저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학석사
학위논문
P301L 형질전환 생쥐의 해마체에서
Reelin을 분비하는 nNOS 양성
GABA성 신경세포의 사멸
아 주 대 학 교 대 학 원
의생명과학과 / 신경과학전공
김 남 중
P301L 형질전환 생쥐의 해마체에서
Reelin을 분비하는 nNOS 양성
GABA성 신경세포의 사멸
지도교수
이 영 돈
이
논문을 이학석사 학위 논문으로 제출함
2015년 6월
아
주 대 학 교 대 학 원
의생명과학과
/ 신경과학전공
김
남 중
김남중의
이학석사 학위 논문을 인준함
심사위원장
이 영 돈
(서명)
심 사 위 원
서 해 영
(서명)
심 사 위 원
백 은 주
(서명)
아
주 대 학 교 대 학 원
2015 년 6 월 22 일
I -국 문 요 약 -
P301L 형 질 전 환 생 쥐 의 해 마 체 에 서 Reelin을
분 비 하 는
nNOS 양 성 GABA성 신 경 세 포 의 사 멸
Reelin은 세포외 기질 당단백질로서 초기 뇌의 발달 과정에서 신경세포의 이동 에 있어서 중요한 역할을 한다. 또한, 성인 뇌의 겉질과 해마에 있는 GABA성 신 경세포에서 발현하여 시냅스 가소성과 기억을 조절한다. Reelin을 발현하는 GABA성 신경세포의 subtype들 중에서 해마에서는 neuronal nitric oxide synthases (nNOS)를 발현하는 GABA성 신경세포가 가장 높은 비율로 존재하고 있다. 이러 한 GABA성 신경세포의 생존에 있어서 Tau의 인산화가 중요한 역할을 담당하고 있으며, 그렇기 때문에 본 연구에서는, FTDP-17 (Frontotemporal dementia and parkinsonism linked to chromosome 17) 질환 모델로서 Tau 단백질의 과인산화를 유 도한 P301L 생쥐를 이용하여 해마 (hippocampus)에서 reelin과 nNOS를 공동으로 발현하는 GABA성 신경세포의 세포사멸에 대해서 알아보고자 했다.이중면역조직화학법을 이용하여 P301L 생쥐의 해마에서 nNOS를 발현하는 신경 세포가 GABA성 신경세포의 한 종류라는 것을 GABA의 마커인 GAD67을 통하여 확인하였으며, nNOS 양성 GABA성 신경세포에서 reelin을 발현하고 있는 신경세 포들을 확인하였다. 이러한 reelin을 발현하는 nNOS 양성 GABA성 신경세포들은 P301L 대조군 생쥐에 비해서 형질전환 P301L 생쥐의 문 (hilus)에서 줄어들었고, 줄어든 신경세포들에서 Tau의 과인산화와 세포사멸이 일어나는 것이 관찰되었다. 또한, Reelin을 발현하는 nNOS 양성 GABA성 신경세포의 사멸로 인하여 reelin의 발현이 감소하였을 때, 이 reelin의 감소는 해마의 피라미드 세포와 reelin을 분비 하는 세포 자체에 영향을 준다.
II
차
례
국 문 요 약... Ⅰ 차 례 ... Ⅱ 그 림 차 례 ... Ⅲ 표 차 례 ... Ⅳ Ⅰ. 서 론 ... 1 Ⅱ. 재 료 및 방 법 ... 4 1. 실험동물 ... 4 2. 조직 표본 제작 ... 4 3. 이중면역조직화학법...
4 Ⅲ. 결 과 ... 7 1. P301L 생쥐의 해마에서 GAD67과 nNOS의 공동발현 ... 7 2. P301L 생쥐의 해마에서 nNOS와 Reelin의 공동발현 ... 10 3. P301L 생쥐의 치아이랑에서 Reelin을 발현하는 nNOS 양성세포 수의 감소 ... 12 4. P301L 생쥐에서 Reelin 수용체의 발현 ... 14 5. P301L 생쥐의 해마에서 Tau의 과인산화에 의한 Reln을 발현하는 nNOS 양성세포의 사멸 ... 16 Ⅳ. 고 찰 ... 19 참 고 문 헌... 22 영 문 요 약... 26III
LIST OF FIGURES
Fig. 1. The coexpr ess ion of GAD67 and nNOS in t he hippoca mpus in P301L mice ... 9 Fig. 2. The coexpr ess ion of r eelin and nNOS in s evera l hippoca mpa l regions in P301L mice ... 11 Fig. 3. The decr eas e of the number of nNOS-Reln coexpr ess ed cells in
P301L mice ... 13 Fig. 4. The expr ess ion of reelin r ecept or in P301L mice ... 15 Fig. 5. The cell deat h of nNOS pos it ive neurons by tau
IV
LIST OF TABLE
1
Ⅰ
. 서 론
1995년에 돌연변이 생쥐인 reeler에서 reelin 유전자가 망가져 있는 것이 발견되었다 (D'Arcangelo et al., 1995). Reelin은 세포외 기질 당단백질로서 수용체인 ApoER2와 VLDLR에 결합하여 dab1의 tyrosine 인산화를 유도하고 glycogen Synthase Kinase 3 beta (GSK3β)와 CREB (cAMP response element-binding protein)을 활성화하여 발달뇌에서 신경세포의 이동과 위치선정 과정을 조절한다. 또한, 장기강화작용 (LTP)을 유도하여 시냅스 가소성 (synaptic plasticity)을 조절하거나 수상돌기 (dendrite)와 수상돌기극 (dendritic spine) 발달을 자극한다 (Weeber et al., 2002; Niu et al., 2004; D'Arcangelo, 2005; Niu et al., 2008). Reelin 유전자가 결여된 reeler는 뇌의 발달 과정에서 야생형 생쥐와는 반대로 대뇌피질의 신경세포층이 pial surface쪽에서부터 ventricular zone 방향으로 층을 형성하게 된다. 또한, 소뇌와 해마의 형태도 구조적으로 망가지거나 크기가 줄어들게 된다 (D'Arcangelo and Curran, 1998). reelin은 태아의 발달하는 뇌의 신피질에서 Cajal-Retzius 신경세포에 의해서 만들어지고, 출생 이후에는 주로 신피질과 해마의 GABA성 신경세포 subtype에 의해서 만들어진다 (Alcantara et al., 1998; Pesold et al., 1998; Tissir and Goffinet, 2003; D'Arcangelo, 2006).
뇌의 신경회로는 흥분성인 glutamatergic 신경세포와 억제성인 GABA성 신경세포
두 가지로 이루어져 있다. 모든 피질 회로에서 80~90%는 glutamatergic 신경세포 이며, 10~20%가 GABA성 신경세포이다 (Halasy and Somogyi, 1993; Aika et al., 1994; Freund and Buzsaki, 1996). 이중에서 GABA성 신경세포는 작은 비율에도 불구하고 전체 뇌에서 으뜸 신경세포들의 활성을 조절하는 중요한 기능을 하며 GABA는 주로 interneuron에 많이 분포한다 (Hu et al., 2014). 이러한 GABA는 glutamic acid decarboxylase (GAD) 효소에 의해서 glutamate로부터 만들어지며, 이 효소가 부족 할 경우에 GABA 기능의 고장으로 경련이 유발되고 간질이 발생할 수 있다 (Benke, 2013). GABA성 신경세포는 화학 표지자들의 종류에 따라서 8가지 종류의 subtype을 가진다. 칼슘 결합 단백질인 parvalbumin (PV), calretinin (CR), calbindin
2
(CB)을 발현하는 GABA성 신경세포와 신경펩티드인 somatostatin (SOM), neuropeptide Y (NPY), cholecystokinin (CCK), vasoactive intestinal protein (VIP)를 발현 하는 GABA성 신경세포 그리고 세포내 화학적 신경전달 물질인 nNOS를 발현하 는 GABA성 신경세포가 있다. 이러한 GABA성 신경세포의 생존과 기능에 있어 서 미세소관 안정화 단백질인 Tau가 중요한 역할을 한다 (Levenga et al., 2013).
미 세 소 관 결 합 단 백 질 (microtubule-ass ocia ted pr ot ein, M AP)의 하 나 인 Tau는 주 로 신 경 세 포 의 축 색 돌 기 에 서 발 현 이 되 며 미 세 소 관 에 결 합 하 여 미 세 소 관 을 안 정 화 시 키 는 역 할 을 한 다 (Dr echs el et al., 1992). 인 간 에 서 발 현 되 는 Tau 단 백 질 은 유 전 자 의 exon 중 에 서 2, 3, 10번 exon의 alt ernat ive splicing으 로 6개 의 subtype을 가 진 다 (Goedert et al., 1989). 그 중 에 서 도 exon 10번 의 splicing 결 과 에 따 라 서 3개 또 는 4 개 의 microtubule-binding r epeats을 가 지 는 subtype이 생 기 게 된 다 . 이 러 한 Tau 단 백 질 의 과 인 산 화 는 신 경 퇴 행 성 질 환 의 주 요 한 원 인 으 로 예 상 되 어 지 고 있 으 며 , Tau 단 백 질 의 미 세 소 관 에 대 한 결 합 력 을 약 화 시 켜 미 세 소 관 의 구 조 를 불 안 정 하 게 만 들 고 , 결 과 적 으 로 신 경 세 포 의 축 삭 이 동 (axonal transport)을 비 정 상 적 으 로 일 어 나 게 만 들 어 신 경 세 포 의 퇴 행 을 야 기 하 게 된 다 (Drechs el et al., 1992). 연 구 에 사 용 하 는 Tau 모델 생쥐는 Tau의 유전자를 과발현시키거나 제거한 모델과 Tau의 인 산화 kinase의 발현을 활성화하거나 억제하는 방식으로 Tau 단백질의 인산화를 비정상적으로 일어나도록 유도한 생쥐 모델이 있다 (Hernandez et al., 2013). 이렇 게 여러가지 종류의 Tau 질환 모델 중에서 인간의 일반형 Tau 유전자를 과발현 시킨 생쥐 모델은 늙은 생쥐에서 인산화된 형태의 Tau 단백질이 발견되었을 뿐, 기억력에는 문제가 나타나지 않았다 (Gotz et al., 1995). 이러한 문제를 보완하기 위해서 Frontotemporal dementia 환자의 뇌에서 나타나는 돌연변이 Tau 유전자 (P301L, R406W)를 과발현시킨 쥐들이 사용되었는데, 이 돌연변이 Tau 유전자를 발현하는 쥐들에서는 Tau의 응집현상과 과인산화, 신경섬유다발 형성, 기억력 감 소 등 Tau 질환에서 나타나는 일반적인 특징들이 나타난다 (Lewis et al., 2000;
3
Spittaels et al., 2000; Lucas et al., 2001). 그 중에서도 P301L 생쥐는 가장 일반적으로 사용되어지는 FTDP-17 질환 동물 모델로서 Tau의 301번 codon에서 Proline이 Leucine으로 돌연변이가 일어나 Tau의 과인산화가 일어나도록 유도된 생쥐 모델 이다. 따라서 본 연구에서는 이 형질전환 P301L 생쥐와 정상 생쥐의 해마를 비 교하여 Tau의 과인산화에 의한 reelin을 발현하는 GABA성 신경세포의 사멸에 대 하여 알아보고자 했다.
4
Ⅱ . 재 료 및 방 법
1. 실험동물 Tau의 과인산화를 유도한 FTDP-17 동물모델인 P301L과 대조군으로 B6D2F1 생 쥐를 사용하였다. 사육 중인 생쥐는 일반시판용 고형사료와 물을 자유롭게 섭취 토록 하였으며 동물실의 실내온도는 23°C, 습도는 60% 그리고 12시간씩의 낮과 밤주기가 항상 유지되도록 하였다. 2. 조직 표본 제작15개월된 P301L 생쥐 4마리를 avertin (2,2,2 tribromoethanol, Sigma-Aldrich, T48402) 0.4 mg/g으로 마취시킨 후, 0.9% 식염수 500ml로 심장을 관류시켜서 생쥐의 혈액을 제거하고, 4% 고정액 (paraformaldehyde, 4°C, PH 6.8~7.2)으로 하루동안 고정시켰다. 다음날 조직을 0.1M PBS (PH 7.2)로 3번 세척하고, Leica VT1200s Vibratome으로 조직을 잘랐다. 자른 부위는 해마의 앞쪽부위가 나타나기 시작하는 Bregma -0.8 mm 에서부터 해마 뒤쪽부위가 끝나기 전 -3.0 mm 사이를 연속해서 관상면 (coronal plane)으로 40 μm씩 잘랐다. 자른 절편은 cryoprotectant solution (0.1M PBS, 30% Glycerin, 30% Ethylen glycol)에 담가 -20°C에 하루동안 보관하고, 다음날 -80°C로 이동하여 보관했다.
3. 이중 면역 조직 화학법
0.1M PBS (PH 7.2)으로 15분씩 3번 세척하고, 비특이적 반응을 제거하기 위해서 차단 용액 (10% normal goat serum, 0.1% BSA, 0.1% triton-X 100)으로 상온에서 1시간 동안 반응시켰다. 이것을 Reelin, GAD67, nNOS, Dab1, ApoER2, VLDLR, AT8, AIF, FAF1에 대한 일차 항체로 16시간 동안 4°C에서 반응시켰다. 다음날 상온에서 1
5
시간 동안 더 반응시키고 PBS로 15분씩 3번 세척했다. 이차항체 anti-mouse IgG coupled to Alexa488, anti-rabbit IgG coupled to Alexa568을 상온에서 1시간 동안 반응 시켰다. 이차항체의 반응이 끝나고, PBS로 15분씩 3번 세척후에 Fluoromount-G (Southern Biotech, inc.)로 봉입한 후 Confocal microscopy로 관찰했다.
6
Table 1. Information and conditions of antibodies
Antibodies Sources Dilutions Companies Reelin Mouse 1:200 Millipore GAD67 Mouse 1:200 Millipore
nNOS Dab1 Rabbit Rabbit 1:200 1:200 Millipore Santa Cruz ApoER2 Rabbit 1:200 Santa Cruz VLDLR Rabbit 1:200 Santa Cruz AIF Rabbit 1:200 Cell Signalling FAF1 Rabbit 1:200 Proteintech AT8 Mouse 1:100 Pierce
7
Ⅲ
. 결 과
1
.
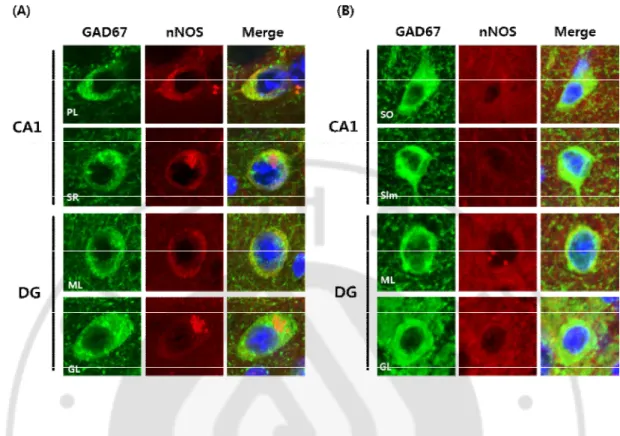
P301L 생쥐의 해 마 에 서 GAD67과 nNO S의 공 동 발 현해마는 시상 (thalamus)을 둘러싸고 있는 원통형 구조로 이루어져 있으며, 크게 CA1~3과 치아이랑으로 구성되어 있다. CA1과 CA2는 다시 상행층 (stratum oriens) 과 피라미드층 (pyramidal layer), 방사층 (stratum radiatum)으로 나뉘어지고, CA3은 이 3개의 층에 투명층 (stratum lucidum)이 더 존재한다. 상행층에는 바구니세포 (basket cell)와 수평세겹세포 (horizontal trilaminar cell)의 세포체 그리고 피라미드세 포 (pyramidal cell)의 바닥가지돌기 (basal dendrite) 부분이 위치하고 있다. 피라미 드층에는 부챗살세겹세포 (radial trilaminar cell)와 축삭축삭세포 (axoaxonic cell)의 세포체가 위치하고 있으며, 방사층에는 사이막 (septal)과 맞교차섬유 (commissural fibers)가 존재한다. CA3에만 존재하는 투명층은 해마에서 가장 얇은 층으로 치아 이랑과 과립세포들로부터 이끼섬유 (mossy fiber)가 뻗어 있다. CA3와 서로 붙어 있는 치아이랑은 분자층 (molecular layer)과 과립층 (granular layer), 다형층 (polymorphic layer)으로 이루어져 있고, 내후각피질 (entorhinal cortex)로부터 뻗어나 오는 관통경로 섬유 (perforant pathway fiber)들이 치아이랑의 분자층을 통해서 CA3와 연결되어 있으며, 과립층 안쪽에서 생성되는 과립세포들은 과립층에서 성 숙한 세포가 되어 다형층을 지나 CA3으로 이동한다. 이 해마의 구조중에서 상대 적으로 적은 면적을 차지하고 있는 CA2를 제외하고, P301L 생쥐의 CA1과 CA3, DG에서의 nNOS를 발현하는 GABA성 신경세포에 대해서 알아봤다. 신경세포에 서 Neuronal nitric oxide synthase (nNOS)는 해마와 치아이랑의 GABA성 세포에서 만들어지는 것으로 알려져있다. CA1의 피라미드세포층 (pyramidal layer)과 방사층 (stratum radiatum)에서 GABA성 신경세포들 중 일부가 nNOS를 공동으로 발현을 하고 있었으며, 치아이랑에서는 분자층과 과립층의 GABA성 신경세포들에서 nNOS를 공동으로 발현하거나 GAD만을 발현하는 신경세포들이 확인되었다. CA1 과 치아이랑의 다른 층들 그리고 CA3에서는 nNOS와 GAD를 공동으로 발현하는
8
세포들이 관찰되지 않았다 (Fig. 1A). 이러한 결과는 GABA성 신경세포가 모두 다 nNOS를 발현하고 있지는 않으며, GABA성 신경세포들 중에서 일부가 nNOS 양성 GABA성 신경세포라는 것을 의미한다.
9
Fig. 1. The coexpression of GAD67 and nNOS in the hippocampus in P301L mice. The
GABAergic interneurons expressing glutamic acid decarboxylase 67 (GAD67) exhibit the colocalization of neuronal nitric oxide synthase (nNOS) in hippocampus. PL, pyramidal layer; SR, stratum radiatum; ML, molecular layer; GL, granular layer.
10
2. P301L 생쥐의 해마에서 nNOS와 Reln의 공동발현
GABA성 신경세포의 subtype들 중에서 Reln을 분비하는 종류로는 PV, SST, VIP, NPY, nNOS, CR 양성 세포들이 있고, 이중에서 nNOS 양성 GABA성 신경세포가 가장 높은 비율로 reelin을 발현하는 것으로 보고되어 있다 (Pohlkamp et al., 2014). 이러한 reelin을 발현하는 nNOS 양성 GABA성 신경세포가 P301L 생쥐의 대조군 에 비해서 형질전환 P301L 생쥐의 해마에서 수적으로 어떠한 변화를 보이는지는 아직 알려져 있지 않다. 따라서 우리는 P301L 생쥐의 대조군과 형질전환 생쥐의 해마를 비교하여 reelin을 발현하는 nNOS 양성 GABA성 신경세포 수의 변화를 알아보려고 했다.
Reeler에서 nNOS와 reelin이 해마와 치아이랑에서 서로 공동발현을 하고 있는 것으로 보고된 바 있다 (Romay-Tallon et al., 2010). nNOS와 reelin을 공동으로 발현
하는 신경세포는 NMDA 수용체 활성조절과 장기강화작용의 증가, 수상돌기극 형
성의 증가에 있어서 nNOS와 reelin간에 기능적인 면에서 서로 상호 보완적으로 작용한다 (Romay-Tallon et al., 2010). 본 연구에서는 FTDP-17 질환 모델인 P301L 생쥐의 뇌에서 nNOS와 reelin과의 공동발현을 살펴보았다.
해마의 CA1 부위와 CA3 부위의 피라미드세포층에서 nNOS를 발현하는 신경세 포들 중의 일부가 reelin과 서로 공동발현을 하고 있었고, 치아이랑에서는 문
(hilus)에서 nNOS를 발현하는 신경세포들 중에서 일부가 reelin과 공동발현을 하고 있었다 (Fig. 2). 이것은 nNOS를 발현하고 있는 GABA성 신경세포중 일부는 reelin 을 공동으로 발현한다는 것을 의미한다.
11
Fig. 2. The coexpression of reelin and nNOS in several hippocampal regions in P301L mice. Reelin was expressed in nNOS-positive GABAergic interneurons located throughout
12
3. P301L 생쥐의 치아이랑에서 Reln을 발현하는 nNOS 양성 신경세포 수의 감소
본 연구에서는 FTDP-17 모델 생쥐인 P301L을 이용하여 reelin과 nNOS를 공동으 로 발현하는 GABA성 신경세포의 수가 대조군에 비해서 P301L 생쥐에서 줄어드 는지를 알아보려고 했다. 대조군에 비해서 P301L 생쥐의 치아이랑 문에서 nNOS 를 발현하는 신경세포와 reelin을 발현하는 신경세포 그리고 reelin과 nNOS를 공 동으로 발현하는 신경세포들이 줄어들었다 (Fig. 3). 이러한 결과는 P301L 생쥐에 서 Tau 단백질의 과인산화에 의한 GABA성 신경세포의 사멸을 유추해 볼 수 있 게 하였다.
13
Fig. 3. Decreased number of nNOS-Reln expressing cells in P301L mice. The reelin
expression in nNOS positive neuronal population (arrows) is significantly decreased in P301L mice. Therefore, we suggested that reduced hippocampal reelin expression may be influence hippocampal functions through activation of reelin receptors, such as VLDLR and ApoER2. GL, granular layer; H, hilus.
14 4. P301L 생쥐에서 Reln 수용체의 발현
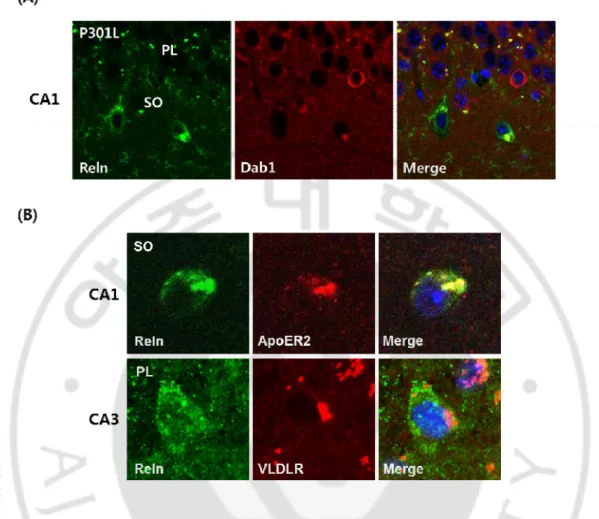
다음으로 P301L 생쥐의 해마에서 reelin의 신호를 받는 세포들은 어느곳에 위치 하고 있는지를 알아보기 위해서 reelin 신호의 하향 조절자인 Dab1과 reelin의 대 표적인 두가지 수용체인 ApoER2, VLDLR의 발현을 알아보고자 했다. CA1의 피 라미드층에서 Dab1을 발현하는 세포들과 reelin을 발현하는 세포들이 둘 다 관찰 되었다. 이것은 피라미드층에서 발현되어지는 reelin이 곁분비 방식으로 작용하거 나, 해마에서 발현되어지는 reelin이 피라미드 세포층으로 이동하여 작용한다는 것을 의미한다. 또한, 해마 부위중 CA1의 방사층에 있는 신경세포의 세포질에서 reelin과 ApoER2가 공동발현을 하고 있는 신경세포들이 발견이 되었고, CA3의 피 라미드층에 있는 신경세포의 세포질에서는 reelin과 VLDLR의 공동발현을 확인 할 수 있었다 (Fig. 4). 이것은 일부의 reelin을 발현하는 GABA성 신경세포는 reelin과 reelin 수용체들이 하나의 신경세포에서 공동으로 발현되어 자가분비 방 식으로 작용이 되어진다는 것을 의미한다.
15
Fig. 4. The expression of reelin receptor in P301L mice. (A) Dab1 was expressed within
the pyramidal layer in the CA regions. (B) Some reelin positive neurons coexpressed the reelin receptor ApoER2 and VLDLR. This result suggests that reelin secreting GABAergic interneurons may be one type of target cells affected by secreting reelin. PL, pyramidal layer; GL, granular layer; SO, stratum oriens.
16
5. P301L 생쥐의 해마에서 Tau의 과인산화에 의한 Reln을 발현하는 nNOS 양성
신경세포의 사멸
다음으로 P301L 생쥐에서 reelin과 nNOS를 공동으로 발현하는 GABA성 신경세
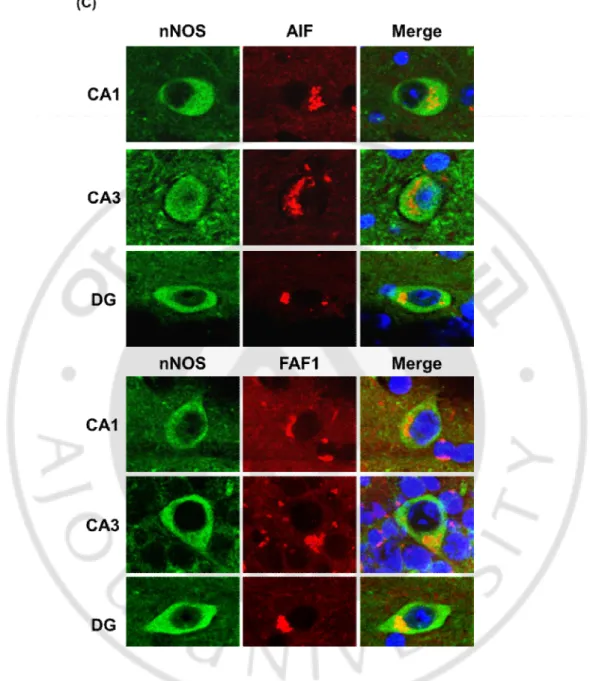
포가 줄어드는 원인을 알아보려고 했다. 해마와 치아이랑에서 nNOS를 발현하는 GABA성 신경세포들 중 일부는 Tau의 과인산화를 공동으로 발현하고 있었다 (Fig. 5A). 그리고 Tau의 과인산화가 일어난 GABA성 신경세포들 중 일부의 세포 에서 세포사멸 인자인 Apoptosis-inducing factor (AIF), Fas-associated factor-1 (FAF1)의 발현이 관찰되었다 (Fig. 5B). 또한, nNOS를 발현하는 GABA성 신경세포 중 일부 는 세포사멸 인자인 AIF, FAF1과 공동으로 발현을 하고 있었다 (Fig. 5C). 이러한 결과는 P301L 생쥐에서 nNOS를 발현하는 GABA성 신경세포가 세포 사멸이 일 어나며, 그 세포 사멸은 Tau의 과인산화에 의한 것이라는 것을 의미한다.
18
Fig. 5. The cell death of nNOS positive neurons by tau hyperphosphorylation in P301L mice. (A) nNOS positive neurons expressed hyperphosphorylated Tau (AT8). And the
expression level of AT8 is increased in P301L (data not shown). (B) AT8 positive neurons also colocalized with apototic molecules, such as AIF and FAF1. (C) Neurons expressing AIF and FAF1 were nNOS positive neurons. Therefore, this results suggest that nNOS positive GABAergic neuronal loss caused by hyperphosphorylated tau-induced apoptosis.
19
Ⅳ
. 고 찰
뇌에서 해마는 크게 치아이랑 (dentate zyrus)과 암몬각 (cornus ammonis)으로 나뉘어지고, CA는 다시 CA1-3 혹은 CA1-4로 나뉘어진다 (Tricoire and Vitalis, 2012). 이전에 연구에서 보고된 바에 따르면 nNOS는 GABA성 신경세포에서 만들어지며, GABA성 신경세포의 subtype은 해마와 신피질에서 서로 다른 비율로 존재한다. 해마에서는 nNOS를 발현하는 GABA성 신경세포가 가장 높은 비율로 존재하고, 신피질에서는 parvalbumin (PV)을 발현하는 GABA성 신경세포가 가장 높은 비율로 존재하고 있다 (Fuentealba et al., 2008). 따라서 본 연구에서는 먼저 알려진 것처럼 nNOS가 GABA성 신경세포에서 발현을 하는지를 알아보았다. 첫번째 결과에서 FTDP-17 질환 모델인 P301L의 해마에서 nNOS와 GABA성 신경세포의 표지자인 GAD가 서로 공동으로 발현을 하고 있는 것을 확인 할 수가 있었고, 이것은 nNOS를 발현하는 신경세포가 GABA성 신경세포의 한 종류라는 것을 의미한다.
Reelin은 NMDA 수용체의 활성을 조절하며 (Chen et al., 2005; Campo et al., 2009), 장기강화작용을 증가시킨다 (Weeber et al., 2002; Beffert et al., 2005; Beffert et al., 2006). 그리고 이러한 작용은 nitric oxide에 의해서 촉진되어진다 (Zhou and Zhu, 2009). 본 연구의 두번째에서 nNOS를 발현하는 신경세포가 reelin과 공동으로 발현하는지를 알아보려고 했다. CA1, CA3 부위의 피라미드층과 치아이랑의 문에서 nNOS와 reelin을 공동으로 발현하는 신경세포가 관찰되었고, 이것은 nNOS를 발현하는 GABA성 신경세포중 일부는 reelin을 공동으로 발현하고 있다는 것을 의미한다.
이전의 연구에서 reeler의 해마와 치아이랑에서 reelin과 nNOS는 서로 공동으로 발현이 되어지며 (Romay-Tallon et al., 2010), 알츠하이머병 모델 생쥐의 해마와 내후각 피질에서 reelin을 발현하는 피라미드 신경세포의 수가 줄어들고, 알츠하이머병 생쥐와 이형접합 reeler를 교배하여 태어난 생쥐에서 reelin의 부족에 의한 β-amyloide plaque과 Tau의 병리학적인 증상이 더 가속화 되어진다는
20
것이 보고되었다 (Herring et al., 2012). 또한, reelin과 nNOS의 공동발현은 reeler의 대조군에 비해서 이형접합 reeler의 치아이랑에서 감소한다. 이러한 이형접합 reeler에서의 reelin 양성 nNOS 신경세포 수의 감소가 결국, 신생 과립세포의 세포이동과 세포성숙을 붕괴시키는 것으로 예상하고 있다 (Lussier et al., 2009). 본 연구에서는 P301L 생쥐의 대조군에 비해서 유전자변형 생쥐에서 reelin을 발현하는 nNOS 신경세포의 수가 어떻게 달라지는지를 알아보려고 했다. 그 결과, reelin과 nNOS를 공동으로 발현하는 신경세포가 유전자변형 P301L 생쥐의 문에서 줄어드는 것이 관찰됐다. 이러한 결과를 통해서 이형접합 reeler의 치아 이랑에서처럼 유전자변형 P301L 생쥐의 치아이랑에서도 reelin과 nNOS를 공동으로 발현하는 신경세포의 감소에 의한 신생 과립세포의 세포이동과 세포 성숙과정이 붕괴되어질 가능성에 대해서 생각을 하게 되었다. 또한, 이러한 세포이동과 세포 성숙과정의 붕괴에 의해서 P301L 생쥐 치아이랑의 시냅스 가소성이 망가지고, reelin을 발현하는 신경세포 수의 감소에 의한 신경재생능력의 기능고장에 대해서 추측해 볼 수가 있었다.
Reelin의 수용체는 VLDLR과 ApoER2 두 가지의 Apolipoprotein E (ApoE) 수용체로 이루어져 있으며, 뇌발달 동안에 reelin의 신호에 의해서 신경세포의 이동이 조절된다 (Weeber et al., 2002). 이 두 가지 reelin 수용체는 reelin외에도 ApoE를 ligand로 가지고 있기 때문에 기억과 신경퇴행성 질환에 간접적으로 관여를 하게 된다 (Weeber et al., 2002). Reelin 수용체와 관련하여 본 연구에서는 reelin의 신호를 받는 세포들이 어느 곳에 위치하고 있는지를 알아보려고 했다. 그 결과, 해마의 피라미드층에서 reelin 신호의 하향조절자인 Dab1의 발현이
관찰되었다. 이것은 reelin을 받는 주요 목표 세포들이 피라미드층의
으뜸세포들이라는 것을 의미한다. 또한, CA1의 상행층 (stratum oriens)과 CA3의 피라미드층에 있는 reelin을 발현하는 세포에서 reelin 수용체들이 공동으로 발현되어지고 있는 것이 관찰되었다. 이것은 reelin을 발현하고 있는 세포가 그 스스로 reelin의 목표 세포가 되기도 한다는 것을 나타낸다.
21
형성하고 결국, 그 신경섬유소에 의해서 신경세포의 기능고장과 사멸이 일어난다 (Johnson and Stoothoff, 2004). 실제로 알츠하이머병 생쥐 모델중에서 삼중 유전자변형 생쥐 모델인 TauPS2APP의 해마에서 GABA성 신경세포의 세포소실이 일어난다고 보고되었다 (Loreth et al., 2012). 그렇기 때문에 FTDP-17 생쥐 모델인 P301L에서도 Tau의 과인산화에 의한 신경세포의 사멸이 일어날 것으로 예상을 할 수 있었다. 실제로, nNOS를 발현하는 신경세포에서 Tau의 과인산화 항체인 AT8과 세포괴사 인자인 AIF, FAF1이 서로 공동으로 발현을 하고 있는 것을 확인했으며, 이러한 결과는 P301L 생쥐의 해마에서 nNOS를 발현하고 있는 신경세포가 Tau의 과인산화에 의해서 세포사멸이 일어난다는 것을 의미한다.
여러가지 subtype의 GABA성 신경세포 중에서 nNOS를 발현하는 GABA성 신경세포만을 선택적으로 사멸시켰을 때, 생쥐 뇌의 전체 겉질망에서 약간의 탈 억제가 일어나는 것으로 보고되었다 (Shlosberg et al., 2012). 따라서, 본 연구와 관련해서 우리는 다음과 같은 것들에 대해서 생각을 하게 되었다. 첫째로, 유전자변형 P301L 생쥐에서 reelin과 nNOS를 공동으로 발현하는 신경세포가 사멸하였을 때, 신경세포의 사멸에 의한 reelin과 nNOS의 감소가 결과적으로 해마의 기능에 있어서 어떠한 영향을 미치게 될 것인지, 그리고 그 다음으로는 Tau의 과인산화에 의해서 GABA성 신경세포가 사멸할 때, nNOS를 발현하는 GABA성 신경세포와 다른 subtype들을 발현하는 GABA성 신경세포들과의 관련성은 없는지, 그리고 마지막으로 해마의 부위와 subtype에 따라서 reelin을 발현하는 GABA성 신경세포들이 사멸하는 경향은 어떻게 다르게 나타나게 될 것인지에 대해서 생각을 하게 되었다. 이러한 의문점들에 대한 결과를 알아내기 위해서 앞으로 behavior test를 비롯하여, 해마의 보다 넓은 범위를 통한 통계학적인 실험이 이루어져야 할 것으로 보인다.
22
참고문헌
1. Aika Y, Ren JQ, Kosaka K, Kosaka T: Quantitative analysis of
GABA-like-immunoreactive and parvalbumin-containing neurons in the CA1 region of the rat hippocampus using a stereological method, the disector. Exp Brain Res 99: 267-276, 1994
2. Alcantara S, Ruiz M, D'Arcangelo G, Ezan F, de Lecea L, Curran T, Sotelo C, Soriano E: Regional and cellular patterns of reelin mRNA expression in the forebrain of the developing and adult mouse. J Neurosci 18: 7779-7799, 1998 3. Beffert U, Durudas A, Weeber EJ, Stolt PC, Giehl KM, Sweatt JD, Hammer RE, Herz
J: Functional dissection of Reelin signaling by site-directed disruption of Disabled-1 adaptor binding to apolipoprotein E receptor 2: distinct roles in development and synaptic plasticity. J Neurosci 26: 2041-2052, 2006
4. Beffert U, Weeber EJ, Durudas A, Qiu S, Masiulis I, Sweatt JD, Li WP, Adelmann G, Frotscher M, Hammer RE, Herz J: Modulation of synaptic plasticity and memory by Reelin involves differential splicing of the lipoprotein receptor Apoer2. Neuron 47: 567-579, 2005
5. Benke D: GABAB receptor trafficking and interacting proteins: targets for the development of highly specific therapeutic strategies to treat neurological disorders? Biochem Pharmacol 86: 1525-1530, 2013
6. Campo CG, Sinagra M, Verrier D, Manzoni OJ, Chavis P: Reelin secreted by
GABAergic neurons regulates glutamate receptor homeostasis. PLoS One 4: e5505, 2009
7. Chen Y, Beffert U, Ertunc M, Tang TS, Kavalali ET, Bezprozvanny I, Herz J: Reelin modulates NMDA receptor activity in cortical neurons. J Neurosci 25: 8209-8216, 2005
8. D'Arcangelo G: Apoer2: a reelin receptor to remember. Neuron 47: 471-473, 2005 9. D'Arcangelo G: Reelin mouse mutants as models of cortical development
disorders. Epilepsy Behav 8: 81-90, 2006
10. D'Arcangelo G, Curran T: Reeler: new tales on an old mutant mouse. Bioessays 20: 235-244, 1998
23
related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature 374: 719-723, 1995
12. Drechsel DN, Hyman AA, Cobb MH, Kirschner MW: Modulation of the dynamic instability of tubulin assembly by the microtubule-associated protein tau. Mol Biol Cell 3: 1141-1154, 1992
13. Freund TF, Buzsaki G: Interneurons of the hippocampus. Hippocampus 6: 347-470, 1996
14. Fuentealba P, Begum R, Capogna M, Jinno S, Marton LF, Csicsvari J, Thomson A, Somogyi P, Klausberger T: Ivy cells: a population of nitric-oxide-producing, slow-spiking GABAergic neurons and their involvement in hippocampal network activity. Neuron 57: 917-929, 2008
15. Goedert M, Spillantini MG, Jakes R, Rutherford D, Crowther RA: Multiple isoforms of human microtubule-associated protein tau: sequences and localization in neurofibrillary tangles of Alzheimer's disease. Neuron 3: 519-526, 1989 16. Gotz J, Probst A, Spillantini MG, Schafer T, Jakes R, Burki K, Goedert M:
Somatodendritic localization and hyperphosphorylation of tau protein in transgenic mice expressing the longest human brain tau isoform. Embo j 14: 1304-1313, 1995
17. Halasy K, Somogyi P: Distribution of GABAergic synapses and their targets in the dentate gyrus of rat: a quantitative immunoelectron microscopic analysis. J Hirnforsch 34: 299-308, 1993
18. Hernandez F, Lucas JJ, Avila J: GSK3 and tau: two convergence points in Alzheimer's disease. J Alzheimers Dis 33 Suppl 1: S141-144, 2013
19. Herring A, Donath A, Steiner KM, Widera MP, Hamzehian S, Kanakis D, Kolble K, ElAli A, Hermann DM, Paulus W, Keyvani K: Reelin depletion is an early
phenomenon of Alzheimer's pathology. J Alzheimers Dis 30: 963-979, 2012 20. Hu H, Gan J, Jonas P: Interneurons. Fast-spiking, parvalbumin(+) GABAergic
interneurons: from cellular design to microcircuit function. Science 345: 1255263, 2014
21. Johnson GV, Stoothoff WH: Tau phosphorylation in neuronal cell function and dysfunction. J Cell Sci 117: 5721-5729, 2004
22. Levenga J, Krishnamurthy P, Rajamohamedsait H, Wong H, Franke TF, Cain P, Sigurdsson EM, Hoeffer CA: Tau pathology induces loss of GABAergic
24
interneurons leading to altered synaptic plasticity and behavioral impairments. Acta Neuropathol Commun 1: 34, 2013
23. Lewis J, McGowan E, Rockwood J, Melrose H, Nacharaju P, Van Slegtenhorst M, Gwinn-Hardy K, Paul Murphy M, Baker M, Yu X, Duff K, Hardy J, Corral A, Lin WL, Yen SH, Dickson DW, Davies P, Hutton M: Neurofibrillary tangles, amyotrophy and progressive motor disturbance in mice expressing mutant (P301L) tau protein. Nat Genet 25: 402-405, 2000
24. Loreth D, Ozmen L, Revel FG, Knoflach F, Wetzel P, Frotscher M, Metzger F, Kretz O: Selective degeneration of septal and hippocampal GABAergic neurons in a mouse model of amyloidosis and tauopathy. Neurobiol Dis 47: 1-12, 2012 25. Lucas JJ, Hernandez F, Gomez-Ramos P, Moran MA, Hen R, Avila J: Decreased
nuclear beta-catenin, tau hyperphosphorylation and neurodegeneration in GSK-3beta conditional transgenic mice. Embo j 20: 27-39, 2001
26. Lussier AL, Caruncho HJ, Kalynchuk LE: Repeated exposure to corticosterone, but not restraint, decreases the number of reelin-positive cells in the adult rat hippocampus. Neurosci Lett 460: 170-174, 2009
27. Niu S, Renfro A, Quattrocchi CC, Sheldon M, D'Arcangelo G: Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway. Neuron 41: 71-84, 2004
28. Niu S, Yabut O, D'Arcangelo G: The Reelin signaling pathway promotes dendritic spine development in hippocampal neurons. J Neurosci 28: 10339-10348, 2008 29. Pesold C, Impagnatiello F, Pisu MG, Uzunov DP, Costa E, Guidotti A, Caruncho HJ:
Reelin is preferentially expressed in neurons synthesizing gamma-aminobutyric acid in cortex and hippocampus of adult rats. Proc Natl Acad Sci U S A 95: 3221-3226, 1998
30. Pohlkamp T, David C, Cauli B, Gallopin T, Bouche E, Karagiannis A, May P, Herz J, Frotscher M, Staiger JF, Bock HH: Characterization and distribution of Reelin-positive interneuron subtypes in the rat barrel cortex. Cereb Cortex 24: 3046-3058, 2014
31. Romay-Tallon R, Dopeso-Reyes IG, Lussier AL, Kalynchuk LE, Caruncho HJ: The coexpression of reelin and neuronal nitric oxide synthase in a subpopulation of dentate gyrus neurons is downregulated in heterozygous reeler mice. Neural Plast 2010: 130429, 2010
25
32. Shlosberg D, Buskila Y, Abu-Ghanem Y, Amitai Y: Spatiotemporal alterations of cortical network activity by selective loss of NOS-expressing interneurons. Front Neural Circuits 6: 3, 2012
33. Spittaels K, Van den Haute C, Van Dorpe J, Geerts H, Mercken M, Bruynseels K, Lasrado R, Vandezande K, Laenen I, Boon T, Van Lint J, Vandenheede J, Moechars D, Loos R, Van Leuven F: Glycogen synthase kinase-3beta phosphorylates protein tau and rescues the axonopathy in the central nervous system of human four-repeat tau transgenic mice. J Biol Chem 275: 41340-41349, 2000
34. Tissir F, Goffinet AM: Reelin and brain development. Nat Rev Neurosci 4: 496-505, 2003
35. Tricoire L, Vitalis T: Neuronal nitric oxide synthase expressing neurons: a journey from birth to neuronal circuits. Front Neural Circuits 6: 82, 2012
36. Weeber EJ, Beffert U, Jones C, Christian JM, Forster E, Sweatt JD, Herz J: Reelin and ApoE receptors cooperate to enhance hippocampal synaptic plasticity and learning. J Biol Chem 277: 39944-39952, 2002
37. Zhou L, Zhu DY: Neuronal nitric oxide synthase: structure, subcellular localization, regulation, and clinical implications. Nitric Oxide 20: 223-230, 2009
26
-ABSTRACT-Cell death of reelin secreting GABAergic interneurons
in the hippocampus of P301L transgenic mice
Reelin is a largely secreted extracellular glycoprotein that plays a critical role in layer formation and neuronal migration in early developmental stage. In the adult brain, it also present particularly in GABAergic interneurons in the hippocampus and cortex and regulate synaptic plasticity and memory. Neuronal nitric oxide synthase (nNOS) expressing cells in hippocampus is one of the major subtypes of GABAergic interneurons. In the survival of these GABAergic interneurons, the hyperphosphorylation of tau is important factor. P301L transgenic mice are characterized by high expression of human tau containing frontotemporal dementia and parkinsonism linked to chromosome 17 (FTDP-17) mutation and show motor and behavioral deficits together with age-dependent development of neurofibrillary tangles. So, we investigated the neuronal death of reelin expressing nNOS-positive GABAergic interneurons in the hippocampus of P301L transgenic mice.
As results, nNOS-positive cells coexpressed GABAergic interneurons and secreted reelin. And the number of these cells be decreased in the hippocampus of P301L mice. We confirmed the coexpression of AT8 and nNOS-positive GABAergic interneurons in the cells contained apoptotic molecules such as AIF, FAF1. Also, the decreation of reelin may affect pyramidal neurons and reelin secreting cells according to analyzing the localization of reelin receptors.
Based on these results, we conclude that reduction of nNOS-positive GABAergic interneurons in hippocampus of P301L mice may be due to tau hyperphosphorylation-induced neuronal cell death.