315
흰쥐의 배양된 간세포에서 ethanol에 의해 유도된 p42/44 MAPkinase가 IGF system에 미치는 효과
이선미·김종훈·강창원*
전북대학교 수의과대학, 생체안전성 연구소 (게재승인: 2006년 12월 3일)
Effects of ethanol-induced p42/44 MAPkinase activity on IGF system in primary cultured rat hepatocytes
Sun-Mi Lee, Jong-Hoon Kim, Chang-Won Kang*
College of Veterinary Medicine, Bio-safety Research Institute, Chonbuk National University, Jeonju 561-756, Korea
(Accepted: December 3, 2006)
Abstract : Ethanol abuse is associated with liver injury, neurotoxicity, modulation of immune responses, and increased risk for cancer, whereas moderate ethanol consumption exerts protective effects against liver injury.
However, the underlying signal transduction mechanisms of insulin-like growth factors (IGFs) which play an important regulatory role in various metabolism mechanisms are not well understood. We investigated the effects of ethanol-induced p42/44 activity on IGF-I secretion, IGF-I receptor and IGFBP-1 secretion using radioimmunoassay and western blotting in primary cultured rat hepatocytes. The p42/44 activity, IGF-I secretion and IGF-I receptor activity significantly accelerated compared to control at 10 and 30 min after 200 mM ethanol treatment, but then it became suppressed at 180 min. In contrast, IGFBP-1 secretion was inhibited compared to control at 30 min after 200 mM ethanol treatment, but increased at 180 min. The IGF- I secretion, IGF-I receptor and p42/44 activity at 30 min after 200 mM ethanol treatment accelerated with increasing ethanol concentration but IGFBP-1 secretion inhibited (
p< 0.05). The increased IGF-I secretion, inhibited IGFBP-1 secretion and IGF-IR activity by ethanol-induced temporal p42/44 activity at 30 min after ethanol treatment was blocked by treatment with PD98059. Alcohol dehydrogenase (ADH) inhibitor, 4- methylpyramazole blocked the changes of IGF-I secretion, IGFBP-1 secretion, and IGF-IR activity by ethanol- induced p42/44 activity at 30 and 180 min. Taken together, these results suggest that ethanol is involved in the modulation of IGF-I and IGFBP-1 secretion and IGF-IR activity by p42/44 activity in primary cultured rat hepatocytes. In addition, changing of p42/44 activity by ethanol was caused with ADH.
Key words : hepatocyte, IGF-I, IGF-I receptor, IGFBP-1, p42/44
서 론
Ethanol
은간장에서 단백질, glucose, triglycerides
및phospholipid
등의변화에따라ethanol
성간염,
섬유증및지방간을유발한다고알려져있다
[10, 20, 21, 26].
또한
ethanol
은간세포에서세포증식과사멸을유발하며
,
여기에관여하는인자로는reactive oxygen species
(ROS), protein kinase C, adenylate cyclase
및mitogen- activated protein kinase(MAPkinase)
가 있다.
특히MAPkinase
중p42/44 (extracellular signal regulated kinase 1
과2)
는ethanol
에대한간세포활성에깊이관여하고있다고보고되었다
[1, 2, 8, 14, 27].
흰쥐의배양된간세포에서
angiotensin-II
와혈청에의하여활성화된
p42/44
는ethanol
에의하여억제되며,
간*Corresponding author: Chang-Won Kang
College of Veterinary Medicine, Chonbuk National University, Jeonju 561-756, Korea [Tel: +82-63-270-3715, Fax: +82-63-270-3780, E-mail: [email protected]]
세포에서
ethanol
은p42/44 MAPkinase
와p38
등을장시간자극한다고보고되었다
[4].
이와같이일차배양된간세포에서
ethanol
에의한p42/44
활성과이에따른세 포사멸과의연관성은이미잘알려진사실이다.
Insulin-like growth factor-I(IGF-I)
은기능적으로생체의내분비
,
방계분비및자기분비를통하여세포증식,
분화및대사에관여하는호르몬이며
,
순환혈액내존재하는대부분
IGF-I
은주로간세포에서생산되는것으로보고되어있다
[7, 9]. IGF-I
은DNA
합성에필요한G1
기에서
S
기로 진입하는데 필요한 인자이며, IGF-I
receptor
에반응함으로써세포의유사분열효과와apop-
tosis
를억제하는역할을 한다[3, 30, 31].
또한순환혈 액내에존재하는IGF-I
은6 types
의IGF-binding proteins (IGFBPs)
과친화력을가지고있다[5, 11].
이중IGFBP- 1
은주로간세포에서생산되며,
그기능이아직까지명 확하지않지만IGF-I
반감기조절,
세포증식억제,
면역억제
,
세포리모델링억제및세포사멸뿐만아니라 세포cytokine
등의분비조절에관여하고있으며,
특히 당뇨및생체염증작용등에의한생체이화학적반응 에관여함이보고되었다[11, 15].
그러나ethanol
에대 한호르몬분비기전이아주복잡하여아직까지명확하 게밝혀지지않았으며,
특히IGFs system
과의연관성은더우기명확하게알려져있지않은상태이다
.
따라서이 연구는배양한흰쥐의간세포에서ethanol
에의해유도 된p42/44 MAPkinase
가IGF-I
와IGFBP-1
분비및IGF- IR
에미치는영향에대하여연구하였다.
재료 및 방법
실험동물
실험동물은체중
150 g
내외의Sprague-Dawley(SD)
계수컷랫트를사용하였다
.
간세포에 ethanol 처리
간세포분리는
Seglen
등[29]
의방법에따라sodium pentobarbital(45 mg/kg BW)
을복강에주입하여마취시킨후
collagenase(Sigma, USA)
를관류하였다.
요약하 면, Ca
2+-free
완충액(8.3 g NaCl, 0.5 g KCl, 2.4 g Hepes,
pH 7.4)
을이용하여간장에충만되어있는혈액을제거하였다
. Collagenase
완충액을이용하여간세포를분리한후
Williams E buffer(Sigma, USA)
로세척하였다. Trypan blue
용액을이용하여세포의생존율(97%
이상)
을확인한후
3
×10
6 세포를collagen
이coating
된petri- dish(60 mm diameter)
에fetal bovine serum(5%, v/v;
Hyclone, USA)
이함유된Williams E
액체배지에넣은다음
37
oC
에2
시간세포를안정화하여간세포를부착시켰으며
,
이후serum
이첨가되지않은부착된간세포에200 mM ethanol
을0.5, 10, 30, 60
및180
분동안처리하였다
.
IGF-I 분석
IGF-I
을분석은Park
등[24]
의방법에따랐다.
요약하 면, ethanol
을처리한간세포배양상층액200
µl
에acid- ethanol(2 M HCl : ethanol = 1 : 7) 800
µl
를 첨가한 후3,000 rpm
으로30
분간냉장원심분리하여IGF-I
을분리 하였다.
여기에100
µl
에polyclonal anti-IGFs(Gro-Pep, Australia) 50
µl
를 넣은 후조제된[
125I]-IGF-I (20,000 cpm/100
µl)
을100
µl
첨가하고60
분후에혼합하여각각 의시험관에4
oC 18
시간반응시켰다.
그후말혈청(Sigma, USA) 50
µl
와12% polyethylene glycol #8000 (PEG) 1 ml(Sigma, USA)
을첨가하여3,000 rpm
에서30
분간원심분리시켜결합형과비결합형을분리시켰고 결합형의방사능을
gamma counter(Packard, USA)
로측 정하였다.
Western immunoblotting 분석
BCA
단백질정량법을이용하여각각40
µg
의sample
들을
10% SDS-PAGE
전기영동을시킨후, polyvinylidine difluoride(PVDF)
에이동하였다. PVDF
은5%
탈지유에1
시간동안차단시키고,
일차항체를1 : 250~1 : 1,000
비 율로희석하여4
oC
에서18
시간이상반응시켰다.
그후, PVDF
을0.1% Tween-20/1X TBS
에10
분간격으로3
회 세척하고, 1%
탈지유에3,000
배 희석된horseradish- peroxidase labeled secondary antibody
에1
시간동안반응시킨후
, 3
회세척하고ECL solution(Santa Cruz, USA)
처리한다음
X-ray
필름에노출시켜현상하였다.
통계처리
이실험에서측정된결과는
mean
±SD
로나타내었고군간의차이는
Student's
t-test
를이용하였으며,
p-value
< 0.05
경우를유의한차이로인정하였다.
결 과
Ethanol 처리시간이 p42/44, IGF-I분비, IGF-IR 및 IGFBP-1 분비에 미치는 효과
배양된간세포에
200 mM ethanol
을0.5, 10, 30, 60
및180
분동안처리한후ethanol
이p42/44, IGF-I
분비, IGF- IR
및IGFBP-1
분비에미치는 효과는Fig. 1A
에서와같 다.
즉인산화된p42/44
활성은10
과30
분에서대조군에 비하여유의성있게증가(
p< 0.05)
되었으나, 180
분에서는유의성있게감소
(
p< 0.01)
되었다.
IGF-I
분비는10
분및30
분처리군에서대조군에비하여증가
(
p< 0.05)
하였으나, 180
분에서대조군에비하여유의성있게감소
(
p< 0.01)
되었다(Fig. 1B).
IGF-IR
활성역시ethanol 10
분과30
분처리군이대조 군에비하여유의성있게증가(
p< 0.01)
되었으나, 180
분에서는대조군에비하여감소
(
p< 0.05)
되었다(Fig. 1C).
IGFBP-1
분비는ethanol 30
분처리군에서대조군에비 하여유의성있게감소(
p< 0.05)
되었으나, ethanol 180
분처리군에서는
IGFBP-1
분비가대조군에비하여유의성 있게증가(
p< 0.05)
되었다(Fig. 1D).
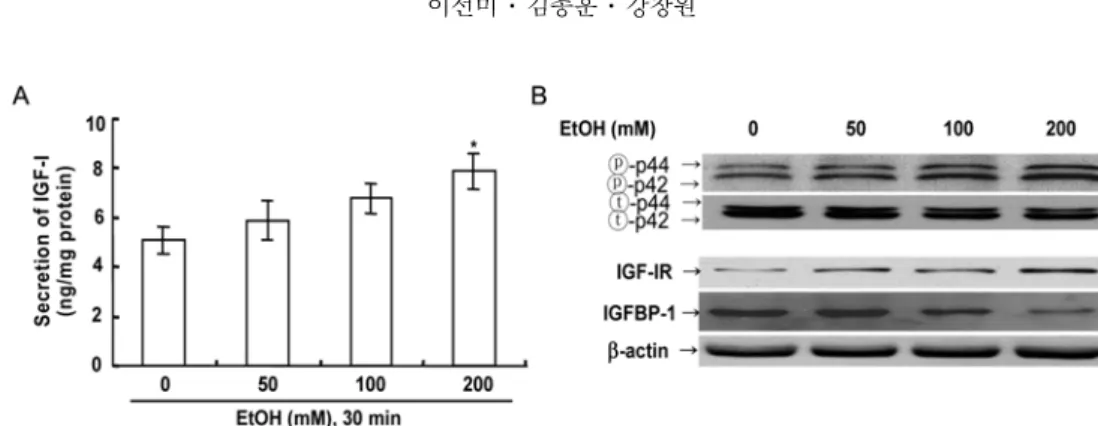
Ethanol 처리농도가 p42/44, IGF-I, IGF-IR 및 IGFBP-1 분비에 미치는 효과
Ethanol 10
분처리에의한IGF-I
분비와p42/44
활성은유의성있게증가하였지만
IGFBP-1
분비에있어서는ethanol 10
분처리가대조군에비하여차이는인정되지않았다
.
따라서저자들은ethanol 30
분처리에의한0, 50, 100
및200 mM ethanol
농도에서확인하였다.
그결 과인산화된p42/44
활성은100
과200 mM ethanol
처 리군이대조군에비하여증가됨을확인하였다(Fig. 2B).
이때
IGF-I
분비는ethanol
농도의존적으로증가되었으며
,
특히200 mM
에서대조군에비하여유의성있게증가
(
p< 0.05)
하였다(Fig. 2A).
그러나예비실험에서500 mM ethanol 30
분처리시간세포가거의사멸하였다(data not shown).
IGF-IR
활성은ethanol 30
분처리에서농도의존적으로증가되었으며
,
특히200 mM ethanol
처리군에서대 조군에비하여유의성 있게증가되었다(Fig. 2B).
이에반하여
IGFBP-1
분비는ethanol
농도의존적으로감소되었으며
,
특히100
과200 mM ethanol
처리군이대조군 에비하여감소되었다(Fig. 2B).
Ethanol에 의한 IGF-I, IGF-IR 및 IGFBP-1분비의 일시적인 변동과 p42/44 활성과의 관련성
배양된간세포에서
ethanol
에의한 일시적인p42/44
활성과이에따른
IGF-I, IGF-IR
활성및IGFBP-1
분비의상호관련성여부를구명하기위하여
p42/44
억제제인
PD98059(10 µM)
를 사용하였다.
그 결과200 mM ethanol 30
분처리군은대조군에비하여p42/44
활성증 가. IGF-1
분비증가, IGF-IR
활성증가및IGFBP-1
분비를감소시킨반면
, p42/44
억제제인PD98059
단독처 리군은대조군에비하여p42/44
활성감소, IGF-I
분비감 소, IGF-IR
활성감소및IGFBP-1
분비증가를유발함으로써
ethanol 180
분처리에따른p42/44, IGF-I
분비, IGF-
IR
활성및IGFBP-1
분비양상과같은결과를관찰하였다
.
또한ethanol 30
분처리후PD98059
처리는ethanol
에의한일시적인인산화
p42/44
증가, IGF-I
분비증가, Fig. 1 . Time course of 200 mM ethanol-induced p42/44
activity (A), IGF-I secretion (B), IGF-IR activity (C) and IGFBP-1 secretion (D) in primary cultured rat hepatocytes.
IGF-I secretion was determined by radioimmunoassay (RIA). IGFBP-1, IGF-IR and p42/44 activity as assessed using a densitometer and Western blotting in primary cultured rat hepatocytes (
n= 7). Equal amounts (40
µg) of total cell lysates were analyzed by SDS-PAGE and immu- noblotted. Data are mean
±SD values. *,
p< 0.05; **,
p< 0.01 compared to control.
Fig. 2. Effects of different concentrations of ethanol on secretion of IGF-I secretion (A), p42/44 activity, IGF-IR activity and IGFBP-1 secretion (B) in primary cultured rat hepatocytes. IGF-I secretion was determined by RIA .IGFBP-1, IGF- IR and p42/44 activity as assessed using a densitometer and Western blotting in primary cultured rat hepatocytes (
n= 7).
Equal amounts (40
µg) of total cell lysates were analyzed by SDS-PAGE and immunoblotted. Data are mean
±SD values.
*,
p< 0.05 compared to control.
Fig. 3. Effects of PD98059 on changing of IGF-I secretion (A), p42/44 activity, IGF-IR activity, and IGFBP-1 secretion (B) at 30 min. after ethanol treatment in primary cultured rat hepatocytes. IGF-I secretion was determined by RIA. IGFBP- 1, IGF-IR and p42/44 activity as assessed using a densitometer and Western blotting in primary cultured rat hepatocytes (
n= 7). Equal amounts (40
µg) of total cell lysates were analyzed by SDS-PAGE and immunoblotted. Data are mean
±SD values. *,
p< 0.05 compared to control.
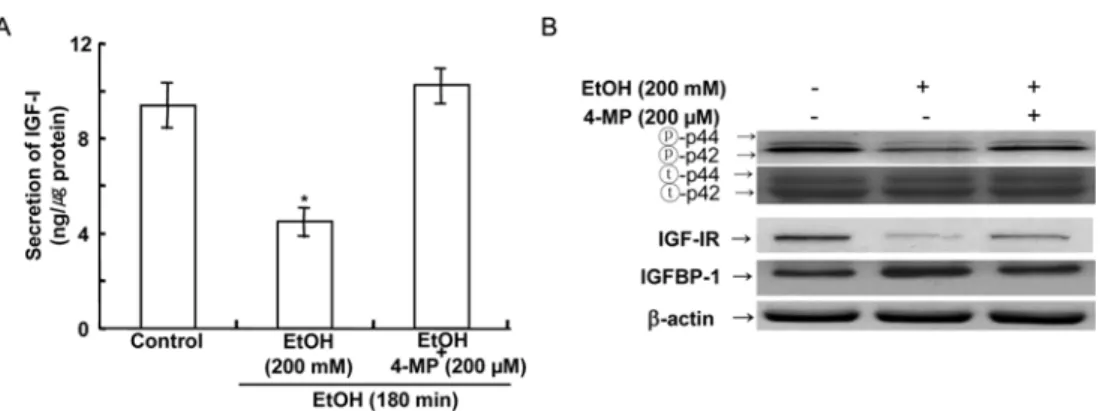
Fig. 4. Effects of 4-MP on the change of IGF-I secretion (A), p42/44 activity, IGF-IR activity, IGFBP-1 secretion (B) at 30 min. after ethanol treatment in primary cultured rat hepatocytes. IGF-I secretion was determined by RIA. IGFBP- 1, IGF-IR and p42/44 activity as assessed using a densitometer and Western blotting in primary cultured rat hepatocytes (
n= 7). Equal amounts (40
µg) of total cell lysates were analyzed by SDS-PAGE and immunoblotted. Data are mean
±SD values. *,
p< 0.05 compared to control.
IGF-IR
활성증가및IGFBP-1
분비감소를억제하였다(Fig. 3A, B).
Ethanol 의한 IGF-I, IGFBP-1, IGF-IR 및 p42/44 활성과 ADH와의 관련성
배양된간세포에서
ethanol
에의한IGF-I
분비, IGFBP- 1
분비, IGF-IR
활성및p42/44
활성의변동이ethanol
분해효소인
alcohol dehydrogenase(ADH)
와의관련성여부를확인하기위하여
ethanol
분해효소억제제인200
µM 4-methylprazol(4-MP)
을적용하였다.
그결과 일차 배양된간세포에200 mM ethanol
을30
분처리에의한일시적인
IGF-I
분비증가, IGFBP-1
분비감소, IGF-IR
활성증가및
p42/44
활성증가는4-MP
에의하여그작 용이 차단됨이확인되었다(Fig. 4A, B).
또한200 mM ethanol 180
분처리에 의한IGF-I
분비 감소, IGFBP-1
분비증가
, IGF-IR
활성감소및p42/44
활성감소역시4-MP
에의하여차단되었다(Fig. 5A, B).
고 찰
이연구에서는배양된간세포에서
ethanol
이세포신호 전달계인p42/44
활성, IGF-I
분비, IGF-IR
활성, IGFBP-
1
분비와의관련성및ethanol
대사와의연관성을검토하였다
.
간세포에서
ethanol
은MAPkinase
를조절하며MAP kinase
중p42/44
는세포분화,
증식및apoptosis
와밀접한관련성이있다고알려져있다
[1].
특히,
간세포에서ethanol
은p42/44
활성을길게유지시키며,
또다른연구 에서는간세포에ethanol
처리는일시적으로p42/44
활성을증가시키지만만성
ethanol
투여는그활성이감소한다고보고하였다
[17, 21].
또한간세포에ethanol 10-
30
분처리에의한p42/44
활성의일시적증가는간세포의스트레스저항력증가를유발하며
, ethanol
처리에의한장시간
p42/44
활성유지는간세포증식주기억제에관여하고있음을보고하였다
[4, 28, 33].
이연구에서는 배양된간세포에ethanol
처리는일시적으로p42/44
활성이증가되었고
180
분에서는감소되었다.
이는장시간ethanol
처리에 의하여 간세포 분화와 증식억제 및apoptosis
에의한것으로사료된다.
또한장시간ethanol
노출시간에따른다른연구와이연구의
p42/44
활성차이는초기배양된흰쥐의간세포상태
,
분리된간세포를배양접시에부착시키는시간및배지의유지조건이 다름으로써그결과가다를수있음을암시하였다
.
또한ethanol 30
분처리에의한일시적인p42/44
활성은농도의존적으로증가하였다
.
이와같이ethanol
에의한일시적인
p42/44
활성증가양상은상기연구결과와일치하였으며
,
이러한활성증가는일시적인세포스트레스저항력이관여할수있음을암시하였다
.
간세포에
IGF-I
첨가는p42/44
활성을증가시키며,
간 세포에서ethanol
은IGF-I
분비를억제한다고보고하였다
[12, 13].
또한전보[15, 24]
에서흰쥐에ethanol
투여는혈액과간조직의
IGF-I
분비를억제시켰으며,
일차배양된간세포에서
ethanol
은산화성스트레스증가에의하여
IGF-I
분비가억제된다고보고하였다.
간세포의스트레스저항력증가는
IGF-I
분비를증가시키며, ethanol
은일시적인세포스트레스저항성에영향이있음을보고 하였다
[22, 32]. IGF-IR
은세포증식에관여하는IGF-I
과결합하여자동인산화를유도하고
ras-raf-MAPkinase cascade
를통하여세포의유사분열,
형질변화및apoptosis
를억제시킴으로써
IGF-I
분비를자극한다고보고하였Fig. 5. Effects of 4-MP on the IGF-I secretion (A), p42/44 activity, IGF-IR activity and IGFBP-1 secretion (B) at 180 min. after ethanol treatment in primary cultured rat hepatocytes. IGF-I secretion was determined by RIA. IGFBP-1, IGF- IR and p42/44 activity as assessed using a densitometer and Western blotting in primary cultured rat hepatocytes (
n= 7).
Equal amounts (40
µg) of total cell lysates were analyzed by SDS-PAGE and immunoblotted. Data are mean
±SD values.
*,
p< 0.05 compared to control.
다
[19].
이연구에서간세포에ethanol
처리는일시적으로
IGF-I
분비증가와IGF-IR
활성이 증가되고ethanol 180
분처리에서는IGF-I
분비억제와IGF-IR
활성억제 가관찰되었다.
이는배양된간세포에서ethanol
에의한일시적인
IGF-I
분비와IGF-IR
활성증가는세포의스트레스저항력증가와관련이있으며
,
장시간ethanol
노 출에대한IGF-I
분비억제와IGF-IR
활성억제는이전보
[16, 24]
에서와같이ethanol
에의한산화성스트레스증가에의한세포분화와증식억제가
IGF-I
분비를억제시킬수있음을암시하였다
.
또한일시적인ethanol
처리에의한
IGF-I
분비와IGF-IR
활성변동은p42/44
억 제제에의하여차단되었으며,
장시간ethanol
처리에의 한IGF-I
분비와IGF-IR
활성변동양상이p42/44
억제제단독처리군과일치함에따라단·장시간
ethanol
에 의한IGF-I
분비와IGF-IR
활성변동은p42/44
활성이 관련될수있는가능성을암시하였다.
배양된간세포에생산되는
IGFBP-1
은Hs578T breast cancer cell
과간세포에서apoptosis
를증가시킨다고보고[11, 14, 25]
하였고이연구자의이전연구역시ethanol
에의한간세포산화성스트레스는
IGFBP-1
분비를증 가시킬수있음을보고하였다[16].
이연구에서는배양 된간세포에ethanol 30
분처리는IGFBP-1
분비를감소시켰으며
, ethanol 180
분처리는그분비를증가시켰다. p42/44
억제제단독투여는ethanol 180
분처리에의한IGFBP-1
분비양상과비슷하였으며, ethanol 30
분처리에의한
IGFBP-1
분비변동역시p42/44
억제제에의하 여그분비가증가되었다.
이는간세포에서일시적혹은장시간
ethanol
에 의하여 유도된산화성 스트레스가IGFBP-1
분비에영향을미쳤으며,
이분비변동 역시p42/44
활성이관련될수있는가능성을암시하였다.
이어이연구자는일시적혹은장기간
ethanol
에유도된
p42/44
활성, IGF-I
분비, IGF-IR
활성및IGFBP-1
분비변동이
ethanol
대사와어떠한연관성이있는지를구명하기위하여단시간혹은장시간
ethanol
을처리한일차배양된간세포에
ADH
억제제를처리한결과그분비와활성이차단됨을확인하였다
. Ethanol
에의한간독성과
apoptosis
는ethanol
대사산물인ADH
활성에의한acetaldehyde
가그원인이라고보고되었다[6, 23, 32].
또 한간세포에서ethanol
대사산물인acetaldehyde
가일시적인
p42/44
활성을증가시키며,
장시간ethanol
처리에 따른간세포내acetaldehyde
축적증가가apoptosis
를유 발한다고보고되었다[17, 18].
이는ethanol
대사산물인ADH
가p42/44
활성, IGF-I, IGFBP-1
및IGF-IR
활성에 영향을미칠수있음을암시하였다.
그러나ethanol
대 사산물인acetaldehyde
가IGF-I, IGFBP-1
및IGF-IR
활성과직접적인관련성은앞으로깊이있게연구를진행
시켜야할부분으로사료된다
.
따라서간세포에서
ethanol
은IGF-I, IGF-IR
활성및IGFBP-1
분비에영향을미치며,
그호르몬과활성변동은
p42/44
활성과관련성이있음을관찰할수있었다.
더욱이
ethanol
에의한p42/44
활성은간세포에서ethanol
산화과정에서생기는
ADH
생성과상관관계가있는것 으로판단되었다.
결 론
Ethanol
은 간에서 단백질, glucose, triglycerid-es
및phospholipid
등의변동에의하여ethanol
성간염,
섬유증 및지방간을유발한다고알려져있다.
그러나ethanol
의호르몬분비기전은아주복잡하기때문에아직까지명 확하게밝혀지지않은상태이다
.
따라서이연구는간세 포에ethanol
이세포신호전달계인p42/44
활성에미치는효과를확인하고그활성이
IGF-I, IGFBP-1
분비및IGF-IR
활성에어떠한관련성이있는지를연구하였다.
이연구는일차배양된간세포에
ethanol
을처리하고이를방사면역측정법과
western blotting
이용하여아래와같 은결론을얻었다.
흰쥐의배양된 간세포에서
p42/44
활성, IGF-I
분비및
IGF-IR
활성은200 mM ethanol
처리후10-30
분에 서 대조군에 비하여 증가하고180
분에 감소하였다(
p< 0.05).
이에반하여IGFBP-1
분비는30
분에일시적으로대조군에비하여감소되었으며
180
분에증가하였 다(
p< 0.05).
흰쥐의배양된간세포에서
p42/44
활성, IGF-I
분비및
IGF-IR
활성은ethanol
농도의존적으로증가하였으 며,
특히200 mM ethanol
에서대조군에비하여증가하고
, IGFBP-1
분비는유의성있게감소하였다(
p< 0.05).
200 mM ethanol 30
분처리에 의한 일시적인IGF-I
와IGFBP-1
분비및IGF-IR
활성변동은PD98059
에의하여차단되었다
.
또한200 mM ethanol 30
혹은180
분처리에의한
p42/44
활성변동은ADH
억제제인4-MP
에 의하여 차단되었으며,
이때의ethanol
에 의한IGF-I, IGFBP-1
분비및IGF-IR
활성역시차단되었다.
이상의결과를종합하면흰쥐의배양된간세포에서
ethanol
에의한p42/44
활성은IGF-I, IGFBP-1
분비및IGF-IR
활성에영향을미쳤으며,
이러한변동은ethanol
대사산물인
ADH
가관여하고있음을알수있었다.
감사의 글
이논문은
2005
년도전북대학교지원연구비에의하여연구되었으며또한