity-related genes associated with
mitochondrial morphology-regulating protein OPA1
Seulbi Park
Department of Medical Sciences
The Graduate School, Ajou University
(Supervised by Professor Seon-Yong Jeong)
Mitochondria are the primary energy-generating organelles in most eukaryotic
cells. Mitochondria continuously change their morphology and this process is
involved in a variety of mitochondrial functions, including apoptosis, intermediary
metabolism, calcium signaling, and obesity-related phenotypes. Mitochondrial
morphology is precisely controlled by fusion and fission machinery involving
many mitochondria-shaping proteins, such as Drp1, Fis1, Mfn1, Mfn2, and Opa1.
In this study, I aimed to identify the key mitochondrial-shaping protein(s)
involved in obesity-related phenotypes. I selected 647 single nucleotide
polymorphisms (SNPs) on 49 genes encoding mitochondrial-shaping proteins.
Using the clinical, SNP genotyped, and epidemiological data from 3,747 subjects
- 45 -
of the Korean Association REsourece (KARE) study cohort, I performed an
association study of 647 SNPs with obesity-related traits. I found one SNP
(rs7646539) located on the OPA1 gene which satisfied the Bonferroni-correction
threshold p-value and was closely associated with male abdominal fat rate (p =
0.00034). Several SNPs located on the OPA1 gene and linked with rs7646539
showed a significant association with male abdominal fat rate. An Opa1
knockdown in 3T3-L1 preadipocyte cells showed a significant decrease in the Oil
Red O staining levels and expression of adipogenesis markers—Adipoq, Cebpa,
Fabp4, Slc2a4, and Pparg— as compared to the Opa1 expressing control cells.
Next, I conducted a whole-genome RNA-sequencing analysis in Opa1
knockdown 3T3-L1 cells and screened the differentially expressed genes. I found
the Fmo2 gene (encoding flavin-containing monooxygenase 2), which has not
previously been reported to be related to any obesity-related traits and diseases.
Knockdown of Fmo2 gene revealed a significant decrease in the Oil Red O staining
levels and in the expression of adipogenesis markers, Adipoq, Cebpa, Fabp4,
Slc2a4, and Pparg genes, as compared to Fmo2 expressing control cells.
These results suggest that the Fmo2 gene may be a new obesity-related gene
which is regulated by mitochondrial fusion Opa1 gene.
_______________________________________________________________________