대구광역시 앞산공원 산림식생의 조성 및 구조 변화
오정학1⋅김준수2⋅김학윤3⋅조현제 2*
1국립산림과학원 도시숲연구센터, 2자연과숲연구소, 3계명대학교 지구환경학과
Changes in Composition and Structure of Forest Vegetation of Apsan Park, Daegu Metropolitan City
Jeong Hak Oh
1, Jun-Soo Kim
2, Hak Yun Kim
3and Hyun Je Cho
2*1Urban Forests Research Center, National Institute of Forest Science, Seoul 02455, Korea
2Nature and Forest Research Institute, Daegu 41476, Korea
3Department of Global Environment, Keimyung University, Daegu 42601, Korea
요 약: 대구광역시의 대표적 도시숲인 앞산공원을 대상으로 1997년과 2016년에 식물사회학적 방법으로 식생조사를 실시 하고 상관 우점종에 의한 정밀식생도를 작성하여 산림식생의 조성, 구조 및 공간적 분포 변화 경향을 분석하였다. 분석결 과, 식생유형의 개수는 1997년과 2016년 모두 22개로 동일하였으나 그 구성 종류에 있어서는 2개 유형이 증가하고 2개 유 형은 감소하였다. 산림식생 구성종의 단위면적당(/100㎡) 총피도는 1997년 약 163%에서 2016년 약 182%로 약 11%가 증 가하였으며, 자연식생이 인공식생에 비하여 두 배 이상 높은 경향이었다. 평균 출현종수는 1997년 약 25종에서 2016년 약 18종으로 7종이 감소하였다. 종다양도(H’)는 1997년 1.654에서 2016년 1.680으로 미미하게 증가하였으나, 종우점도(D)는 1997년 0.304에서 2016년 0.276으로 약 9%가 감소하였다. 전체 산림식생의 구성적 유사도는 약 78%로 종구성적 특성이 거의 동일한 것으로 나타났으며, 생활형 조성은 1997년 ‘G-R5-D4-e’에서 2016년 ‘MM-R5-D4-e’로 휴면형의 중심형이 과거 지중식물(G)에서 대형지상식물(MM)로 변화된 경향이었다. 산림식생의 공간 분포 면적은 인공식생이 자연식생에 비해 약 4배 이상 높은 감소율을 보였다. 산림경관 요소(패치)의 개수는 1997년 269개에서 2016년 294개로 약 9%가 증가한 반면, 그 평균 크기는 1997년 5.8 ha에서 2016년 5.1 ha로 약 12%가 감소된 경향이었다.1)
Abstract: The aim of our study was to identify the changing trends in the composition, structure, and spatial distribution of forest vegetation in Apsan Park, a representative urban forest in Daegu, South Korea. A vegetation survey was conducted in 1997 and 2016 using phytosociological methods, and a detailed vegetation map was created using the physiognomic dominant species. There were 22 vegetation types in both 1997 and 2016, but two of those types increased and two decreased.
The total coverage per unit area (100 ㎡) of the component vegetation species increased from 163% in 1997 to 182%
in 2016, and natural vegetation tended to be more than twice that of artificial vegetation. The average number of species decreased by seven from 25 in 1997 to 18 in 2016. Species diversity (Hʹ) increased only slightly from 1,654 in 1997 to 1,680 in 2016, while species dominance (D) decreased by 9% from 0.304 in 1997 to 0.276 in 2016. The similarity in the composition of the forest vegetation was about 78%, which was nearly the same. The life form spectrums of vascular plants changed from ‘G-R5-D4-e’ in 1997 to ‘MM-R5-D4-e’ in 2016 and the central dormancy type changed from geophytes (G) to megaphanerophytes (MM). The spatial distribution of the forest vegetation was reduced by approximately four times that of artificial vegetation. The number of forest landscape elements (patches) increased from 269 in 1997 to 294 in 2016, while the average area decreased by 12% from 5.8 ha in 1997 to 5.1 ha in 2016.
Key words: urban forest, vegetation change, landscape characteristics, spatial distribution
* Corresponding author E-mail: [email protected] ORCID
Hyun Je Cho https://orcid.org/0000-0003-0390-9386
JOURNAL OFKOREANSOCIETY OFFORESTSCIENCE ISSN 2586-6613(Print), ISSN 2586-6621(Online) http://e-journal.kfs21.or.kr https://doi.org/10.14578/jkfs.2019.108.2.177
177
서 론
도시화로 인한 토지이용의 변화 및 식생경관의 파편화 현상 등은 생물다양성의 손실과 열섬현상과 같은 많은 도시생활환경 생태문제의 원인으로 인식되고 있고 있는 데(Peng et al., 2019), 도시숲은 인위적 요소가 우점하는 도시환경에 다양한 생태계서비스와 환경적 혜택을 제공 하는 도시생태계의 가장 중요한 요소 중 하나로 부각되 고 있다(Achard, 2009). 이러한 환경적 편익에는 에너지 절약, 도시 열섬 효과의 감소, 공기와 수질 개선, 탄소 격 리 및 생물다양성 보존이 포함될 수 있다(Bergstedt, 1997). 즉 도시숲은 인위환경이 지배하는 도시지역에서 비생물 요소와 생물 요소의 안정성과 다양성을 유지 및 개선하는데 있어 마중물의 역할을 하고 있다. 그러나, 도 시숲은 산림생태계와 달리 자연적 요인보다는 인위적 요 인에 의해 그 조성과 구조 그리고 기능이 변화될 우려가 상존하는 다소 가변적인 생태계이다(Spies and Cline, 1988; McComb et al., 1993). 따라서, 다양한 환경압에 따 른 직·간접적 영향으로 양적 감소와 더불어 질적 쇠퇴가 점점 현저해지고 있는 도시환경의 생물적 및 비생물적 건강성과 안정성을 유지하고 개선하는데 있어서는 도시 숲의 생태적 특성과 그 공간분포 변화 경향을 주기적으 로 파악하는 것이 중요한 수단 중의 하나라고 할 수 있 다. 또한, 도시숲의 식생 유형과 그 변화 양상에 대한 지 리적 분포, 범위 및 경관 패턴을 기술하거나 식생조사 정 보를 공간적으로 묘사하기 위한 분석적 기반으로 종조성 적 식생단위에 의한 식물사회학적 식생도와 상층우점종 에 의한 상관식생도가 널리 활용되고 있다(Küchler and Zonneveld, 1988; Pedrotti, 2013). 식물사회학적 식생도는 국가식생단위체계가 정립되어 있는 유럽이나 일본 등에 서 주로 작성되고 있지만 우리나라의 경우 아직 국가식 생단위체계가 정립되어 있지 않아 대개 상층우점종의 상 관에 의한 상관식생도가 작성되고 있다(Oh et al., 2010;
Cho et al., 2016; Kim and Cho, 2017). 상관식생도는 첫 째, 모든 축척의 지도에 적용할 수 있으며, 둘째, 모든 국 가나 지역의 지도에서 적용할 수 있고, 셋째, 적용되는 식생유형의 이름이 명확하게 표현되며, 넷째, 식물사회학 적 전문지식이 요구되지 않기 때문에 누구나 쉽게 적용 할 수 있고, 식물사회학적 식생도 작성의 기반을 제공한 다는 점에서 식물사회학적 식생도에 비해 실무적 활용성 이 보다 높다고 할 수 있다. 최근 들어서는 초고해상도 위성영상 및 칼라 항공사진 활용 기술의 발달로 상관식 생 판독에 있어서 정밀성이 점점 높아져 가고 있어 상관 식생도의 잠재적 활용성은 보다 높아지고 있다(Feng et al., 2015; Han et al., 2016; Ishida et al., 2018). 이에 본
연구에서는 도심과 주거지에 연속된 자연녹지 공간이자 연간 약 1,300만명 이상의 이용객(Lee et al., 2010)으로 인해 산림지역에 대한 물리적 훼손 및 생물적 교란이 심 화되고 있어 산림생태계의 자연성, 고유성, 다양성, 그리 고 생태적 연속성 등이 급격히 저하되고 있는 대구광역 시의 대표적인 도시숲인 앞산공원 대상으로 산림식생의 체계적이고 효율적인 보전 및 관리 등 도시숲 생태계서 비스의 기반을 조성하는데 기본정보를 제공하기 위해 1997년과 2016년의 산림식생의 종조성, 구조 및 정밀식 생도에 의한 공간분포 특성을 비교·분석하였다.
재료 및 방법
1. 연구대상지
본 연구의 대상지역인 앞산공원은 대구광역시의 남쪽에 위치하고 주봉인 앞산(660.3m)과 주변의 산성산(653.4 m) 과 대덕산(583.5 m)을 포함하여 고산골, 달비골 등을 비롯 한 크고 작은 8개의 계곡과 20여 개의 약수터 및 수많은 등산로가 자리하고 있으며 총면적은 1,677.2 ha이다 (Daegu metropolitan city, 2019). 앞산공원이 위치한 대구 광역시는 우리나라 광역시 중 폭염에 가장 취약한 도시 (National Institute of Environmental Research and Ministry of Environment, 2012)로, 연평균 기후는 평균 기온 14.1℃
(1981~2010년), 평년값 기준 강수량 1,064.4 ㎜이며(Korea Meteorological Administration, 2018), 지질은 중생대 백악 기 말에 분출, 관입한 화산암 지대로 산정부는 안산암질각 력암으로 구성되어 있으며 동서방향으로 단층선이 나타나 고 있다(Park, 2011).
2. 조사 및 분석방법
본 연구의 기초라 할 수 있는 식생조사는 1997년과 2016년 5월에서 9월 사이에 동일한 표본 조사구에서 식물 사회학적 조사방법(Braun-Blanquet, 1964)으로 반복 실시 하였다. 표본 조사구의 수는 165개이며, 조사구 크기는 국 소환경과 상관식생의 군락 크기를 고려하여 10 m × 10 m 에서 20 m × 20 m를 적절히 적용하였다. 표본 조사구에 서는 층위별 구성종의 우점도와 군도를 비롯하여 국소환 경 특성(해발고, 사면경사도, 사면장위각, 암석노출도 등) 에 대한 정보를 수집 기록하였다.
야외에서 수집된 식생정보는 MS-Excel 2016 프로그램을 활용하여 DB화 한 자료를 상층 우점종을 기준으로 산림식 생 유형을 구분한 후, 식생유형간 구성종의 총피도(total coverage), 상대중요치(relative importance value; RIV), 생활 형 조성(life forms), 종다양성 지수(species diversity indices) 등을 각각 산출하였다. 상대중요치는 Curtis와 McIntosh
(1951)의 기준으로 산출하였으며 평균상대중요치(mean importance value; MIV)는 상대중요치에 식생 층위간 구성 종의 개체 크기를 고려하여 층위별로 가중치를 부여한 (교목 층 IV×3+아교목층×2+관목층×1+초본층×0.5)/6.5으로 산출 하였다(Lee et al., 2018). 생활형 조성은 Raunkiaer(1934) 의 휴면형(dormancy form)과 Numata (1947)의 산포형 (disseminule forms), 근계형(radicoid forms), 그리고 생육형 (growth forms) 구분 기준에 의해 비교하였다. 종다양성 지수 는 종풍부도(species richness index; S), 종다양도(Shannon diversity index; H’), 종균재도(Pielou evenness index; J’), 그리고 종우점도(Simpson dominance index; D)를 산출하였 다(Shanon and Weaver, 1949; Brower and Zar, 1977). 지표종 선정은 Dufrêne and Legendre(1997)의 IndVal(indicator value)방법을 적용하였으며, 군락유사성은 수치분류학에서 사용되는 연관 계수인 Jaccard 계수(Jaccard, 1901)를 사용하 여 PC-ORD 7 프로그램(McCune and Mefford, 2016)을 통해 비교·분석하였다. 식생유형의 공간분포 및 산림경관 요소 변화 경향을 파악하기 위한 정밀식생도는 수치지형도 (1/5,000), 고해상도 정사항공사진(National Geographic Information Institute, 2010) 및 수치임상도(1/5,000) 등을 바탕으로 일차적으로 상관식생구분도를 작성한 후, 현지 조사를 통하여 상관식생구분도상 상관식생의 유형을 동 정하고 그 경계를 확정하였으며, 실내에서 QGIS Desktop 2.18.15 프로그램(Qgis Development Team, 2017)을 이용 하여 작성하였다. 정밀상관식생도상 범례는 자연식생과
인공식생으로 구분한 후 모두 ○○군락으로 표시하였다. 식물 동정은 원색식물도감(Lee, 2003)을 참조하였으며, 그 명명 은 국가표준식물목록(Korea Forest Service, 2018) 기준을 적용하였다.
결과 및 고찰
1. 산림식생 유형
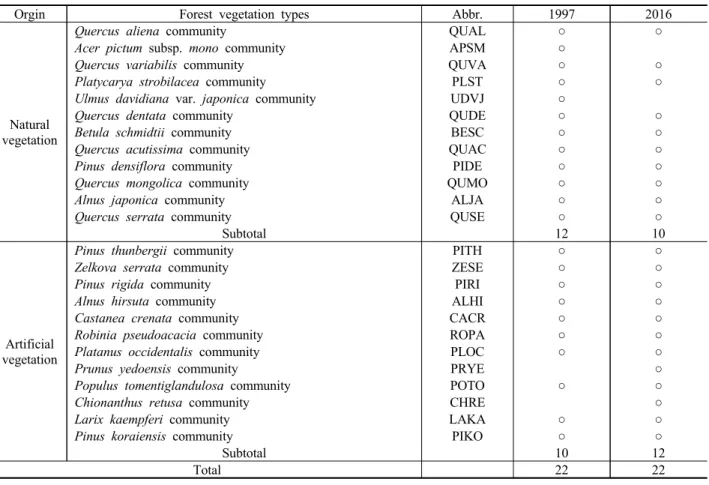
산림식생 유형의 경시적 변화 경향을 최상층 우점종의 상관(physiognomy)을 기준으로 비교한 결과, 1997년과 2016년 모두 22개 유형으로 나타났으나, 그 구성에 있어 서는 성립기원 간 다소 차이가 있었다(Table 1). 자연식생 의 경우 1997년 12개 유형에서 2016년 10개 유형으로 2개 유형(고로쇠나무군락, 느릅나무군락)이 감소하였는데, 두 유형 모두 개체군 크기가 자기 유지가 어려울 정도로 매 우 작아 인접 식생에 자연적으로 편입된 것이었다. 인공 식생의 경우 1997년 10개 유형에서 2016년 12개 유형으 로 2개 유형(왕벚나무군락, 이팝나무군락)이 증가하였는 데, 이는 경관숲 조성사업 등에 따른 영향으로 판단된다.
2. 산림식생의 구조적 특성 변화
산림식생의 구조적 특성 변화 경향을 파악하기 위하여 그 유형별 구성종의 단위면적당(/100㎡) 총피도, 상대중 요치, 종다양성 지수 등과 아울러 생활형 조성, 군락유사 성, 그리고 지표종 등을 비교 분석하였다.
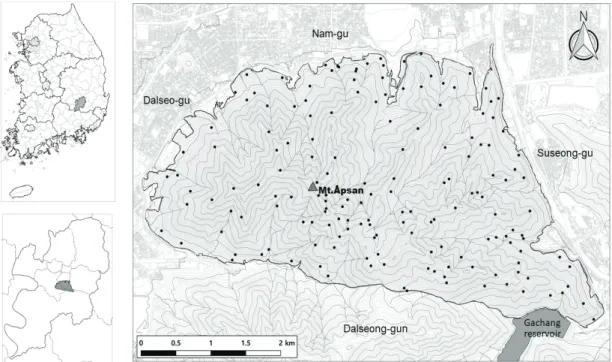
Figure 1. Location of the study site and field sample plots (black dots).
Orgin Forest vegetation types Abbr. 1997 2016
Natural vegetation
Quercus aliena community QUAL ○ ○
Acer pictum subsp. mono community APSM ○
Quercus variabilis community QUVA ○ ○
Platycarya strobilacea community PLST ○ ○
Ulmus davidiana var. japonica community UDVJ ○
Quercus dentata community QUDE ○ ○
Betula schmidtii community BESC ○ ○
Quercus acutissima community QUAC ○ ○
Pinus densiflora community PIDE ○ ○
Quercus mongolica community QUMO ○ ○
Alnus japonica community ALJA ○ ○
Quercus serrata community QUSE ○ ○
Subtotal 12 10
Artificial vegetation
Pinus thunbergii community PITH ○ ○
Zelkova serrata community ZESE ○ ○
Pinus rigida community PIRI ○ ○
Alnus hirsuta community ALHI ○ ○
Castanea crenata community CACR ○ ○
Robinia pseudoacacia community ROPA ○ ○
Platanus occidentalis community PLOC ○ ○
Prunus yedoensis community PRYE ○
Populus tomentiglandulosa community POTO ○ ○
Chionanthus retusa community CHRE ○
Larix kaempferi community LAKA ○ ○
Pinus koraiensis community PIKO ○ ○
Subtotal 10 12
Total 22 22
Table 1. Comparison of the forest vegetation types (based on physiognomy of the uppermost dominant species) in 1997 and 2016.
Orgin Physiognomic vegetation types Tree Subtree Shrub Herb All strata
97 16 97 16 97 16 97 16 97 16
Natural vegetation
(NV)
Quercus aliena community 72.0 75.2 20.9 23.3 32.0 55.5 21.7 14.5 140.0 168.5
Acer mono community 79.0 71.5 17.2 1.4 169.1
Quercus variabilis community 72.2 84.5 29.3 26.8 38.6 37.4 26.8 17.1 166.9 165.7
Platycarya strobilacea community 40.5 98.0 34.0 23.0 50.7 23.0 17.5 35.5 142.7 179.5
Ulmus davidiana var. japonica community 60.5 16.7 48.6 29.3 155.1
Quercus dentata community 70.0 62.3 32.2 32.6 50.8 61.9 40.1 29.8 193.0 186.6
Betula schmidtii community 79.4 80.0 47.1 54.0 40.1 62.5 18.4 0.4 184.9 196.9
Quercus acutissima community 72.5 74.6 25.4 23.6 34.6 61.7 38.0 26.2 170.4 186.1
Pinus densiflora community 70.2 82.7 24.3 23.8 38.8 47.7 23.2 23.1 156.5 177.3
Quercus mongolica community 65.4 82.9 26.2 30.9 46.6 55.2 18.9 20.2 157.1 189.1
Alnus japonica community 68.0 97.5 59.8 42.6 63.5 53.2 30.3 23.6 221.6 216.8
Quercus serrata community 61.8 19.9 50.9 36.4 168.9 186.1
Artificial vegetation
(AV)
Pinus thunbergii community 70.6 85.2 23.2 60.6 24.0 43.8 26.2 21.5 143.9 211.1
Zelkova serrata community 100.0 0.4 100.4
Pinus rigida community 69.6 63.9 56.0 13.5 203.1
Alnus hirsuta community 62.8 78.0 30.6 45.2 44.6 15.5 18.9 154.1 141.5
Castanea crenata community 60.5 80.3 54.0 34.9 46.6 49.1 7.3 25.3 168.4 189.5
Robinia pseudoacacia community 74.7 75.2 27.8 44.4 41.6 43.2 33.6 29.4 177.7 192.3
Platanus occidentalis community 87.5 23.0 68.1 61.7 240.3
Prunus yedoensis community 87.5 68.0 55.4 28.5 29.2 151.9 116.7
Populus tomentiglandulosa community 68.0 66.0 51.5 23.0 54.4 44.8 63.3 13.5 237.2 147.3
Chionanthus retusa community 100.0 11.0 56.0 167.0
Larix leptolepis community 73.5 70.1 45.1 55.8 96.7 51.0 6.8 17.7 222.1 194.7
Pinus koraiensis community 43.8 55.5 43.8 0.0 36.1 1.9 50.5 10.1 174.1 67.5
NV 68.4 80.3 26.4 28.5 40.8 51.5 24.7 21.8 160.3 182.0 AV 68.3 75.9 28.7 41.0 40.5 40.9 34.0 24.3 171.6 182.1 Total (NV+AV) 69.0 79.1 26.9 32.0 40.8 48.5 26.8 22.5 163.4 182.1 Table 2. Comparison of the average total coverage (%) by vegetation strata for the forest vegetation types in 1997 and 2016.
1) 총피도
산림식생 구성종의 총피도는 지난 20년간 전체적으로 약 11%가 증가하였고 자연식생(약 14%)이 인공식생(약 6%)에 비해 두 배 이상 높은 경향이었다. 층위 간에는 교 목층, 아교목층, 그리고 관목층이 대개 15~20%의 증가 율을 보인 반면 초본층은 약 16%의 감소율 보였다(Table 2). 이는 지난 20년 간 자연 천이에 따른 임분 층위의 발 달로 임내가 울폐되고 유기물이 집적되면서 초본층에 자 리하고 있던 호광성 또는 일년생의 선구식물들이 자연 감소하였기 때문인 것으로 생각된다. 식생 유형 간에는 인공식생인 곰솔군락이 가장 높은 증가율(약 47%)을 보
였으며, 은사시나무군락이 가장 높은 감소율(약 38%)을 보였다. 그러나 인위적 간섭이 상존하는 도시숲의 특성 상 이와 같은 총피도의 증감 경향은 보다 많은 후속 연구 가 담보되어야 보다 객관적인 평가가 가능할 것으로 생 각된다.
2) 상대중요치
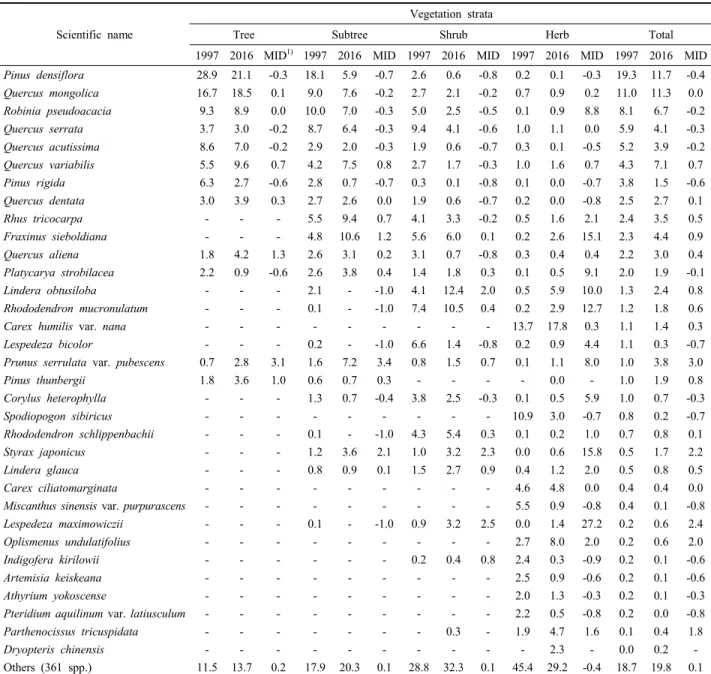
산림식생의 층위별 상위 10개 식물종을 중심으로 상대 중요치 변화 경향을 비교한 결과(Table 3), 전체 층위에 서의 구성종의 상대중요치 순위는 1997과 2016년 간에 큰 차이가 없었으나, 구성종의 개별 상대중요치에 있어
Scientific name
Vegetation strata
Tree Subtree Shrub Herb Total
1997 2016 MID1) 1997 2016 MID 1997 2016 MID 1997 2016 MID 1997 2016 MID Pinus densiflora 28.9 21.1 -0.3 18.1 5.9 -0.7 2.6 0.6 -0.8 0.2 0.1 -0.3 19.3 11.7 -0.4 Quercus mongolica 16.7 18.5 0.1 9.0 7.6 -0.2 2.7 2.1 -0.2 0.7 0.9 0.2 11.0 11.3 0.0
Robinia pseudoacacia 9.3 8.9 0.0 10.0 7.0 -0.3 5.0 2.5 -0.5 0.1 0.9 8.8 8.1 6.7 -0.2
Quercus serrata 3.7 3.0 -0.2 8.7 6.4 -0.3 9.4 4.1 -0.6 1.0 1.1 0.0 5.9 4.1 -0.3
Quercus acutissima 8.6 7.0 -0.2 2.9 2.0 -0.3 1.9 0.6 -0.7 0.3 0.1 -0.5 5.2 3.9 -0.2
Quercus variabilis 5.5 9.6 0.7 4.2 7.5 0.8 2.7 1.7 -0.3 1.0 1.6 0.7 4.3 7.1 0.7
Pinus rigida 6.3 2.7 -0.6 2.8 0.7 -0.7 0.3 0.1 -0.8 0.1 0.0 -0.7 3.8 1.5 -0.6
Quercus dentata 3.0 3.9 0.3 2.7 2.6 0.0 1.9 0.6 -0.7 0.2 0.0 -0.8 2.5 2.7 0.1
Rhus tricocarpa - - - 5.5 9.4 0.7 4.1 3.3 -0.2 0.5 1.6 2.1 2.4 3.5 0.5
Fraxinus sieboldiana - - - 4.8 10.6 1.2 5.6 6.0 0.1 0.2 2.6 15.1 2.3 4.4 0.9
Quercus aliena 1.8 4.2 1.3 2.6 3.1 0.2 3.1 0.7 -0.8 0.3 0.4 0.4 2.2 3.0 0.4
Platycarya strobilacea 2.2 0.9 -0.6 2.6 3.8 0.4 1.4 1.8 0.3 0.1 0.5 9.1 2.0 1.9 -0.1
Lindera obtusiloba - - - 2.1 - -1.0 4.1 12.4 2.0 0.5 5.9 10.0 1.3 2.4 0.8
Rhododendron mucronulatum - - - 0.1 - -1.0 7.4 10.5 0.4 0.2 2.9 12.7 1.2 1.8 0.6
Carex humilis var. nana - - - - - - - - - 13.7 17.8 0.3 1.1 1.4 0.3
Lespedeza bicolor - - - 0.2 - -1.0 6.6 1.4 -0.8 0.2 0.9 4.4 1.1 0.3 -0.7
Prunus serrulata var. pubescens 0.7 2.8 3.1 1.6 7.2 3.4 0.8 1.5 0.7 0.1 1.1 8.0 1.0 3.8 3.0
Pinus thunbergii 1.8 3.6 1.0 0.6 0.7 0.3 - - - - 0.0 - 1.0 1.9 0.8
Corylus heterophylla - - - 1.3 0.7 -0.4 3.8 2.5 -0.3 0.1 0.5 5.9 1.0 0.7 -0.3
Spodiopogon sibiricus - - - - - - - - - 10.9 3.0 -0.7 0.8 0.2 -0.7
Rhododendron schlippenbachii - - - 0.1 - -1.0 4.3 5.4 0.3 0.1 0.2 1.0 0.7 0.8 0.1
Styrax japonicus - - - 1.2 3.6 2.1 1.0 3.2 2.3 0.0 0.6 15.8 0.5 1.7 2.2
Lindera glauca - - - 0.8 0.9 0.1 1.5 2.7 0.9 0.4 1.2 2.0 0.5 0.8 0.5
Carex ciliatomarginata - - - - - - - - - 4.6 4.8 0.0 0.4 0.4 0.0
Miscanthus sinensis var. purpurascens - - - - - - - - - 5.5 0.9 -0.8 0.4 0.1 -0.8
Lespedeza maximowiczii - - - 0.1 - -1.0 0.9 3.2 2.5 0.0 1.4 27.2 0.2 0.6 2.4
Oplismenus undulatifolius - - - - - - - - - 2.7 8.0 2.0 0.2 0.6 2.0
Indigofera kirilowii - - - - - - 0.2 0.4 0.8 2.4 0.3 -0.9 0.2 0.1 -0.6
Artemisia keiskeana - - - - - - - - - 2.5 0.9 -0.6 0.2 0.1 -0.6
Athyrium yokoscense - - - - - - - - - 2.0 1.3 -0.3 0.2 0.1 -0.3
Pteridium aquilinum var. latiusculum - - - - - - - - - 2.2 0.5 -0.8 0.2 0.0 -0.8
Parthenocissus tricuspidata - - - - - - - 0.3 - 1.9 4.7 1.6 0.1 0.4 1.8
Dryopteris chinensis - - - - - - - - - - 2.3 - 0.0 0.2 -
Others (361 spp.) 11.5 13.7 0.2 17.9 20.3 0.1 28.8 32.3 0.1 45.4 29.2 -0.4 18.7 19.8 0.1
1) MID indicate the multiple increase and decrease
Table 3. Changes in the relative importance values of the major plant species of the forest vegetation types in 1997 and 2016.
서는 다소 간의 변화 경향을 보였다. 소나무와 리기다소 나무 등 침엽수종과 싸리, 억새, 큰기름새, 고사리 등 호 광성 선구식물 등은 0.5~1배 정도 감소한 반면, 잔털벚 나무, 조록싸리, 주름조개풀 등은 2배 이상 증가하였다.
식생 층위간에는 교목층과 아교목층의 경우 잔털벚나무 가 3배 정도, 관목층과 초본층의 경우 조록싸리, 생강나 무, 때죽나무, 쇠물푸레나무 등이 2~10배 정도의 매우 높은 증가율을 보였는데 이는 자연 천이와 인위적 간섭 이 복합적으로 영향을 미친 결과로 생각된다.
3) 종다양성
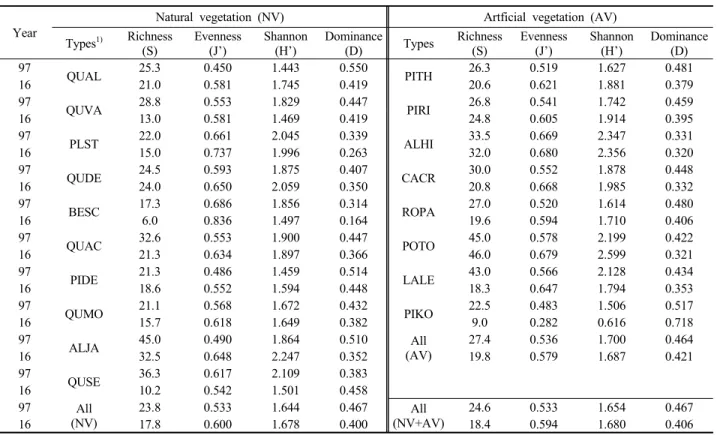
산림식생의 구성종의 단위면적당(/100㎡) 종풍부도(S) 와 종다양도(H’), 종균등도(J’), 그리고 종우점도(D)의 지 수 변화 경향을 평균값을 기준으로 비교하였다(Table 4).
종풍부도는 전체적으로 1997년 23.8종에서 2016년 17.8 종으로 6종이 감소하였다. 식생유형 간 증감률은 졸참나 무군락이 약 72%의 가장 높은 감소율을 보였으며, 박달 나무, 잣나무군락, 일본잎갈나무군락 및 굴참나무군락 등 도 50~70%의 비교적 높은 감소율을 보였다. 종균등도 는 전체적으로 1997년 0.533에서 2016년 0.594로 약 11%
증가하였고 식생유형 간에는 오리나무군락과 갈참나무 군락이 약 30%의 가장 높은 증가율을 보였고 잣나무군
락이 약 42%의 가장 높은 감소율을 보였다. 종다양도는 전체적으로 1997년 1.654에서 2016년 1.680으로 거의 유 사하였고 식생 유형 간에는 갈참나무군락과 오리나무군 락이 약 20%의 가장 높은 증가율을 보였고 잣나무군락 이 약 59%의 가장 높은 감소율을 보였다. 종우점도는 1997년 0.304에서 2016년 0.276으로 약 9%의 감소율을 보였다. 이를 종합하여 보면 앞산 산림식생 구성종의 종 다양성 변화 경향은 과거에 비해 우점종 및 균등도 및 생 태적 안정성도 약한 수준이지만 높아지는 방향으로 진행 됨을 알 수 있었다(Whittaker, 1965; Odum, 1969; Brower and Zar, 1977). 그러나 도시숲의 특성상 산림식생 성립 입지와 구성종에 대한 물리적 훼손 및 인위적 간섭이 지 속되고 있어 잠재적인 추세를 단언하기는 어려웠다.
4) 생활형 조성
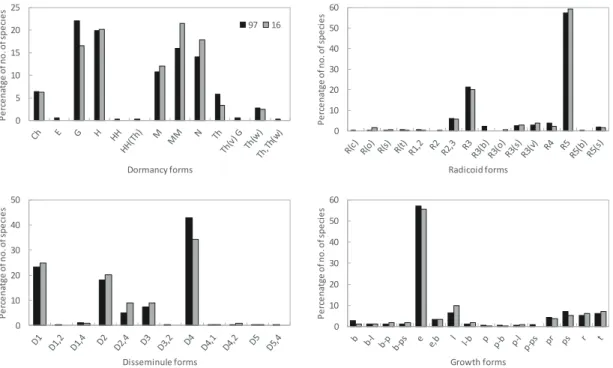
산림식생 구성종의 생활형 조성 중심 체계(휴면형-지 하기관형-산포기관형-생육형)는 1997년 ‘G-R5-D4-e’에서 2016년 ‘MM-R5-D4-e’로 휴면형에 있어서의 중심형이 과 거 지중식물(G)에서 대형지상식물(MM)로 변화된 경향이 었다(Figure 2). 휴면형의 경우 주로 대형지상식물(MM), 소형지상식물(M) 및 미소지상식물(N)이 10~30%의 증가 율을 보였고 나머지 유형들은 모두 감소 경향을 보였다.
Year
Natural vegetation (NV) Artficial vegetation (AV) Types1) Richness
(S) Evenness
(J’) Shannon
(H’) Dominance
(D) Types Richness
(S) Evenness
(J’) Shannon
(H’) Dominance (D) 97 QUAL 25.3 0.450 1.443 0.550
PITH 26.3 0.519 1.627 0.481
16 21.0 0.581 1.745 0.419 20.6 0.621 1.881 0.379 97 QUVA 28.8 0.553 1.829 0.447
PIRI 26.8 0.541 1.742 0.459
16 13.0 0.581 1.469 0.419 24.8 0.605 1.914 0.395 97 PLST 22.0 0.661 2.045 0.339
ALHI 33.5 0.669 2.347 0.331
16 15.0 0.737 1.996 0.263 32.0 0.680 2.356 0.320 97 QUDE 24.5 0.593 1.875 0.407
CACR 30.0 0.552 1.878 0.448
16 24.0 0.650 2.059 0.350 20.8 0.668 1.985 0.332 97 BESC 17.3 0.686 1.856 0.314
ROPA 27.0 0.520 1.614 0.480
16 6.0 0.836 1.497 0.164 19.6 0.594 1.710 0.406 97 QUAC 32.6 0.553 1.900 0.447
POTO 45.0 0.578 2.199 0.422
16 21.3 0.634 1.897 0.366 46.0 0.679 2.599 0.321 97 PIDE 21.3 0.486 1.459 0.514
LALE 43.0 0.566 2.128 0.434
16 18.6 0.552 1.594 0.448 18.3 0.647 1.794 0.353 97 QUMO 21.1 0.568 1.672 0.432
PIKO 22.5 0.483 1.506 0.517
16 15.7 0.618 1.649 0.382 9.0 0.282 0.616 0.718 97 ALJA 45.0 0.490 1.864 0.510 All
(AV)
27.4 0.536 1.700 0.464
16 32.5 0.648 2.247 0.352 19.8 0.579 1.687 0.421 97 QUSE 36.3 0.617 2.109 0.383
16 10.2 0.542 1.501 0.458 97 All
(NV)
23.8 0.533 1.644 0.467 All (NV+AV)
24.6 0.533 1.654 0.467
16 17.8 0.600 1.678 0.400 18.4 0.594 1.680 0.406
1)Abbreviation of types were shown Table 1.
Table 4. Comparison of species diversity indices among major 17 types of forest vegetation in 1997 and 2016.
지하기관형의 경우 뿌리가 지하에서 비스듬히 뻗어 있는 식물인 R(o)형과 다육경 식물인 R(s)형이 2~4배의 아주 높 은 증가율을 보였고 구경식물인 R(c)형, 가장 좁은 범위의 연결체를 갖는 근경식물의 변형체인 R3(b) 및 단립식물인 R5형은 모두 약 10%의 감소율을 보였다. 산포기관형의 경 우 중력산포형과 동물산포형의 복합체인 D4,2형이 약 2배 정도의 높은 증가율을 보였고, 풍수산포형과 동물산포형 의 복합체인 D1,2형, 자동산포형과 동물산포형의 복합체인
D3,2형 및 영양번식형과 중력산포형의 복합체인 D5,4형은 약 1배 정도의 감소율을 보였다. 생육형의 경우 덩굴형인 l형이 약 50%의 가장 높은 증가율을 보였고, 분지형인 b형 과 포복형과 위로제트형의 복합체인 p-ps형이 모두 60%
이상의 높은 감소율을 보였다.
5) 군락유사도
산림식생 구성종의 경시적 유사도를 Jaccard 계수(Jaccard
0 5 10 15 20 25
Percenatge of no. of species
Dormancy forms
97 16
0 10 20 30 40 50 60
Percenatge of no. of species
Radicoid forms
0 10 20 30 40 50
Percenatge of no. of species
Disseminule forms
0 10 20 30 40 50 60
Percenatge of no. of species
Growth forms
Figure 2. Changes in life-forms composition (dormancy, radicoid, disseminule, and growth forms) based on percentage of number of species in 1997 and 2016.
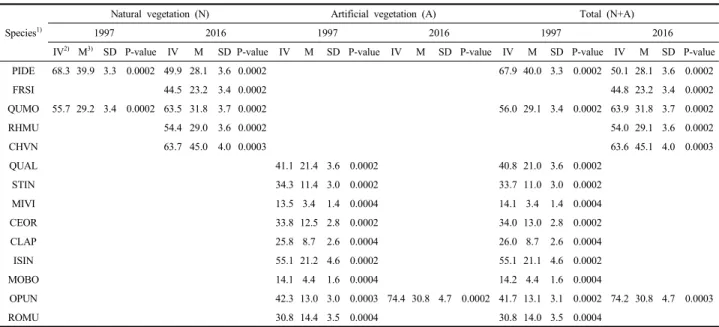
Figure 3. Jaccard similarity percentages of constituent species in major 18 types of forest vegetation in 1997 and 2016. Abbreviation of forest vegetation types were shown Table 1.
similarity coefficient)를 통해 분석한 결과(Figure 3), 1997년 과 2016년의 전체 산림식생 구성종의 유사도는 약 78%로 종구성이 거의 동일하게 나타났으며, 자연식생(약 79%)이 인공식생(약 69%)에 비해 10% 정도 높게 나타났다. 자연식 생과 인공식생 간 구성종의 유사도는 1997년 59%에서 2016 년 61%로 미미한 증가 경향을 보였다. 산림식생 유형 간에는 신갈나무군락이 약 82%의 가장 높은 유사도를 보였으며, 갈참나무군락, 굴참나무군락, 떡갈나무군락, 박달나무군락, 상수리나무군락, 소나무군락, 곰솔군락, 리기다소나무군락, 아까시나무군락 등이 50% 이상 유사도를 나타내어 종구성 에 큰 차이가 없는 유형으로는 분류되었다(Buell, 1966; Cox, 1996). 한편 식생 층위 간에는 교목층이 약 90%로 가장 유사도가 높았고, 하위 층위로 갈수록 유사도가 낮아지는 경향을 보였다.
6) 지표종
산림식생의 지표종 변화 경향을 Dufrene and Legendre (1977)의 IndVal(indicator value) 방법으로 분석한 결과 (Table 5), 1997년에는 소나무, 신갈나무, 산박하, 갈참나무 등 11종의 지표종이, 2016년에는 1997년 지표종인 소나무, 신갈나무, 주름조개풀 등과 당시 제외되었던 쇠물푸레나무, 진달래, 가는잎그늘사초 등 모두 6종이 유의한 지표종 (p<0.0005)으로 나타났다. 자연식생의 경우 1997년에는 소
나무, 신갈나무 등 2종이, 2016년에는 1997년 지표종과 당시 제외되었던 쇠물푸레나무, 진달래 등 4종이 유의한 지표종 으로 나타났으며, 인공식생의 경우 1997년 갈참나무, 국수나 무, 나도바랭이새, 노박덩굴, 사위질빵, 산박하, 산뽕나무, 주름조개풀, 찔레꽃 등 9종이, 2016년에는 1997년 지표종 중 주름조개풀 1종만이 유의한 지표종으로 나타났다. 이를 종합해서 살펴보면, 과거에 비해 임분 구조의 발달로 인한 생태적 안정성 높아지면서 호광성 및 덩굴성 식물 등 교란 임지에서 출현하던 식물들이 지표종에서 제외되고 소나무, 신갈나무, 쇠물푸레나무, 진달래, 가는잎그늘사초 등 온대 남부 산림식생을 특징짓는 식물들이 지표종으로 포함되는 변화 경향을 파악할 수 있었다.
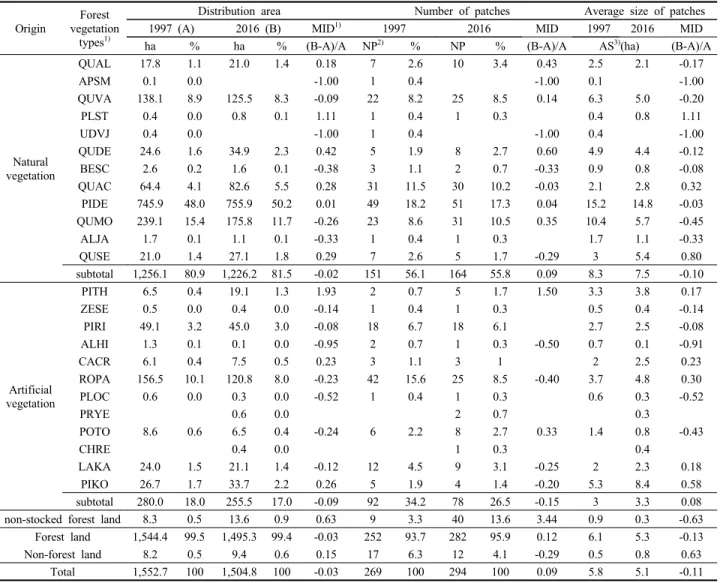
3. 산림식생 유형의 공간 분포 및 산림경관 요소
산림식생 유형의 경시적 공간 분포 변화 정도를 파악하기 위하여 대축척(1/5,000) 정밀식생도를 작성하고 유형별 분 포 면적과 패치 수(number of patches)를 비교하였다(Figure 4, Table 6). 정밀식생도 작성 면적은 1997년 1,552.7 ha에서 2016년 1,504.8 ha로 약 47.9 ha가 감소하였는데 이는 지난 20년간 저지대를 중심으로 외곽순환도로 개설 등 다양한 임지전용이 이루어진 결과로 생각된다. 산림식생의 공간 분포 면적 변화 경향을 살펴보면, 인공식생의 감소율(약 9%)이 자연식생의 감소율(약 2%)에 비해 약 4배 이상 높게
Species1)
Natural vegetation (N) Artificial vegetation (A) Total (N+A)
1997 2016 1997 2016 1997 2016
IV2) M3) SD P-value IV M SD P-value IV M SD P-value IV M SD P-value IV M SD P-value IV M SD P-value PIDE 68.3 39.9 3.3 0.0002 49.9 28.1 3.6 0.0002 67.9 40.0 3.3 0.0002 50.1 28.1 3.6 0.0002
FRSI 44.5 23.2 3.4 0.0002 44.8 23.2 3.4 0.0002
QUMO 55.7 29.2 3.4 0.0002 63.5 31.8 3.7 0.0002 56.0 29.1 3.4 0.0002 63.9 31.8 3.7 0.0002
RHMU 54.4 29.0 3.6 0.0002 54.0 29.1 3.6 0.0002
CHVN 63.7 45.0 4.0 0.0003 63.6 45.1 4.0 0.0003
QUAL 41.1 21.4 3.6 0.0002 40.8 21.0 3.6 0.0002
STIN 34.3 11.4 3.0 0.0002 33.7 11.0 3.0 0.0002
MIVI 13.5 3.4 1.4 0.0004 14.1 3.4 1.4 0.0004
CEOR 33.8 12.5 2.8 0.0002 34.0 13.0 2.8 0.0002
CLAP 25.8 8.7 2.6 0.0004 26.0 8.7 2.6 0.0004
ISIN 55.1 21.2 4.6 0.0002 55.1 21.1 4.6 0.0002
MOBO 14.1 4.4 1.6 0.0004 14.2 4.4 1.6 0.0004
OPUN 42.3 13.0 3.0 0.0003 74.4 30.8 4.7 0.0002 41.7 13.1 3.1 0.0002 74.2 30.8 4.7 0.0003
ROMU 30.8 14.4 3.5 0.0004 30.8 14.0 3.5 0.0004
1) Indicator species: PIDE Pinus densiflora, FRSI Fraxinus sieboldiana, QUMO Quercus mongolica, RHMU Rhododendron mucronulatum, CHVN Carex humilis var. nana, QUAL Quercus aliena, STIN Stephanandra incisa, MIVI Microstegium vimineum, CEOR Celastrus orbiculatus, CLAP Clematis apiifolia, ISIN Isodon inflexus, MOBO Morus bombycis, OPUN Oplismenus,undulatifolius, ROMU Rosa multiflora).
2) IV indicate indicator value(IndVal).
3) M indicate mean value.
Table 5. Indicator species analysis (IndVal) of the component species of forest vegetation in 1997 and 2016. Only species showing significant indication value (p<0.0005) are shown.
나타났으며, 식생 유형 간에는 굴피나무군락과 곰솔군락이 1~2배 정도의 높은 증가율을 보였다. 고로쇠나무군락, 느릅 나무군락 그리고 물오리나무군락은 인접식생에 편입되어 독립적인 군락 크기를 갖지 못하고 거의 소실된 상태이었다. 2016년 기준 산림식생 유형 간 공간 분포 면적은 소나무군락 이 전체 면적의 약 50%인 755.9 ha로 가장 크게 나타났고 다음으로 신갈나무군락 약 11.7%(175.8 ha), 굴참나무군락 약 8.3%(125.5 ha), 아까시나무군락 약 8.0%(120.8 ha), 그리 고 상수리나무군락 약 5.5% (82.6 ha) 등의 순이었다.
한편, 정밀식생도 상 구분된 모든 상관(상관식생유형+
무립목지+비산림지)의 개별 공간 형태(식생도상 경계가 구획된 각 구역)를 하나의 산림경관 요소 즉 패치(patch) 로 보고 그 공간적 변화 경향을 비교하였다. 분석결과,
전체 패치 수는 1997년 269개 패치에서 2016년 294개 패 치로 약 9%가 증가하였고, 자연식생은 약 9%의 증가율 을 그리고 인공식생은 약 15%의 감소율을 보였다. 식생 유형 간에는 인공식생인 곰솔군락이 약 1.5배의 가장 높 은 증가율을 보이는 가운데 떡갈나무군락, 갈참나무군락, 신갈나무군락 등 참나무과 유형들도 대개 40~60% 정도 의 비교적 높은 증가율을 보였다. 이와 같은 패치 개수의 증감 원인은 주로 자연 천이의 결과로 판단되었지만 일 부 인위적 간섭에 의한 것도 배제할 수 없다. 패치당 평 균 면적 변화 경향을 보면, 전체적으로 1997년 5.8 ha에서 2016년 5.1 ha로 약 0.7 ha가 감소하였다. 이상을 종합하 여 보면 과거에 비하여 패치 개수는 다소 증가하고 패치 당 평균 면적은 다소 감소하는 경향을 보여 향후 앞산공
Origin
Forest vegetation
types1)
Distribution area Number of patches Average size of patches 1997 (A) 2016 (B) MID1) 1997 2016 MID 1997 2016 MID ha % ha % (B-A)/A NP2) % NP % (B-A)/A AS3)(ha) (B-A)/A
Natural vegetation
QUAL 17.8 1.1 21.0 1.4 0.18 7 2.6 10 3.4 0.43 2.5 2.1 -0.17
APSM 0.1 0.0 -1.00 1 0.4 -1.00 0.1 -1.00
QUVA 138.1 8.9 125.5 8.3 -0.09 22 8.2 25 8.5 0.14 6.3 5.0 -0.20
PLST 0.4 0.0 0.8 0.1 1.11 1 0.4 1 0.3 0.4 0.8 1.11
UDVJ 0.4 0.0 -1.00 1 0.4 -1.00 0.4 -1.00
QUDE 24.6 1.6 34.9 2.3 0.42 5 1.9 8 2.7 0.60 4.9 4.4 -0.12 BESC 2.6 0.2 1.6 0.1 -0.38 3 1.1 2 0.7 -0.33 0.9 0.8 -0.08 QUAC 64.4 4.1 82.6 5.5 0.28 31 11.5 30 10.2 -0.03 2.1 2.8 0.32
PIDE 745.9 48.0 755.9 50.2 0.01 49 18.2 51 17.3 0.04 15.2 14.8 -0.03 QUMO 239.1 15.4 175.8 11.7 -0.26 23 8.6 31 10.5 0.35 10.4 5.7 -0.45
ALJA 1.7 0.1 1.1 0.1 -0.33 1 0.4 1 0.3 1.7 1.1 -0.33
QUSE 21.0 1.4 27.1 1.8 0.29 7 2.6 5 1.7 -0.29 3 5.4 0.80 subtotal 1,256.1 80.9 1,226.2 81.5 -0.02 151 56.1 164 55.8 0.09 8.3 7.5 -0.10
Artificial vegetation
PITH 6.5 0.4 19.1 1.3 1.93 2 0.7 5 1.7 1.50 3.3 3.8 0.17
ZESE 0.5 0.0 0.4 0.0 -0.14 1 0.4 1 0.3 0.5 0.4 -0.14
PIRI 49.1 3.2 45.0 3.0 -0.08 18 6.7 18 6.1 2.7 2.5 -0.08 ALHI 1.3 0.1 0.1 0.0 -0.95 2 0.7 1 0.3 -0.50 0.7 0.1 -0.91
CACR 6.1 0.4 7.5 0.5 0.23 3 1.1 3 1 2 2.5 0.23
ROPA 156.5 10.1 120.8 8.0 -0.23 42 15.6 25 8.5 -0.40 3.7 4.8 0.30
PLOC 0.6 0.0 0.3 0.0 -0.52 1 0.4 1 0.3 0.6 0.3 -0.52
PRYE 0.6 0.0 2 0.7 0.3
POTO 8.6 0.6 6.5 0.4 -0.24 6 2.2 8 2.7 0.33 1.4 0.8 -0.43
CHRE 0.4 0.0 1 0.3 0.4
LAKA 24.0 1.5 21.1 1.4 -0.12 12 4.5 9 3.1 -0.25 2 2.3 0.18 PIKO 26.7 1.7 33.7 2.2 0.26 5 1.9 4 1.4 -0.20 5.3 8.4 0.58 subtotal 280.0 18.0 255.5 17.0 -0.09 92 34.2 78 26.5 -0.15 3 3.3 0.08 non-stocked forest land 8.3 0.5 13.6 0.9 0.63 9 3.3 40 13.6 3.44 0.9 0.3 -0.63
Forest land 1,544.4 99.5 1,495.3 99.4 -0.03 252 93.7 282 95.9 0.12 6.1 5.3 -0.13 Non-forest land 8.2 0.5 9.4 0.6 0.15 17 6.3 12 4.1 -0.29 0.5 0.8 0.63
Total 1,552.7 100 1,504.8 100 -0.03 269 100 294 100 0.09 5.8 5.1 -0.11
1) MID indicate the multiple increase and decrease.
2) NP indicate the number of patches.
3) AS indicate average size of patches.
Table 6. Change of spatial distribution area, number of patches, and average size of patches by forest vegetation types of Apsan park in the year 1997 and 2016.
원 산림식생의 생태적 안정성 유지를 위해서는 단편화되 어 있는 패치들의 생태적 연결성을 확보해 주는 방향으 로 적절한 관리 대책 마련이 필요할 것으로 생각되었다.
결 론
도시숲은 인위환경이 지배하는 도시지역에서 비생물 요소와 생물 요소의 안정성과 다양성을 유지 및 개선하 는데 있어 마중물의 역할을 하고 있지만, 도시화에 따른 직·간접적 영향으로 양적 감소와 더불어 질적 쇠퇴가 점 점 현저해지고 있다. 대구광역시의 대표적인 도시숲인 앞산공원은 연간 1,300만명에 달하는 이용객의 수가 말 해주듯이 과도한 이용압으로 생태적 측면에서 도시숲으 로서의 정상적인 기능 유지에 있어서 여러 가지 문제가 현존하고 있어, 이의 개선을 위한 대책 마련이 시급한 실 정이다. 이에 본 연구는 앞산공원이 산림생태계로서 기 능을 유지할 수 있는 기반인 산림식생을 대상으로 1997 년과 2016년에 그 조성 및 구조 그리고 공간 분포의 변화 정도를 비교 연구하였다. 그 결과 지난 20년간 식생 유형 의 수는 모두 22개로 나타났지만 그 구성 종류에 있어서 는 다소 간의 변화가 있었고, 단위면적당 총피도는 증가 한 반면 출현종수는 다소 감소하였다. 이는 Lee et al.
(1994; 2012)이 서울시의 대표적 도시숲인 관악산을 대상 으로 수행한 식물군집 구조 변화 연구에서 경시적 종수 감소 원인을 토양산성화 등 지속적인 환경악화에 의한 것으로 분석한 것과 동일한 결과라 할 수 있다. 그러나 도시숲은 모암, 성립과정, 지리적 위치, 면적, 교란정도, 입지 환경 특성 등에 따라 식생구조와 조성 그리고 천이 단계가 다르므로 지속적인 환경악화가 직접적인 종수 감 소를 초래하였다고 판단하기에는 추후 많은 논의가 있어 야 될 것으로 생각된다. 한편, 자연식생 구성 군락 중에 서 과거에 비해 2개 군락이 인접식생의 영향으로 감소한
것으로 나타나는데, 이들은 대상지역의 고유식생으로 도 시숲의 생태적 온전성 또는 다양성을 유지하는 측면에서 지속적인 모니터링을 통한 생육환경 개선으로 자기 유지 가 가능한 크기의 개체군으로 유도하는 보다 적극적인 현지 내 보전대책을 수립할 필요가 있다고 판단된다. 산 림식생의 공간 분포 면적은 인공식생이 자연식생에 비해 약 4배 이상 높은 감소율을 나타내었고, 그리고 산림경관 패치 수는 약 9% 증가한 반면 그 평균 크기는 약 12%
감소된 경향을 알 수 있었다. 이러한 결과는 현시쯤에서 앞산공원의 생태적 보전 및 관리 방안을 마련하는 데 있 어 하나의 기초자료로 활용성이 있을 것으로 보인다. 그 러나 인위적 간섭과 환경적 가변성이 상존하는 도시숲의 특성상 지속적인 후속 연구가 이루어져야 그 객관성이 보다 담보될 것으로 판단된다.
감사의 글
본 연구는 2016년도 국립산림과학원 위탁연구과제 “대 구 앞산의 식생구조 변화 조사 및 정밀식생도 작성” 연구 에 의해 수행되었다.
References
Achard, F. 2009. Vital forest graphics. Arendal, Norway: UNEP/
GRID-Arendal. http://www.grida.no/_res/site/file/publicati on/vital_forest_graphics.pdf(2019.2.15.).
Bergstedt, J. 1997. Theory of nature conservation. In: Handbook Applied Biotope Protection II-3, 10. Erg.Lfg. 10(97): 3-10.
Braun-Blanquet, J. 1964. Plant Sociology, Essentials of Vegetation Science. 3rd ed. Springer-Verlag. Wien, New York, USA. pp. 865.
Brower, J.E. and Zar, J.H. 1977. Field and Laboratory Methods for General Ecology. Wm. C. Brown Company. Iowa, USA.
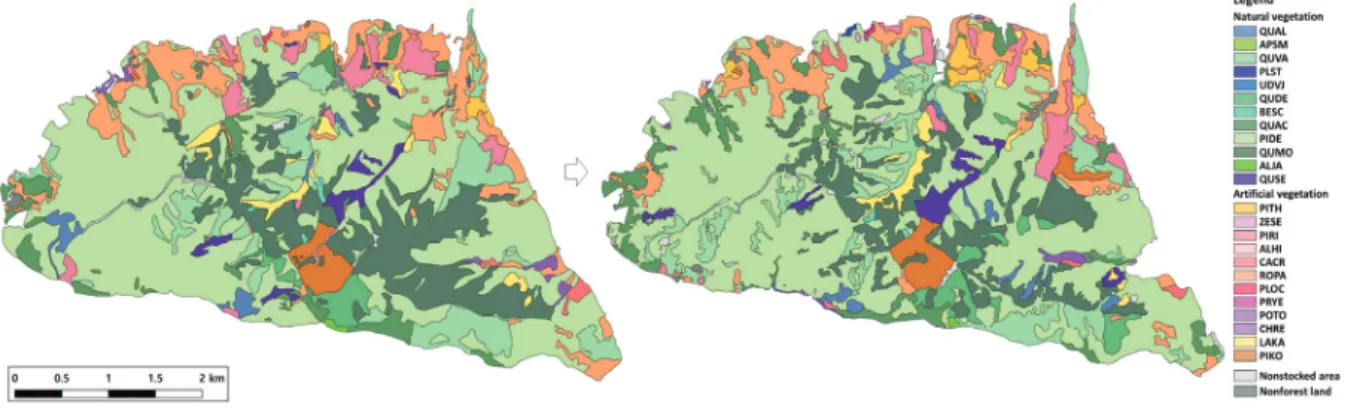
Figure 4. A detailed forest vegetation map of Apsan park
(Abbreviations of the name for forest vegetation types were shown Table 1).
pp. 288.
Buell, M.F., Langford, A.N., Davidson, D.W. and Ohmann, L.F. 1966. The upland forest continuum in northern New Jersey. Ecology 47: 416-432.
Cho, J.H., Park, C.R., Oh, J.H., Kim, J.S. and Cho, H.J. 2016.
Changes in Vegetation Characteristics Over Time in the Isolated Forests of the Urban Areas: A Case Study on the Mt. Hwangyeong, Busan Metropolitan City. Journal of Korean Forest Society 105(3): 284-293. (in Korean with English abstract)
Cox, G.W. 1996. Laboratory Manual of General Ecology. 7th ed. William C. Brown Company Publishers, Dubuque, Iowa.
pp. 278.
Curtis, J.T. and Mcintosh, R.P. 1951. An upland forest continuum in the prairie-forest border region of Wis-consin. Ecology 32:4 76-498.
Daegu metropolitan city. 2019. http://www.daegu.go.kr/cts/inde x.do(2019.2.18.). (in Korean)
Dufrêne, M. and Legendre, P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67(3): 345-366.
Feng, Q., Liu, J. and Gong, J. 2015. UAV Remote Sensing for Urban Vegetation Mapping Using Random Forest and Texture Analysis. Remote Sensing 7(1): 1074-1094.
Han, Y.G., Jung, S.H. and Kwon, O.S. 2016. How to utilize vegetation survey using drone image and image analysis software. Journal of Ecology and Environment 41(1):18.
(in Korean with English abstract)
Ishida, T., Kurihara, J., Viray, F.A., Namuco, S.B., Paringit, E.C., Perez, G.J., Takahashi, Y. and Marciano, J.J. 2018.
A novel approach for vegetation classification using UAV-based hyperspectral imaging. Computers and Elec- tronics in Agriculture 144: 80-85.
Jaccard, P. 1901. Distribution of alpine flora in the Dranses basin and in some neighboring regions. Bulletin of the Vaudoise Society of Natural Sciences 37: 241-272.
Kim, H.Y. and Cho, H.J. 2017. Vegetation Composition and Structure of Sogwang-ri Forest Genetic Resources Reserve in Uljin-gun, Korea. Korean Journal of Environment and Ecology 31(2): 188-201. (in Korean with English abstract) Korea Forest Service. 2018. Korean Plant Names Index.
http://www.nature.go.kr/kpni/index.do (2019.1.12.). (in Korean)
Korea Meteorological Administration. 2018. Climatological data. http://www.kma.go.kr (2018. 2. 11.). (in Korean) Küchler, A.W. and Zonneveld, I.S. 1988. Vegetation mapping.
Dr W. Junk, Dordrecht. pp. 635.
Lee, H.Y., Park, I.H., Cho, K.J. and Jang, G.S. 2010. A Study on the Damage Condition of the Major Trail in Apsan
Park, Daegu Metropolitan City. Proceedings of the 2010 Meeting of Korean Institute of Landscape Architecture 2010:
183-186. (in Korean)
Lee, J.E., Shin, J.K., Kim, D.K. and Yun, C.W. 2018.
Classification of Forest Vegetation for Forest Genetic Resource Reserve Area in Heuksando sland. Korean Journal of Environment and Ecology 32(3): 289-302. (in Korean with a English abstract)
Lee, K.J., Song, K.J. and Cho, W. 1994. Changes of the Plant Community Structure during the Twenty-two Years (1972~1993) in Forest of Mt. Kwanak. Journal of Korean Institute of Landscape Architecture 22(3): 79-90. (in Korean with English abstract)
Lee, K.J., Choi, J.W. and Noh, T.H. 2012. Change of the Plant Community Structure during the Thirty-nine Years (1972~
2010) in Forest of Mt. Kwanak. Proceedings of the 2010 Conference of Korean Society of Environment and Ecology 22(1): 68-72. (in Korean)
Lee, T.B. 2003. Coloured flora of Korea, Hyangmunsa press, Seoul. pp. 910. (in Korean)
McComb, W.C., Spies, T.A. and Emmingham, W.H. 1993.
Douglas-fir forests: managing for timber and mature-forest habitat. Journal of Forestry 91(12): 31-42.
McCune, B. and Mefford, M.J. 2016. PC-ORD. Multivariate analysis of Ecological Data, Version 7.0 for Windows. MjM Software, Oregon, U.S.A.
MjM Software Design. 2016. PC-ORD Multivariate Analysis of Ecological Data(Version 7).
National Geographic Information Institute. 2010. High Resolution Airborne Digital Ortho Imagery(Daegu sheet). National Geographic Information Institute. Korea.
Numata, M. 1947. Ecological judgement of grassland condition and trend: I. Judgement by biological spectra. Grassland Science 11: 20-33.
Odum, E. P. 1969. The strategy of ecosystem development.
Science 164: 262-270.
Oh, H.K., Kahng, B.S., Yu, B.H. and Song, J.Y. 2010. Vegetation Analysis to Detailed Hierarchical Vegetation Map Using Geographic Information Systems and Remote Sensing -A Case Study on the Jirisan National Park-. Proceedings of the 2010 Conference of Korean Society of Environment and Ecology 20(1): 45-48. (in Korean)
Park, J.S. 2011. Geo Trail Planning of Mt. Apsan. Research Report of Daegu-Gyeongbuk Development Institute 2011(24): 27-28. (in Korean)
Pedrotti, F. 2013. plant and vegetation mapping. Springer, Heidelberg. pp. 292.
Peng, Y., Mi, K., Wang, H., Liu, Z., Lin, Y., Sang, W. and Cui, Q. 2019. Most suitable landscape patterns to preserve
indigenous plant diversity affected by increasing urbanization: A case study of Shunyi District of Beijing, China. Urban Forestry & Urban Greening, 38, 33-41.
QGIS Development Team. 2017. QGIS ver. 2.18.15. http://www.
qgis.org/ko/site /forusers/download.html(2018.1.11.).
Raunkiaer, C., 1934. Life Form of Plants and Statistical Plant Geography, Charendon Press, Oxford Univ. Press, London.
pp. 632.
Shannon, C.E. and Weaver, W. 1949. The mathematical theory of communication. University of Illinois. Illinois, USA.
pp. 144.
Spies, T.A. and Cline, S.P. 1988. Coarse wood debris in mani-
pulated and unmanipulated coastal Oregon forests. From the Forest to the Sea: A Story of Fallen Trees. Pacific North-west Research Station., Gen. Tech. Rep. PNW-GTR- 229: 5-23.
Whittaker, R. H. 1965. Dominance and Diversity in Land Plant Communities: Numerical relations of species express the importance of competition in community function and evolution. Science 147(3655): 250-260.
Manuscript Received : March 6, 2019 First Revision : March 27, 2019 Accepted : April 2, 2019