저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Master’s Thesis of Science
in Agricultural Biotechnology
Prevalence and characteristics of
extended-spectrum β-lactamase and
plasmid-mediated quinolone resistance genes in
Escherichia coli isolated from retail raw
chicken in Korea
한국 유통닭고기에서 분리된
extended-spectrum
β-lactamase와 plasmid-mediated quinolone resistance
genes를 보유한 대장균의 분포와 특성 분석
February 2019
The Graduate School
Seoul National University
Department of Agricultural Biotechnology
Prevalence and characteristics of
extended-spectrum β-lactamase and
plasmid-mediated quinolone resistance genes in
Escherichia coli isolated from retail raw
chicken in Korea
Advisor: Sangryeol Ryu
Submitting a Master
’s Thesis of Science
in Agricultural Biotechnology
February, 2019
The Graduate School
Seoul National University
Department of Agricultural Biotechnology
Hyeeun Park
Confirming the master
’s thesis written by
Hyeeun Park
February, 2019
ABSTRACT
Hyeeun Park
Department of Agriculture Biotechnology
The Graduate school
Seoul National University
Extended-spectrum β-lactamase (ESBL)-producing Escherichia coli is a serious public health concern. The plasmid-mediated quinolone resistance (PMQR) genes are increasingly prevalent in the isolates from food-producing animals and food products, some of which also produce extended-spectrum β- lactamases (ESBLs). In this study, I investigated the antibiotic resistance, phylogenetic group, and virulence gene prevalence of 67 ESBL-producing E. coli strains isolated from retail chicken in Korea. In addition, I investigated the distribution of plasmid-mediated quinolone resistance (PMQR) genes in the total of 67 strains of ESBL-producing Escherichia coli. All of the strains possessed the blaCTX-M genes, dominantly blaCTX-M-65
(52.2%, 35/67) and blaCTX-M-55 (25.4%, 17/67), and three strains harbored
coli strains also carried blaTEM. Surprisingly, 35 (52.2%, 35/67)
ESBL-positive isolates were co-harboring PMQR determinants. The most prevalent PMQR was qnrS (91.4%, 32/35), and 6 of them also carried aac(6′)-Ib-cr. The oqxAB and aac(6′)-Ib-cr genes were detected in 2 (5.7%) isolates, and 1 (2.9%) isolate, respectively. No isolate was positive for qnrA, qnrB, and
qepA. Besides, the most common ESBL-type among the PMQR-positive
isolates was CTX-M-65 (91.4%, 32/35), and some of them simultaneously belonged to CTX-M-55, OXA, and/or TEM type. The results of antibiotic susceptibility tests revealed that 98.5% of the ESBL-positive strains were resistant to ≥ 3 different antibiotic classes, and all of the 35 strains that co-harbored PMQR determinants were found to be multi-drug resistant (MDR). Among them, the strains harboring both qnrS and aac(6′)-Ib-cr showed a synergistic effect on the resistance to ciprofloxacin and kanamycin. The results of a phylogenetic analysis showed that the group A is predominant (56.7%), followed by B1 (19.4%), E (8.9%), B2 (6%), and D (6%). Furthermore, the virulence genes associated with extraintestinal pathogenic
E. coli (ExPEC) were frequently detected in the strains of the groups B1, B2,
D, and E. The results in this study demonstrated that retail chicken is highly contaminated with ESBL-producing, multi-drug resistant E. coli and may
Keyword : Antibiotic resistance, Extended-spectrum spectrum β-lactamases
(ESBLs), Plasmid-mediated quinolone resistance (PMQR), Escherichia coli, Chicken
CONTENTS
ABSTRACT……….ⅰ
CONTENTS……….ⅳ
List of Figures………...………...ⅷ
List of Tables………....ⅸ
Chapter 1. General Introduction……….1
Chapter 2. Characterization of extended-spectrum β-lactamase
(ESBL)-producing Escherichia coli strains isolated from retail
raw chickens in Korea………3
2.1. INTRODUCTION……….4
2.2. MATERIALS & METHODS………...……….7
2.2.1. Collection of ESBL-producing E. coli strains from retail
chicken in Korea………...……….7
2.2.2. Antimicrobial susceptibility test………...8
2.2.3. Phylogenetic analysis of ESBL-producing E. coli
analysis………...9
2.2.5. Detection of virulence genes associated with pathogenic
E. coli……….9
2.2.6. Statistical analysis……...………...10
2.3. RESULTS………18
2.3.1. Distribution of ESBL genes in E. coli strains from retail
chicken……….18
2.3.2. Antimicrobial susceptibility of ESBL-producing E. coli
strains from retail chicken………21
2.3.3. Phylogenetic group analysis of ESBL-producing E. coli
strains from retail chicken………23
2.3.4. Prevalence of virulence genes in ESBL-producing E. coli
strains from retail chicken………26
2.4. DISCUSSION……….29
Chapter 3. Co-existence of plasmid-mediated quinolone
resistance (PMQR) genes in extended-spectrum β-lactamase
(ESBL) producing Escherichia coli strains from retail raw
3.1. INTRODUCTION………...34
3.2. MATERIALS & METHODS………..36
3.2.1. Bacterial strains and detection of PMQR and ESBL
genes………...………..36
3.2.2. Antimicrobial susceptibility testing and phylogenetic
grouping analysis……….36
3.2.3 Statistical analysis………...36
3.3. RESULTS………39
3.3.1. Prevalence of PMQR genes in ESBL-producing E. coli
strains isolated from retail raw chicken in
Korea………...………39
3.3.2. Antimicrobial resistance in PMQR and ESBL-positive
strains of E. coli………...……...41
3.4. DISCUSSION………..43
REFERENCES……….46
List of Figures
Fig. 2.1. Prevalence of ESBL genes in different ESBL
type………...19
Fig. 2.2. Phylogenetic analysis of 67 ESBL-producing E. coli
strains with RAPD-PCR………...24
List of Tables
Table 2.1. Bacterial strains used in this chapter………...11
Table 2.2. Primers used in this chapter………13
Table 2.3. Antibiotic resistance patterns of 67 ESBL-producing E.
coli strains depending on the ESBL gene type……….22
Table 2.4. Phylogenetic group of ESBL-producing E. coli isolates
from chicken……….25
Table 2.5. Prevalence of the virulence genes of E. coli isolates
according to phylogenetic group………..27
Table 2.6. Distribution of strains in the phylogenetic group
according to the number of virulence genes………28
Table 3.1. Primers used in this chapter………37
Table 3.2. Distribution of ESBL types in PMQR-positive E. coli
isolates (n=35)………..40
Table 3.3. Comparison of antimicrobial susceptibility between
qnrS positive strains and both qnrS and aac(6
′
)-Ib-cr positive
Chapter 1. General Introduction
Extended-spectrum β-lactamases (ESBLs) are a group of enzymes conferring resistance to most β-lactams, including third-generation cephalosporins(Rawat and Nair 2010). ESBLs have become widespread in a significant percentage of Gram-negative bacteria, especially in
Enterobacteriaceae throughout the world (Bradford 2001). Four enzyme
families, TEM-type, SHV-type, CTX-M-type, and OXA-type are currently regarded the most common ESBLs among Enterobacteriaceae (Bradford 2001). The emergence and spread of ESBLs is well recognized as a public health threat as they seriously compromised the efficacy of β-lactams, particularly extended-spectrum cephalosporins (Bonnet 2004, Canton, Novais et al. 2008). ESBLs are often encoded by genes located on large plasmids, which also often encode genes for resistance to other antimicrobial agents, such as aminoglycosides, trimethoprim, sulphonamides, tetracyclines and chloramphenicol (Paterson 2000).
Fluoroquinolones (FQs) are another antibiotic class of clinical importance since they have a broad spectrum antibacterial activity and are widely used in human and veterinary medicine. However, increasing resistance to FQs in Enterobacteriaceae is also a threat to the veterinary clinical therapy and human public health. Quinolone resistance is mediated
by chromosomal mutations in DNA gyrase and topoisomeraseIV (Huang, Dai et al. 2009) and plasmid-mediated quinolone resistance (PMQR) mechanisms. To date, at least three types of PMQR determinants, including
qnr families (qnrA, qnrB, qnrS), aac(6′)-Ib-cr and quinolone efflux pump
(qepA and oqxAB) have been extensively reported (Robicsek, Strahilevitz et al. 2006, Yamane, Wachino et al. 2007, Strahilevitz, Jacoby et al. 2009). These PMQR genes as well as ESBL genes are usually carried on plasmids and can be transferred among different bacterial strains and species (Strahilevitz, Jacoby et al. 2009) (Overdevest, Willemsen et al. 2011). Recent studies have demonstrated FQ resistance mediated by co-transfer of the PMQR genes on ESBL-encoding plasmids (Mammeri, Van De Loo et al. 2005, Guillard, Bertrand et al. 2014).
Through the food supply chain, humans are exposed to a large amount of bacterial flora originated from farms. Therefore, food and food-producing animals may potentially serve as a reservoir for zoonotic pathogens and may mediate the transfer of pathogens and their resistance genes from food-producing animals to humans (Madec and Haenni 2018). In this study, I investigated the prevalence of ESBL-producing E. coli in chickens across whole provinces in South Korea and characterized antibiotic
Chapter 2. Characterization of extended-spectrum
β-lactamase (ESBL)-producing Escherichia coli strains
2.1. INTRODUCTION
Enterobacteriaceae that produce extended-spectrum β-lactamases
(ESBLs), particularly Escherichia coli producing CTX-M, have emerged as significant antibiotic-resistant human pathogens across the world (Hawkey and Jones 2009). Since ESBL genes are typically encoded by mobile genetic elements, usually plasmids, ESBL genes may be easily disseminated (Pitout, Nordmann et al. 2005), and the mobility of ESBL genes has resulted in the rapid increase in the prevalence of ESBL-producing Enterobacteriaceae in food-producing and companion animals, various environmental samples (e.g., wastewater), and food (Iovleva and Bonomo 2017). ESBL-producing E. coli is frequently isolated from chicken meat (Ewers, Antão et al. 2009, Yuan, Liu et al. 2009, Overdevest, Willemsen et al. 2011, Olsen, Bisgaard et al. 2014, Nahar, Awasthi et al. 2018). In some cases, the high prevalence of ESBL-producing E. coli hampers the isolation of other fastidious bacteria (e.g.,
Campylobacter) from chicken as it outgrows during the enrichment step
using cephalosporins as a selective supplement (Hazeleger, Jacobs-Reitsma et al. 2016).
develops infections outside the intestines, ExPEC first colonizes the gastrointestinal tracts and is transmitted primarily by the consumption of food, particularly chicken (Nordstrom, Liu et al. 2013). Commensal E. coli isolates usually harbor no or only very few virulence genes; however, ExPEC possesses a broad range of virulence genes, such as adhesins, iron acquisition, serum survival, and toxins associated with extraintestinal disease (Russo and Johnson 2000). ExPEC commonly possesses large, transmissible, multidrug resistance (MDR) plasmids encoding ESBLs (Johnson, Logue et al. 2012), suggesting that chicken could be a source for both ExPEC and ESBL-producing E. coli.
A number of studies have shown that retail chicken is significantly involved in transmitting ESBL-producing E. coli and ExPEC to humans (Manges, Smith et al. 2007, Ewers, Antão et al. 2009, Johnson, Logue et al. 2012). Despite the public health importance, the prevalence of ESBL-producing E. coli on retail chicken in Korea has been reported only in a single study, which characterized only a limited number (n=6) of ESBL-producing E. coli from retail chicken (Jo and Woo 2016). Furthermore, no studies have performed to examine if ESBL-producing E. coli from retail chicken in Korea may have potential to cause extraintestinal infections in humans. To fill this important knowledge gap, in this study, I characterized the antibiotic resistance and virulence gene prevalence of ESBL-producing E.
2.2. MATERIALS & METHODS
2.2.1. Collection of ESBL-producing E. coli strains
from retail chicken in Korea
A total of 67 strains of ESBL-producing E. coli were isolated from 40 retail whole chicken samples from 28 companies in six different provinces of South Korea in our previous study (submitted). E. coli strains were grown on MacConkey agar and were confirmed with 16S rRNA sequencing (Macrogen, Korea). To confirm the ESBL phenotype, the E. coli strains were subjected to the modified ESBL confirmatory test of the Clinical and Laboratory Standards Institute (CLSI), which uses antimicrobial disks of cefotaxime (CTX) and ceftazidime (CAZ) with or without clavulanic acid (CA), an ESBL inhibitor, and boric acid (BA) and EDTA to inhibit AmpC and carbapenemases, respectively (Poulou, Grivakou et al. 2014). E. coli ATCC 25922, a CLSI quality control strain, was used as a negative control. The presence of the ESBL genes was PCR-determined using primers to detect all types of ESBL (SHV, TEM, CTX-M, and OXA) (Fang, Ataker et al. 2008). The PCR amplicons were sequenced, and the translated amino acid sequences were used to determine the ESBL gene type (Kim, Lim et al. 2005, Costa, Poeta et al. 2006). The E. coli strains were routinely cultured on Luria-Bertani (LB) media.
2.2.2. Antimicrobial susceptibility test
The antimicrobial susceptibility of the strains was determined with a broth dilution method using a total of 12 antibiotics, including ampicillin, cephalothin, tetracycline, chloramphenicol, kanamycin, gentamicin, streptomycin, polymyxin B, colistin, ceftriaxone, cefotaxime, and ciprofloxacin. The minimum inhibitory concentrations (MICs) were determined according to the CLSI guidelines and a previous report (Bell, Gatus et al. 1999, Institute 2017). E. coli ATCC 25922 was used as a quality control strain according to the CLSI protocol.
2.2.3 Phylogenetic analysis of ESBL-producing E.
coli strains
The phylogenetic group was determined using a multiplex PCR-based method amplifying chuA, yjaA, and DNA fragment TspE4C2 (Clermont, Bonacorsi et al. 2000). E. coli strains MG 1655 and ATCC 25922 were used as controls for the phylogenetic group A and B2, respectively.
2.2.4 Random amplified polymorphic DNA (RAPD)
analysis
RAPD was used as a PCR-based DNA fingerprinting method to analyze clonal similarity. Extracted DNA from 67 strains of E. coli were distinguished with a single primer, the arbitrary primer 1254, introduced in the previous report (Madico, Akopyants et al. 1995). The PCR results were analyzed with BioNumerics (ver. 7; Applied Maths).
2.2.5 Detection of virulence genes associated with
pathogenic E. coli
PCR was performed to analyze the presence of virulence genes associated with five major intestinal pathogenic E. coli groups, including Shiga-toxin producing E. coli (stx1 and stx2 encoding Shiga toxins, hylA encoding enterohemolysin, and espP encoding serine protease), enteropathogenic (eaeA encoding intimin), enterotoxigenic (st and lt encoding heat-stable and heat-labile enterotoxins), and enteroaggregative (aggR encoding a transcription regulator for aggregative adherence fimbria I), and ExPEC (Paton and Paton 1998, Cerna, Nataro et al. 2003). The tested virulence genes related to ExPEC include extraintestinal E. coli attachment
factors (fimH, iha), iron uptake (aer, irp2, iutA), iron transporter (feoB), serum survival (iss), and heat stable enterotoxin (astA) (Dezfulian, Batisson et al. 2003, Jakobsen, Spangholm et al. 2010, Guastalli, Guastalli et al. 2013, Lee, Subhadra et al. 2016). E. coli ATCC 35150 was used as a positive control for STEC and EPEC. E. coli NCCT 14039 was a positive control for EAEC. E. coli MG 1655 was used as a negative control for the tested virulence genes except for fimH and feoB. E. coli strains ATCC 43888 and O169 were used as positive controls for iha and astA, respectively. E. coli strains MG 1655 and ATCC 25922 were used as positive and negative controls, respectively, for fimH and feoB.
2.2.6 Statistical analysis
The statistical significance of the distribution of virulence genes was analyzed statistically with a chi-square test using GraphPad Prism (ver.5; GraphPad Software)
Table 2.1. Bacterial strains used in this chapter
Strains Characteristics References
E. coli
ATCC 25922
Phylogenetic group B2
Quality control strain for MICatest
Laboratory collection
MG 1655
Phylogenetic group A
Positive control for fimH and feoB
Negative control for stx1, stx2, hylA, eaeA, espP,
aggR, iha, astA, iss, aer, irp2, iutA, st, and lt
Laboratory collection
ATCC 35150
Positive control for stx1, stx2, hylA, eaeA, and
espP
NCTC 14039 Positive control for aggR Laboratory collection
ATCC 43888 Positive control for iha Laboratory collection
O169 Positive control for astA Laboratory collection
Table 2.2. Primers used in this chapter
Gene Primers used Amplicon
size (bp)
References ESBL gene
blaTEM F : CGC CGC ATA CAC TAT TCT CAG AAT
GA
R : ACG CTC ACC GGC TCC AGA TTT AT
445 (Fang, Ataker et al. 2008)
blaSHV F : CTT TAT CGG CCC TCA CTC AA
R : AGG TGC TCA TCA TGG GAA AG 237 (Fang, Ataker et al. 2008)
blaCTX-M F : ATG TGC AGY ACC AGT AAR GTK ATG
GC
R : TGG GTR AAR TAR GTS ACC AGA AYC AGC GG
blaOXA F : ACA CAA TAC ATA TCA ACT TCG C
R : AGT GTG TTT AGA ATG GTG ATC
813 (Fang, Ataker et al. 2008)
Phylogenetic grouping
chuA F : GAC GAA CCA CGG TCA GGA T R : TGC CGC CAG TAC C AA AGA CA
279
(Clermont, Christenson et al. 2013)
yjaA F : TGA AGT GTC AGG AGA CGC TG R : ATG GAG AAT GCG TTC CTC AAC
211
(Clermont, Christenson et al. 2013)
DNA fragment TspE4.C2.
F : GAG TAA TGT CGG GGC ATT CA
R : CGC GCC AAC AAA GTA TTA CG 152
(Clermont, Christenson et al. 2013)
arpA F : AAC GCT ATT CGC CAG CTT GC
1254 CCG CAG CCA A Random
(Madico, Akopyants et al. 1995)
Virulence gene
stx1 F : ACC TCA CTG ACG CAG TCT GTG G
R : TCT GCC GGA CAC ATA GAA GGA AA 350 (Paton and Paton 1998)
stx2 F : ACT GTC TGA AAC TGC TCC TGT G
R : TTA TTT TTA TAA CGG GCC TGT TCG C 262 (Paton and Paton 1998)
hylA F : GCA TCA TCA AGC GTA CGT TCC
R : AAT GAG CCA AGC TGG TTA AGC T 534 (Paton and Paton 1998)
espP F : AAA CAG CAG GCA CTT GAA CG R : GGA GTC GTC AGT CAG TAG AT
1830
(Dezfulian, Batisson et al. 2003)
eaeA F : GAC CCG GCA CAA GCA TAA GC R : CCA CCT GCA GCA ACA AGA GG
aggR F : CTA ATT GTA CAA TCG ATG TA R : AGA GTC CAT CTC TTT GAT AAG
457 (Cerna, Nataro et al. 2003)
iha F : CTG GCG GAG GCT CTG AGA TCA R : TCC TTA AGC TCC CGC GGC TGA
827 (Dezfulian, Batisson et al. 2003)
astA F : TGC CAT CAA CAC AGT ATA TCC R : TCA GGT CGC GAG TGA CGG C
116 (Guastalli, Guastalli et al. 2013)
iss F : ATC ACA TAG GAT TCT GCC G R : CAG CGG AGT ATA GAT GCC A
309 (Guastalli, Guastalli et al. 2013)
aer F : AAA CCT GGC TTA CGC AAC TGT R : ACC CGT CTG CAA ATC ATG GAT
269 (Lee, Subhadra et al. 2016)
irp2 F : AAG GAT TCG CTG TTA CCG GAC
R : CGC ATT TAC CGT CGG GAA CGG 2010)
fimH F : TGC AGA ACG GAT AAG CCG TGG R : GCA GTC ACC TGC CCT CCG GTA
508 (Lee, Subhadra et al. 2016)
feoB F : AAT TGG CGT GCA TGA AGA TAA CTG R : AGC TGG CGA CCT GAT AGA ACA ATG
470 (Lee, Subhadra et al. 2016)
st F : TCT TTC CCC TCT TTT AGT CAG TC R : CCA GCA CAG GCA GGA TTA C
170 In this study
lt F : TCT CTA TGT GCA CAC GGA GC R : CCA TAC TGA TTG CCG CAA T
2.3 RESULTS
2.3.1 Distribution of ESBL genes in E. coli strains
from retail chicken
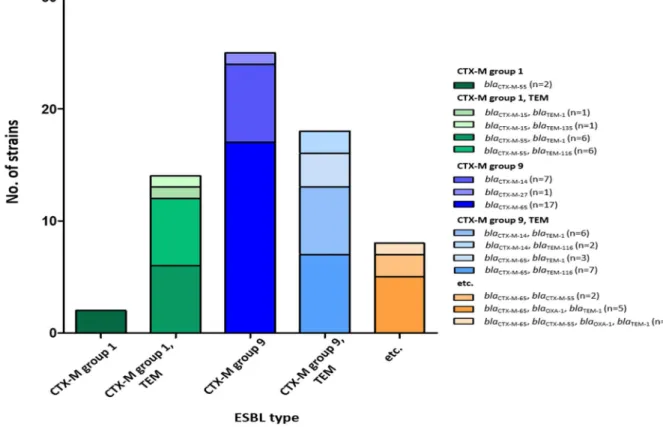
All the ESBL-producing E. coli strains from retail poultry possessed
blaCTX-M (Fig. 2.1), suggesting that CTX-M was the predominant ESBL type
in E. coli from retail chicken in Korea (Fig. 2.1). Approximately 56.7% of the blaCTX-M-positive strains also carried blaTEMand/or blaOXA(Fig. 2.1), and
blaSHVwas not detected in the strains (Fig. 2.1). The dominant CTX-M types
included CTX-M-65 (52.2%) in the CTX-M group 9 and CTX-M-55 (25.4%) in the CTX-M group 1, and three strains harbored both blaCTX-M-65 and
blaCTX-M-55(Fig. 2.1). The blaOXA-1 gene was detected in six (9.0%) of the 67
strains of ESBL-producing E. coli, and the blaOXA-1-positive strains also
Four types of ESBL genes (blaCTX-M, blaTEM, blaOXA, and blaSHV) were detected by PCR. ATCC 25922 was used as a
negative control. The PCR products were sequenced commercially (Macrogen, Korea), and the nucleotide sequences were analyzed with BLAST.
2.3.2 Antimicrobial susceptibility of ESBL-producing
E. coli strains from retail chicken
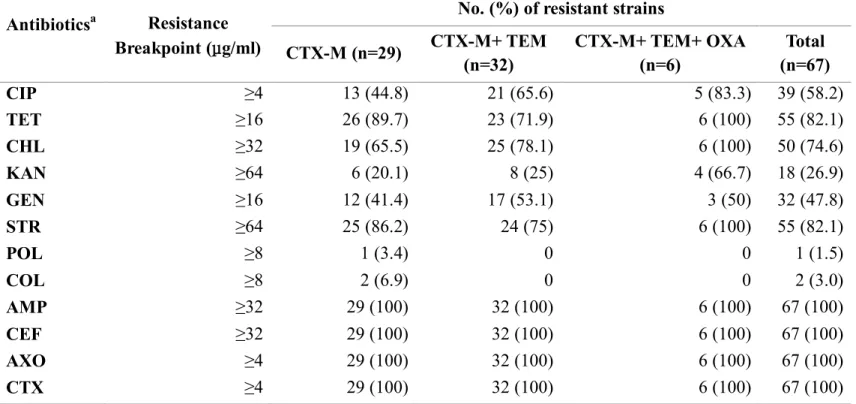
All E. coli strains carrying blaCTX-Mwere resistant to cephalosporins
of different generations, such as cephalothin (1stgeneration), ceftriaxone (3rd generation), and cefotaxime (3rd generation) (Table 2.3). The strains harboring all of the blaCTX-M, blaTEM, and blaOXA genes were highly resistant
to all tested antibiotics except for polymyxin B and colistin (Table 2.3). The results showed that 98.5% (66/67) of the tested ESBL-producing E. coli strains were resistant to ≥ 3 antibiotic classes tested in this study, suggesting that ESBL-producing E. coli on retail chicken in Korea is highly multidrug-resistant.
Table 2.3. Antibiotic resistance patterns of 67 ESBL-producing E. coli strains depending on the ESBL gene type
a12 antimicrobial agents used include CIP: ciprofloxacin, AMP: ampicillin, CEF: cephalothin, TET: tetracycline, CHL: Antibioticsa Resistance
Breakpoint (μg/ml)
No. (%) of resistant strains CTX-M (n=29) CTX-M+ TEM (n=32) CTX-M+ TEM+ OXA (n=6) Total (n=67) CIP ≥4 13 (44.8) 21 (65.6) 5 (83.3) 39 (58.2) TET ≥16 26 (89.7) 23 (71.9) 6 (100) 55 (82.1) CHL ≥32 19 (65.5) 25 (78.1) 6 (100) 50 (74.6) KAN ≥64 6 (20.1) 8 (25) 4 (66.7) 18 (26.9) GEN ≥16 12 (41.4) 17 (53.1) 3 (50) 32 (47.8) STR ≥64 25 (86.2) 24 (75) 6 (100) 55 (82.1) POL ≥8 1 (3.4) 0 0 1 (1.5) COL ≥8 2 (6.9) 0 0 2 (3.0) AMP ≥32 29 (100) 32 (100) 6 (100) 67 (100) CEF ≥32 29 (100) 32 (100) 6 (100) 67 (100) AXO ≥4 29 (100) 32 (100) 6 (100) 67 (100) CTX ≥4 29 (100) 32 (100) 6 (100) 67 (100)
2.3.3 Phylogenetic group analysis of ESBL-producing
E. coli strains from retail chicken
The predominant phylogenetic group of the total of 67 ESBL-producing E. coli strains from retail chicken was the group A (56.7%) and group B1 (19.4%). Four strains of E. coli belonged to the group B2 and group D, respectively. Eight strains were classified as minor group; 6 out of 8 strains belonged to the group E, one strain to group F, and one strain unknown (Table 2.4). The phylogenetic analysis using RAPD-PCR showed that ESBL-producing E. coli strains belonging to the same phylogenetic group tended to form the same cluster (Fig. 2.2).
Fig 2.2. Phylogenetic analysis of 67 ESBL-producing E. coli strains with RAPD-PCR
RAPD-Table 2.4. Phylogenetic group of ESBL-producing E. coli isolates from
chicken
Phylogenetic group No. (%) of strains
A 38 (56.7) B1 13 (19.4) B2 4 (6.0) D 4 (6.0) E 6 (8.9) F 1 (1.5) Unknown 1 (1.5) Total 67
2.3.4 Prevalence of virulence genes in
ESBL-producing E. coli strains from retail chicken
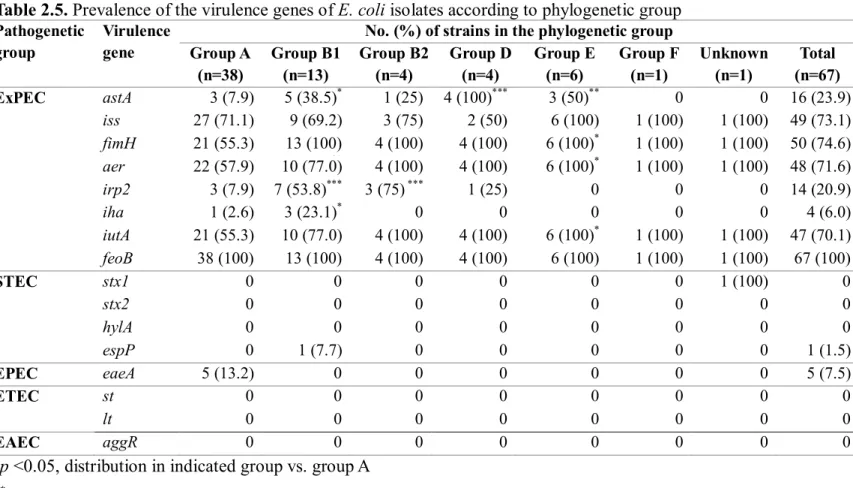
We examined the prevalence of the virulence genes that represent the five major pathogenic groups of E. coli, including enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), enteroaggregative E. coli (EAEC), Shiga toxin-producing E. coli (STEC), and ExPEC. The espP and eaeA genes were detected in one strain in the group B1 and five strains in the group A, respectively, and toxin genes were not detected (Table 2.5). Interestingly, all of the 67 strains carried at least one ExPEC-related virulence genes (Table 2.5). Compared to the group A, the prevalence of ExPEC-related virulence genes was more frequently detected in the groups B1, B2, D, and E (Table 2.5), and those harboring more than six ExPEC-related virulence genes belonged to the phylogenetic groups B1, B2, D, and E with a statistical significance (Table 2.6).
Table 2.5. Prevalence of the virulence genes of E. coli isolates according to phylogenetic group
Pathogenetic group
Virulence gene
No. (%) of strains in the phylogenetic group Group A (n=38) Group B1 (n=13) Group B2 (n=4) Group D (n=4) Group E (n=6) Group F (n=1) Unknown (n=1) Total (n=67) ExPEC astA 3 (7.9) 5 (38.5)* 1 (25) 4 (100)*** 3 (50)** 0 0 16 (23.9) iss 27 (71.1) 9 (69.2) 3 (75) 2 (50) 6 (100) 1 (100) 1 (100) 49 (73.1) fimH 21 (55.3) 13 (100) (100.0)** 4 (100) 4 (100) 6 (100)* 1 (100) 1 (100) 50 (74.6) aer 22 (57.9) 10 (77.0) 4 (100) 4 (100) 6 (100)* 1 (100) 1 (100) 48 (71.6) irp2 3 (7.9) 7 (53.8)*** 3 (75)*** 1 (25) 0 0 0 14 (20.9) 5 () iha 1 (2.6) 3 (23.1)* 0 0 0 0 0 4 (6.0) iutA 21 (55.3) 10 (77.0) 4 (100) 4 (100) 6 (100)* 1 (100) 1 (100) 47 (70.1) feoB 38 (100) (100.0) 13 (100) 4 (100) 4 (100) 6 (100) 1 (100) 1 (100) 67 (100) (100.0) STEC stx1 0 0 0 0 0 0 1 (100) 0 stx2 0 0 0 0 0 0 0 0 hylA 0 0 0 0 0 0 0 0 espP 0 1 (7.7) 0 0 0 0 0 1 (1.5) EPEC eaeA 5 (13.2) 0 0 0 0 0 0 5 (7.5) ) ETEC st 0 0 0 0 0 0 0 0 lt 0 0 0 0 0 0 0 0 EAEC aggR 0 0 0 0 0 0 0 0
*p <0.05, distribution in indicated group vs. group A **p <0.01, distribution in indicated group vs. group A ***p <0.001, distribution in indicated group vs. group A
Table 2.6. Distribution of strains in the phylogenetic group according to the number of virulence genes
*p <0.05, distribution in indicated group vs. group A **p <0.01, distribution in indicated group vs. group A ***p <0.001, distribution in indicated group vs. group A
Phylogenetic group No. (%) of virulence genes
7 6 5 4 3 2 1 Group A (n=38) 0 0 7 (18.4) 15 (39.5) 11 (28.9) 3 (7.9) 2 (5.3) Group B1 (n=13) 3 (23.1)** 5 (38.5)*** 2 (15.4) 0** 3 (23.1) 0 0 Group B2 (n=4) 1 (2.5)** 1 (2.5)** 2 (50) 0 0 0 0 Group D (n=4) 1 (2.5)** 1 (2.5)** 2 (50) 0 0 0 0 Group E (n=6) 0 3 (50)*** 3 (50) 0 0 0 0 Group F (n=1) 0 0 1 (100)* 0 0 0 0 Unknown (n=1) 0 0 1 (100)* 0 0 0 0
2.4 DISCUSSION
ESBL-producing E. coli has been frequently isolated from poultry (Olsen, Bisgaard et al. 2014), and retail chicken is considered as an important vehicle transmitting ESBL-producing E. coli to humans (Overdevest, Willemsen et al. 2011). Consistent with the global expansion of CTX-M ESBLs (Cantón, González-Alba et al. 2012), CTX-M was the predominant ESBL type in E. coli strains from retail chicken in Korea (Fig. 2.1). In this study, blaCTX-M-65 (52.2%), blaCTX-M-55 (25.4%), and blaCTX-M-14
(22.4%) were the most common ESBL genes (Fig. 2.1). Similarly, bla CTX-M-55, blaCTX-M-65, and blaCTX-M-14 were commonly detected in ESBL-producing
E. coli strains from chicken in China (Zheng, Zeng et al. 2012). In Japan, blaCTX-M-2, blaTEM, and blaCTX-M-1 were present in 45%, 36%, and 34% of
ESBL-producing E. coli strains from domestic retail chicken meat samples, respectively (Nahar, Awasthi et al. 2018). In Netherlands, approximately 94% of retail chicken samples were positive for ESBL-producing E. coli, and
blaCTX‐M‐1 was predominant (49%) in the ESBL-producing E. coli
(Leverstein‐van Hall, Dierikx et al. 2011). Similarly, 65.4% of chicken meat samples in the UK were contaminated with ESBL-producing E. coli, and
blaCTX‐M‐1 was predominant (82.7%) in ESBL-positive E. coli strains from
others, ESBL-producing E. coli is highly prevalent on retail chicken in most countries, but dominant ESBL gene types are different depending on the country.
All of the ESBL-producing E. coli strains from retail chicken in this study were resistant to multiple drugs belonging to different classes, such as ampicillin, tetracycline, chloramphenicol, and streptomycin (Table 2.3). The high prevalence of MDR ESBL-producing E. coli has been reported in previous reports. In China, 80.6% of ESBL-producing E. coli strains were resistant to at least three different antibiotic classes (Yuan, Liu et al. 2009). MDR strains were highly distributed in clinical samples as well as chicken samples. Approximately, 76.6% of the ESBL-producing E. coli isolated from hospitals in Malaysia were MDR (Lim, Yasin et al. 2009). Two strains were resistant to colistin, one of the last-resort antibiotics (Table 2.3). Further investigation found that the strains are positive for mcr-1 (data not shown), the plasmid-encoded gene conferring resistance to colistin (Liu, Wang et al. 2016). This observation may be explained by the fact that ESBL genes are usually encoded on transmissible plasmids that harbor multiple resistance genes (Paterson 2000). The strains carrying three different ESBL genes (i.e.,
(Table 2.3). Presumably, the co-existence of different ESBL genes may result from the co-presence of multiple resistance plasmids in the same strain. For instance, it has been demonstrated that plasmid-mediated quinolone resistance genes are frequently detected in ESBL-producing E. coli isolates from poultry (Xia, Tao et al. 2011).
Virulent ExPEC strains usually belong to phylogenetic groups B2 and D (Picard, Garcia et al. 1999, Cortés, Blanc et al. 2010). However, in our study, ESBL-producing E. coli strains isolated from retail chicken mainly belonged to the group A (56.7%) and group B1 (19.4%). And only four strains were assigned to each group of B2 and D (Table 3). Interestingly, the number of strains in the group E, which is classified as minor group, were larger than those belonging to the group B2 and D (Table 3). E. coli O157:H7 EDL933 is the best-known member belonging to the Group E (Clermont, Christenson et al. 2013). Consistently, a previous study in Jeonnam Province in Korea showed that the majority of E. coli isolates from chicken were localized to phylogenetic group A, followed by group B1, and no isolate belonged to the group B2 (Unno, Han et al. 2009). A similar pattern of distribution was also found among isolates from chicken carcasses in China (Xu, Cui et al. 2014). In contrast, E. coli isolates from chicken meat had the greatest percentage of group B1 strains (44%), followed by group A (28%) and D (23%) in Netherland (Kluytmans, Overdevest et al. 2012). E.
coli isolates from chicken meat in the United States, groups D, B2, A, and
B1 were dominant as given order (Mitchell, Johnson et al. 2015). Thus, the distribution of phylogenetic groups may be affected by their geographical region. However, the prevalence of strains belonging to the group E in our study was similar to a previous report from Netherland (Kluytmans, Overdevest et al. 2012). Whereas the isolates harbored no or only very few virulence factors related to gastrointestinal infection, various virulence factors associated with ExPEC were frequently detected in ESBL-producing
E. coli strains, particularly in those belonging to groups B1, B2, D, and E
except group A (Tables 4 and 5). These results suggest that ESBL-producing
E. coli strains in groups B1, B2, D, and E from retail chicken may potentially
be implicated in extraintestinal infections in humans in Korea.
The findings in this study demonstrated that retail chicken in Korea is a major source of disseminating ESBL-producing E. coli strains with a great potential to cause extraintenstinal infections. Since the ESBL-producing E. coli strains from chicken are often MDR, further investigation is required to control and reduce the contamination of chicken meat by ESBL-producing E. coli to protect public health.
Chapter 3. Co-existence of plasmid-mediated
quinolone resistance (PMQR) genes in
extended-spectrum β-lactamase (ESBL) producing Escherichia
3.1 INTRODUCTION
Fluoroquinolones (FQs) and 3rd generation cephalosporins are among the World Health Organization (WHO) list of critically important antibiotics for human medicine due to their common use in both humans and animals (Organization 2017).
FQ resistance is mediated by multiple mechanisms, including spontaneous mutations in genes encoding the cellular targets of FQ (e.g., DNA gyrase and topoisomeraseIV) (Hopkins, Davies et al. 2005) and plasmid-mediated quinolone resistance (PMQR) genes, such as the qnr genes whose protein products protect DNA gyrase and topoisomerase IV from quinolones, aac(6′)-Ib-cr encoding a variant aminoglycoside acetyltransferase that inactivates quinolones by acetylation, and oqxAB and qepA encoding efflux pumps that extrude quinolones (Strahilevitz, Jacoby et al. 2009). Whereas the antibiotic selection pressure plays a role in enriching FQ-resistant bacteria with spontaneous mutations (Luangtongkum, Jeon et al. 2009), PMQR genes can be easily transmitted horizontally and cause clinical problems by reducing FQ susceptibility (Molbak 2005, Strahilevitz, Jacoby et al. 2009).
cephalosporins (Rawat and Nair 2010). The increasing rate of antimicrobial resistance has shown that the detection rate of PMQR genes and ESBLs has increased significantly not only in human clinical Escherichia coli isolates (Briales, Rodríguez-Martínez et al. 2012, Park, Kim et al. 2012), but also in
E. coli isolates from food-producing animals and food products, which has
been raising concern about their possible transmission through food chain (Yue, Jiang et al. 2008). As food and food-producing animals are a major reservoir for plasmid-encoded resistance genes, the resistance is likely to be transmitted even by international trade of food (Madec and Haenni 2018).
It has been reported that PMQR genes are frequently associated with ESBL-producing E. coli (Guillard, Bertrand et al. 2014). In South Korea, ESBL-producing clinical isolates carrying PMQR genes have also been reported (Kim, Jeong et al. 2009, Park, Kim et al. 2012); however, nothing has been reported about the prevalence of PMQR genes in ESBL-producing
E. coli isolates from food in this country. In this study, we investigated the
prevalence of PMQR genes in ESBL-positive E. coli strains isolated from retail raw chickens in South Korea.
3.2 MATERIALS & METHODS
3.2.1 Bacterial strains and detection of PMQR and
ESBL genes
A total of 67 ESBL-producing E. coli strains were isolated from retail raw chickens in South Korea from June to August 2017 in the previous study (Chapter 2). Species identification and screening for ESBL type were also done in the previous study (Chapter 2). All isolates were PCR-screened for PMQR genes (qnrA, qnrB, qnrS, aac(6′)-Ib-cr, qepA, oqxA, and oqxB) using primers as described previously (Yamane, Wachino et al. 2008, Yue, Jiang et al. 2008, Kim, Wang et al. 2009). E. coli ATCC 25922 was used as a negative control. The PCR products were subjected to direct sequencing (Macrogen, Korea).
3.2.2 Statistical analysis
The statistical significance of the distribution of antibiotics resistant strains was analyzed statistically with a chi-square test using GraphPad Prism (ver.5; GraphPad Software)
Table 3.1. Primers used in this chapter
Gene Primers used
Amplicon
size (bp) References PMQR genes
qnrA F : TCA GCA AGA GGA TTT CTC A R : GGC AGC ACT ATT ACT CCC A
627 (Yue, Jiang et al. 2008)
qnrB F : GAT CGT GAA AGC CAG AAA GG
R : ACG ATG CCT GGT AGT TGT CC 469 (Yue, Jiang et al. 2008)
qnrS F : ACG ACA TTC GTC AAC TGC AA
R : TAA ATT GGC ACC CTG TAG GC 417 (Yue, Jiang et al. 2008)
aac(6′)-Ib-cr F : TTG CGA TGC TCT ATG AGT GGC TA R : CTC GAA TGC CTG GCG TGT TT
482 (Yue, Jiang et al. 2008)
R : CCA CTC TTC ACG GGA GAC GA
oqxB F : TTC TCC CCC GGC GGG AAG TAC R : CTC GGC CAT TTT GGC GCG TA
512 (Kim, Wang et al. 2009)
qepA F : GCA GGT CCA GCA GCG GGT AG R : CTT CCT GCC CGA GTA TCG TG
3.3 RESULTS
3.3.1 Prevalence of PMQR genes in ESBL-producing
E. coli strains isolated from retail raw chicken in
Korea
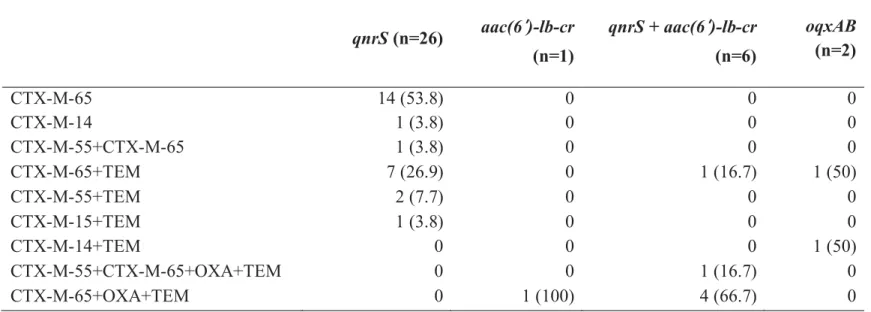
The distribution of PMQR-encoding genes among the 67 ESBL-producing E. coli isolates was shown in Table 3.2. Approximately 52.2% (35/67) of the isolates harbored at least one of qnrS, aac(6′)-Ib-cr, and
oqxAB in the ESBL-producing E. coli strains; however, qnrA, qnrB, and qepA were not detected (Table 3.2). The most prevalent PMQR gene was qnrS, which was detected in 26 (74.3%) out of the 35 PMQR-positive strains,
and six strains harbored both qnrS and aac(6′)-Ib-cr (Table 3.2). Among the
qnrS-positive strains, 22 (84.6%, 22/26) strains co-harbored blaCTX-M-65, of
which seven (26.9%, 7/26) and one (3.8%, 1/26) strains also carried blaTEM
and blaCTX-M-55, respectively (Table 3.2). The blaOXA-1 gene was detected in
six of seven aac(6′)-Ib-cr-positive strains (Table 3.2). The results showed that PMQR genes are highly prevalent in ESBL-producing E. coli strains isolated from retail chicken in Korea and that qnrS was predominant.
Table 3.2. Distribution of ESBL types in PMQR-positive E. coli isolates (n=35)
ESBL type No. (%) of strains carrying PMQR genes
qnrS (n=26) aac(6′)-lb-cr (n=1) qnrS + aac(6′)-lb-cr (n=6) oqxAB (n=2) CTX-M-65 14 (53.8) 0 0 0 CTX-M-14 1 (3.8) 0 0 0 CTX-M-55+CTX-M-65 1 (3.8) 0 0 0 CTX-M-65+TEM 7 (26.9) 0 1 (16.7) 1 (50) CTX-M-55+TEM 2 (7.7) 0 0 0 CTX-M-15+TEM 1 (3.8) 0 0 0 CTX-M-14+TEM 0 0 0 1 (50) CTX-M-55+CTX-M-65+OXA+TEM 0 0 1 (16.7) 0 CTX-M-65+OXA+TEM 0 1 (100) 4 (66.7) 0
3.3.2
Antimicrobial resistance in PMQR and
ESBL-positive strains of E. coli
Comparison of the antimicrobial susceptibility of strains carrying only qnrS and strains carrying both qnrS and aac(6′)-Ib-cr is shown in Table 3.3. Both groups of strains showed high resistance towards ampicillin, tetracyclin, streptomycin, ceftriaxone, cefotaxime, and cephalothin (Table 3.3). The group of strains carrying both qnrS and aac(6′)-Ib-cr showed slightly increased MIC for chloramphenicol and gentamycin. In particular, they showed significantly difference in resistance to ciprofloxacin (100%, 6/6) and kanamycin (83.3%, 5/6) (Table 3.3) compared only qnrS harboring strains. In addition, this result showed that all isolates exhibited resistance to third-generation cephalosporins and suggested that an additive effect for
aac(6′)-Ib-cr on the MIC for antibiotics in FQ group and aminoglycoside
Table 3.3. Comparison of antimicrobial susceptibility between qnrS positive strains and both qnrS and
aac(6′)-Ib-cr positive strains
Abbreviation; MIC, minimum inhibitory concentration; S, susceptible; I, intermediate; R, resistant.
Antimicrobial agentsa
MIC range (μg/ml)
No. (%) of strains carrying
qnrS (n=26)
No. (%) of strains carrying both qnrS and aac(6′)-lb-cr (n=6)
S I R S I R S I R Ciprofloxacin ≤1 2 ≥4 3 (11.5) 15 (57.7) 8 (30.8) 0 0 6 (100)** Ampicillin ≤8 16 ≥32 0 0 26 (100) 0 0 6 (100) Tetracyclin ≤4 8 ≥16 2 (7.7) 0 24 (92.3) 0 0 6 (100) Chloramphenicol ≤8 16 ≥32 7 (26.9) 1 (3.8) 18 (69.2) 0 0 6 (100) Kanamycin ≤16 32 ≥64 21 (80.8) 0 5 (19.2) 1 (16.7) 0 5 (83.3)** Gentamicin ≤4 8 ≥16 16 (61.5) 0 10 (38.5) 3 (50) 0 3 (50) Streptomycin ≤16 32 ≥64 0 1 (3.8) 25 (96.2) 0 0 6 (100) Polymyxin B ≤2 4 ≥8 4 (15.4) 21 (80.8) 1 (3.8) 1 (16.7) 5 (83.3) 0 Colistin ≤2 4 ≥8 11 (42.3) 13 (50) 2 (7.7) 3 (50) 3 (50) 0 Ceftriaxone ≤1 2 ≥4 0 0 26 (100) 0 0 6 (100) Cefotaxime ≤1 2 ≥4 0 0 26 (100) 0 0 6 (100) Cephalothin ≤8 16 ≥32 0 0 26 (100) 0 0 6 (100)
3.4 DISCUSSION
PMQR genes may facilitate the spread of quinolone-resistant bacteria due to the mobility of resistance plasmids (Robicsek, Jacoby et al. 2006). In this study, approximately 52.2% (35/67) of ESBL-producing E.
coli strains from retail chicken in Korea harbored at least one of qnrS, aac(6′)-Ib-cr, and oqxAB in ESBL-producing E. coli strains; however, qnrA, qnrB, and qepA were not detected (Table 3.2). The qnrS gene was detected
predominantly in 91.4% of ESBL-producing E. coli strains, which is 74.3% of PMQR & ESBL-positive strains, and mostly (i.e., 84.6%; 22/26) in ESBL-producing E. coli strains harboring blaCTX-M-65 (Table 3.2). The
blaOXA-1 gene was detected in six of seven aac(6′)-Ib-cr-positive strains
(Table 3.2). The results showed that PMQR genes are highly prevalent in ESBL-producing E. coli strains isolated from retail chicken in Korea and that
qnrS was predominant.
Although a number of studies have shown that PMQR and ESBL genes are frequently co-localized in the same strain, the dominant PMQR and ESBL genes are different depending on the sample type and the geographical location. PCR screening of human clinical strains of E. coli and
Klebsiella pneumoniae in China showed that qnrA, qnrB, and qnrS genes
aac(6′)-Ib-cr, and ESBL genes were always located on the same plasmids (Jiang,
Zhou et al. 2008). Another study from China screening PMQR and ESBL genes in 202 chicken isolates of E. coli reported that approximately 37.1% isolates dominantly possessed qnrS and aac(6′)-Ib-cr (Li, Wang et al. 2014), which is similar to our findings (Table 3.2).
More than two PMQR genes were detected in the same strain (Table 3.2). Particularly, six strains harbored both aac(6′)-Ib-cr and qnrS, whereas only one strain carried only aac(6′)-Ib-cr, and six of seven aac(6′)-Ib-cr-positive isolates co-produced blaOXA-1 as well as blaCTX-M-65 (Table 3.2).
Compared to the strains carrying only qnrS, interestingly, the strains with both qnrS and aac(6′)-Ib-cr exhibited increased resistance to ciprofloxacin (100%, 6/6) and kanamycin (83.3%, 5/6) with a slight increase in the MICs of chloramphenicol and gentamycin (Table 3.3), suggesting an additive effect of simultaneous presence of qnrS and aac(6′)-Ib-cr on FQ and aminoglycoside resistance. Similarly, the co-localization of qnrS and
aac(6′)-Ib-cr in Klebsiella pneumoniae significantly increased resistance compared
to the strain carrying aac(6′)-Ib-cr alone (Kim, Jeong et al. 2009). This would be because aac(6′)-Ib-cr may reduce susceptibility to aminoglycosides as well as FQs (Ma, Zeng et al. 2009) and/or the co-existence of both qnrS
In summary, this study first showed the prevalence of PMQR genes in ESBL-producing E. coli strains isolated from retail chicken in Korea and that a co-localization of multiple resistance genes (i.e., qnrS and aac(6′)-Ib-cr) may further affect antibiotic resistance.
REFERENCES
Bell, S., et al. (1999). "Antibiotic susceptibility testing by the CDS method." A concise laboratory manual. Arthur Productions Pty Ltd, Sydney. NSW: The antibiotic reference laboratory, South Eastern Area Laboratory Services.
Bonnet, R. (2004). "Growing group of extended-spectrum β-lactamases: the CTX-M enzymes." Antimicrobial agents and chemotherapy 48(1): 1-14.
Bradford, P. A. (2001). "Extended-spectrum β-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat." Clinical microbiology reviews 14(4): 933-951.
Briales, A., et al. (2012). "Prevalence of plasmid-mediated quinolone resistance determinants qnr and aac (6′)-Ib-cr in Escherichia coli and Klebsiella pneumoniae producing extended-spectrum β-lactamases in Spain." International journal of antimicrobial agents 39(5): 431-434.
Cantón, R., et al. (2012). "CTX-M enzymes: origin and diffusion." Frontiers in microbiology 3: 110.
Canton, R., et al. (2008). "Prevalence and spread of extended‐spectrum β‐ lactamase‐producing Enterobacteriaceae in Europe." Clinical Microbiology and Infection 14: 144-153.
Cerna, J. F., et al. (2003). "Multiplex PCR for detection of three plasmid-borne genes of enteroaggregative Escherichia coli strains." Journal of clinical microbiology 41(5): 2138-2140.
Clermont, O., et al. (2013). "The Clermont Escherichia coli phylo‐typing method revisited: improvement of specificity and detection of new phylo‐ groups." Environmental microbiology reports 5(1): 58-65.
Cortés, P., et al. (2010). "Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain." Applied and environmental microbiology 76(9): 2799-2805.
Dezfulian, H., et al. (2003). "Presence and characterization of extraintestinal pathogenic Escherichia coli virulence genes in F165-positive E. coli strains isolated from diseased calves and pigs." Journal of clinical microbiology 41(4): 1375-1385.
Ewers, C., et al. (2009). "Intestine and environment of the chicken as reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential." Applied and environmental microbiology 75(1): 184-192.
Fang, H., et al. (2008). "Molecular epidemiology of extended-spectrum β-lactamases among Escherichia coli isolates collected in a Swedish hospital and its associated health care facilities from 2001 to 2006." Journal of clinical microbiology 46(2): 707-712.
Guastalli, E., et al. (2013). "Virulence characteristics of Escherichia coli isolates obtained from commercial one-week-old layer chicks with diarrhea." African Journal of Microbiology Research 7(47): 5306-5313.
Guillard, T., et al. (2014). "aac(6′)-Ib-cr is the major plasmid-mediated quinolone resistance determinant in extended-spectrum β-lactamase-producing Escherichia coli in eastern France." Journal of global antimicrobial resistance 2(2): 111-113.
Hawkey, P. M. and A. M. Jones (2009). "The changing epidemiology of resistance." Journal of Antimicrobial Chemotherapy 64(suppl_1): i3-i10.
Hazeleger, W. C., et al. (2016). "Quantification of growth of Campylobacter and extended spectrum β-lactamase producing bacteria sheds light on black box of enrichment procedures." Frontiers in microbiology 7: 1430.
Huang, S.-Y., et al. (2009). "Increased prevalence of plasmid-mediated quinolone resistance determinants in chicken Escherichia coli isolates from 2001 to 2007." Foodborne pathogens and disease 6(10): 1203-1209.
Institute, C. a. L. S. (2017). "Performance standards for antimicrobial susceptibility testing : M100–S27." Clinical and Laboratory Standards Institute, Wayne, PA.
Iovleva, A. and R. A. Bonomo (2017). "The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world." Journal of travel medicine 24(suppl_1): S44-S51.
Jakobsen, L., et al. (2010). "Broiler chickens, broiler chicken meat, pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients." International journal of food microbiology 142(1-2): 264-272.
Jiang, Y., et al. (2008). "Plasmid-mediated quinolone resistance determinants qnr and aac(6′)-Ib-cr in extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae in China." Journal of Antimicrobial Chemotherapy 61(5): 1003-1006.
Johnson, T. J., et al. (2012). "Associations between multidrug resistance, plasmid content, and virulence potential among extraintestinal pathogenic and commensal Escherichia coli from humans and poultry." Foodborne pathogens and disease 9(1): 37-46.
Kim, E. S., et al. (2009). "Prevalence of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme among Enterobacteriaceae blood isolates in Korea." Antimicrobial agents and chemotherapy 53(6): 2643-2645.
Kim, H. B., et al. (2009). "oqxAB encoding a multidrug efflux pump in human clinical isolates of Enterobacteriaceae." Antimicrobial agents and chemotherapy 53(8): 3582-3584.
Kim, J., et al. (2005). "CTX-M and SHV-12 β-lactamases are the most common extended-spectrum enzymes in clinical isolates of Escherichia coli and Klebsiella pneumoniae collected from 3 university hospitals within Korea." FEMS Microbiology letters 245(1): 93-98.
Kluytmans, J. A., et al. (2012). "Extended-spectrum β-lactamase–producing Escherichia coli from retail chicken meat and humans: comparison of strains, plasmids, resistance genes, and virulence factors." Clinical Infectious Diseases 56(4): 478-487.
Lee, J., et al. (2016). "Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea." Letters in applied microbiology 62(1): 84-90.
β-lactamase and plasmid-mediated fluoroquinolone resistance genes in Escherichia coli isolated from chickens in Anhui province, China." PloS one 9(8): e104356.
Lim, K.-T., et al. (2009). "Characterization of multidrug resistant ESBL-producing Escherichia coli isolates from hospitals in Malaysia." BioMed Research International 2009.
Liu, Y.-Y., et al. (2016). "Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study." The Lancet infectious diseases 16(2): 161-168.
Luangtongkum, T., et al. (2009). "Antibiotic resistance in Campylobacter: emergence, transmission and persistence." Future Microbiol 4(2): 189-200.
Ma, J., et al. (2009). "High prevalence of plasmid-mediated quinolone resistance determinants qnr, aac(6′)-Ib-cr, and qepA among ceftiofur-resistant Enterobacteriaceae isolates from companion and food-producing animals." Antimicrobial agents and chemotherapy 53(2): 519-524.
Madec, J.-Y. and M. Haenni (2018). "Antimicrobial resistance plasmid reservoir in food and food-producing animals." Plasmid.
Madico, G., et al. (1995). "Arbitrarily primed PCR DNA fingerprinting of Escherichia coli O157: H7 strains by using templates from boiled cultures." Journal of clinical microbiology 33(6): 1534-1536.
Mammeri, H., et al. (2005). "Emergence of plasmid-mediated quinolone resistance in Escherichia coli in Europe." Antimicrobial agents and chemotherapy 49(1): 71-76.
Manges, A. R., et al. (2007). "Retail meat consumption and the acquisition of antimicrobial resistant Escherichia coli causing urinary tract infections: a case–control study." Foodborne pathogens and disease 4(4): 419-431.
Nahar, A., et al. (2018). "Prevalence and characteristics of extended-spectrum β-lactamase-producing Escherichia coli in domestic and imported chicken meats in Japan." Journal of Veterinary Medical Science 80(3): 510-517.
Nordstrom, L., et al. (2013). "Foodborne urinary tract infections: a new paradigm for antimicrobial-resistant foodborne illness." Frontiers in
microbiology 4: 29.
Olsen, R. H., et al. (2014). "Extended-spectrum β-lactamase-producing Escherichia coli isolated from poultry: a review of current problems, illustrated with some laboratory findings." Avian Pathology 43(3): 199-208.
Organization, W. H. (2017). "Critically important antimicrobials for human medicine: ranking of antimicrobial agents for risk management of antimicrobial resistance due to non-human use."
Overdevest, I., et al. (2011). "Extended-spectrum β-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands." Emerging infectious diseases 17(7): 1216.
Park, K. S., et al. (2012). "Prevalence of the plasmid-mediated quinolone resistance genes, aac(6′)-Ib-cr, qepA, and oqxAB in clinical isolates of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli and Klebsiella pneumoniae in Korea." Annals of Clinical & Laboratory Science 42(2): 191-197.
Paterson, D. (2000). "Recommendation for treatment of severe infections caused by Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs)." Clinical Microbiology and Infection 6(9): 460-463.
Paton, A. W. and J. C. Paton (1998). "Detection and Characterization of Shiga Toxigenic Escherichia coli by Using Multiplex PCR Assays forstx 1, stx 2, eaeA, Enterohemorrhagic E. coli hlyA, rfb O111, and rfb O157." Journal of clinical microbiology 36(2): 598-602.
Picard, B., et al. (1999). "The link between phylogeny and virulence in Escherichia coli extraintestinal infection " Infection and immunity 67(2): 546-553.
Pitout, J. D., et al. (2005). "Emergence of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs) in the community." Journal of Antimicrobial Chemotherapy 56(1): 52-59.
Poulou, A., et al. (2014). "Modified CLSI extended-spectrum β-lactamase (ESBL) confirmatory test for phenotypic detection of ESBLs among
stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (ESBL)-producing and carbapenem-resistant Escherichia coli." International journal of food microbiology 241: 283-290.
Rawat, D. and D. Nair (2010). "Extended-spectrum β-lactamases in Gram Negative Bacteria." Journal of global infectious diseases 2(3): 263.
Robicsek, A., et al. (2006). "The worldwide emergence of plasmid-mediated quinolone resistance." The Lancet infectious diseases 6(10): 629-640.
Robicsek, A., et al. (2006). "Fluoroquinolone-modifying enzyme: a new adaptation of a common aminoglycoside acetyltransferase." Nature medicine 12(1): 83.
Russo, T. A. and J. R. Johnson (2000). "Proposal for a new inclusive designation for extraintestinal pathogenic isolates of Escherichia coli: ExPEC." The Journal of infectious diseases 181(5): 1753-1754.
Smith, J. L., et al. (2007). "Extraintestinal pathogenic Escherichia coli." Foodborne pathogens and disease 4(2): 134-163.
Strahilevitz, J., et al. (2009). "Plasmid-mediated quinolone resistance: a multifaceted threat." Clinical microbiology reviews 22(4): 664-689.
Unno, T., et al. (2009). "Absence of Escherichia coli phylogenetic group B2 strains in humans and domesticated animals from Jeonnam Province, Republic of Korea." Applied and environmental microbiology 75(17): 5659-5666.
Xia, L.-N., et al. (2011). "A survey of β-lactamase and 16S rRNA methylase genes among fluoroquinolone-resistant Escherichia coli isolates and their horizontal transmission in Shandong, China." Foodborne pathogens and disease 8(12): 1241-1248.
Xu, X., et al. (2014). "Prevalence and characterization of cefotaxime and ciprofloxacin co-resistant Escherichia coli isolates in retail chicken carcasses and ground pork, China." Microbial Drug Resistance 20(1): 73-81.
Yamane, K., et al. (2008). "Plasmid-mediated qepA gene among Escherichia coli clinical isolates from Japan." Antimicrobial agents and chemotherapy 52(4): 1564-1566.
pump, QepA, found in an Escherichia coli clinical isolate." Antimicrobial agents and chemotherapy 51(9): 3354-3360.
Yuan, L., et al. (2009). "Molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli isolates from chickens in Henan Province, China." Journal of medical microbiology 58(11): 1449-1453.
Yue, L., et al. (2008). "Prevalence of plasmid-mediated quinolone resistance qnr genes in poultry and swine clinical isolates of Escherichia coli." Veterinary microbiology 132(3-4): 414-420.
Zheng, H., et al. (2012). "Prevalence and characterisation of CTX-M β-lactamases amongst Escherichia coli isolates from healthy food animals in China." International journal of antimicrobial agents 39(4): 305-310.
국문 초록
광범위 베타 락탐계 분해 효소 (ESBL)를 생산하는 대장균은 심각 한 공중 보건 문제로 대두되고 있다. 플라스미드 매개로 전달되는 퀴놀론 저항성 (PMQR) 유전자는 식용 동물 또는 식품으로부터 분리 된 균주에 널리 퍼져 있으며 그 중 일부는 광범위 베타 락탐 계 분해 효소 (ESBLs) 생성 균주이다. 본 연구에서는 한국의 유 통 중인 소매 닭에서 분리 된 ESBL 생성 대장균 67주의 항생제 저항성, phylogenetic group 및 병원성 유전자 분포를 조사 하였다. 또한, 총 67주의 ESBL 생성 대장균의 plasmid-mediated quinolone resistance (PMQR) 유전자 분포를 조사 하였다. 모든 균주는 blaCTX-M을 보유하고 있었고, blaCTX-M-type 중 blaCTX-M-65(52.2 %, 35/67)와 blaCTX-M-55 (25.4 %, 17/67)가 우세하였으며,
3주는 blaCTX-65와 blaCTX-M-55 모두 보유하고 있었다. 또한, ESBL
생성 대장균의 절반 이상이 blaTEM을 보유하고 있었다. 놀랍게도,
ESBL 생성 균주 중 35 (52.2 %, 35/67) 주가 PMQR 결정 인자 를 함께 보유하고 있었다. 가장 많이 보유한 PMQR 유전자는 qnrS (91.4 %, 32/35)이었고, 그 중 6주는 qnrS에 더하여 aac(6')-Ib-cr도 가지고 있었다. oqxAB 및 aac(6')-Ib-cr는 각
각 2주 (5.7 %), 1주 (2.9 %)에서 검출되었다. qnrA, qnrB 및 qepA를 보유한 균주는 없었다. PMQR 보유 균주 중 가장 흔한 ESBL type은 CTX-M-65 (91.4 %, 32/35)이었다. 항생제 감수 성 검사 (MIC test) 결과 ESBL 생성 균주 중 98.5%가 3가지 이 상의 서로 다른 그룹에 속한 항생제에 내성을 가진 다제내성 균주 였고 PMQR gene을 함께 보유한 35주는 모두 다제내성 (MDR) 균주 임이 밝혀졌다. 그 중 qnrS와 aac(6')-Ib-cr을 동시에 보유 하고 있는 균주는 ciprofloxacin과 kanamycin에 대한 내성에 시너 지 효과를 보였다. Phylogenetic group 분석 결과 group A (56.7%)에 속한 균주가 가장 많았고 group B1 (19.4%), group E (8.9%), group B2 (6%), group D (6%) 순으로 분포해 있었다. 또한, 장외전염 병원성 대장균 (ExPEC)과 관련된 병원성 유전자 는 group B1, B2, D, 그리고 E에 속한 균주에 높게 분포해 있음을 밝혀내었다. 본 연구에서의 결과는 한국의 유통중인 닭에 높은 비 율로 ESBL 생성하는 다제내성 대장균이 분포해 있으며, 닭이 ExPEC를 사람에게 전이시키는 reservoir가 될 수 있음을 밝혀내
주요어: 항생제 내성, 광범위 베타 락탐 분해 효소 (ESBL), 플라스미드 매개 퀴놀론 저항성 유전자, 대장균, 닭