Kor. J. Hort. Sci. Technol. 29(6):511-522, 2011
플라보노이드 대사 조절을 통한 화색 변경
임선형1ㆍ김재광2ㆍ김동헌3ㆍ손성한4ㆍ이종렬1ㆍ김영미1ㆍ하선화1*
1국립농업과학원 기능성물질개발과, 2국립농업과학원 생물안전성과, 3국립농업과학원 신작물개발과, 4국립농업과학원 유전자분석개발과
Flower Color Modification by Manipulating Flavonoid Biosynthetic Pathway
Sun-Hyung Lim
1, Jae Kwang Kim
2, Dong Hern Kim
3, Seong-Han Sohn
4, Jong-Yeol Lee
1, Young-Mi Kim
1, and Sun-Hwa Ha
1*1Functional Biomaterial Division, National Academy of Agricultural Science, Suwon 441-707, Korea
2Biosafety Division, National Academy of Agricultural Science, Suwon 441-707, Korea
3Bio-crop Development Division, National Academy of Agricultural Science, Suwon 441-707, Korea
4Genomics Division, National Academy of Agricultural Science, Suwon 441-707, Korea
Abstract. Flower color is one of the main target traits in the flower breeding. Recently, technological advances in genetic engineering have been successfully reported the flower colors, such as blue roses and blue carnations that are impossible to develop by traditional breeding. Accumulated knowledge-based approaches for flavonoid biosynthesis enabled to introduce novel and unique colors into flowers. These flower color modifications have been made through the regulation of flavonoid metabolic pathway - control of endogenous gene expression and introduction of foreign genes to produce novel and specific flavonoids - and the introduction of transcription factors that are known to regulate sets of genes being involving in the flavonoid biosynthetic pathway. More empirical regulation of the flavonoids metabolism requires the understanding for regulatory mechanism of intrinsic flavonoids depending on the flower crops and the very sophisticated control of flavonoid metabolic flow. In this review, we summarized successful examples of flower color modification. It might be useful to deduce the strategy for the creation of exquisite colors in flower plants.
Additional key words: anthocyanin, genetically modified, metabolic engineering, transcription factor
*Corresponding author: [email protected]
※ Received 5 September 2011; Accepted 28 December 2011. 본 연구는 농촌진흥청 농업과학기술 연구개발사업 (PJ006834)과 바이오그린 21 사업(PJ008184)의 공동 지원에 의해 이루어진 것임
서 언
화훼류는 원예산업의 중요한 부분으로 유럽, 북아메리카, 일본시장에서 년간 1,500억 달러의 시장을 형성하고 있으며 특히 코스타리카, 콜롬비아, 케냐, 에티오피아, 에콰도르 등 여러 개발도상국에서의 화훼산업은 그 나라 경제에 중요한 부분을 차지하고 있다(Chandler and Tanaka, 2007). 화훼는 그 용도에 따라 수확된 꽃 자체를 이용하는 절화(cut flower) 와 집과 정원 장식을 목적으로 하는 분화(potted flower)로 나눌 수 있다. 현재 가장 많이 유통되는 절화 및 분화용 화훼 류는 Table 1에 정리하였고, 이들의 다양한 화색에 영향을 주는 주요한 색소물질도 함께 나타내었다.
자연계에는 다양한 형태와 색깔을 지닌 많은 종류의 꽃이
존재한다. 이러한 다양성은 풍매수분을 촉진하거나 수분매
개자를 끌어들여 성공적인 번식을 보장하기 위한 식물의 진
화산물이다. 피자식물의 화색은 곤충이나 새와 같은 수분매
개자를 끌어들이는 매우 중요한 요인으로 작용한다. 육종적
인 측면에서도 화색은 식물 육종가의 주요한 목표 형질이며 ,
자연 돌연변이나 유전적으로 연관된 종과 교배를 통하여 다
양한 색깔을 갖는 품종들이 육종되어 왔다. 그러나 전통적
인 육종 방법으로는 파란장미와 같이 특정 화색을 갖는 품
종을 육종하는 것이 불가능하다. 전통적인 육종 방법은 원
하는 화색을 지닌 같은 종이나 근연 종내의 개체와 기존의
품종을 교잡하여 원하는 화색을 지닌 식물을 개발하는 것인
데, 유전적으로 원연관계에 있는 종을 이용해야 하는 경우 에는 생식 장벽으로 인해 전통적인 육종방법을 이용하여 새 로운 화색을 도입하는 것이 매우 어렵다. 배배양과 원형질 체 융합과 같은 식물세포 및 조직배양기술로 생식장벽을 극 복하는 것이 기술적으로 가능하기는 하나 이러한 기술은 모 든 식물에 적용 가능한 것이 아니다.
화훼류에서 화색변경은 유전자 변형(genetically modified:
이하 GM) 기술이 가장 널리 적용된 분야이고, GM 화훼 작 물 개발연구의 많은 부분이 화색을 변경시키는 기술이다 (Nishihara and Nakatsuka, 2010; Tanaka et al., 2005, 2009).
화색을 결정하는 식물 색소에는 플라보노이드(flavonoid), 카로티노이드(carotenoid), 베타라인(betalain)이 있다. 플라 보노이드는 수용성물질이고 노란색에서 빨간색, 청색에 이 르는 넓은 영역의 색을 나타낸다. 카로티노이드는 탄소 40 개로 이루어진 테트라테르펜(tetraterpenoid)계 지용성 물질 로서, 색소체에 위치하여 꽃에서는 주로 노란색에 관여하지 만, 장미나 국화 같은 경우에는 안토시아닌 색소와 함께 주 황색/빨간색, 청동색/갈색의 색을 나타내며, 베타라인은 상 아색, 노란색, 주황색, 빨간색, 보라색을 나타내는 색소이다.
따라서 이러한 대사산물의 축적은 다양한 화색 개발을 위한 가장 중요한 목표형질로 고려되어 왔다.
이 중 플라보노이드는 식물 이차 대사산물 가운데 가장 많이 연구된 물질 중 하나이며, 생합성 경로는 모델작물인 애기장대 뿐만 아니라 옥수수, 페튜니아 등 다양한 식물에 서 보고되었다(Grotewold, 2006; Koes et al., 2005). 생합성 경로의 분자생물학적인 지식을 기반으로 페튜니아에 옥수 수의 dihydroflavonol 4-reductase(DFR)을 도입하여 화색이 변경된 것이 최초의 보고이다(Meyer et al., 1987). 그 이후 장미와 카네이션과 같은 중요한 화훼작물을 포함하여 새로 운 화색을 지닌 식물체를 만들기 위한 다양한 시도들이 있었 다. 카로티노이드와 베타라인도 중요한 화색소임에도 불구하 고, 카로티노이드의 생합성 대사경로를 조절하여 화색변경이 된 것은 몇 가지 예만이 있고, 베타라인의 경우에는 보고된 바가 거의 없다. 카로티노이드는 엽록체에서 식물 광합성에 중요한 기능을 수행하는 식물 색소이며, 앱시스산(abscisic acid), 스트리고락톤(strigolactone), 지베렐린(gibberellins)과 같은 식물호르몬 합성의 전구체로서 이들의 유전자 조작은 상대적으로 어렵다(Grotewold, 2006; Umehara et al., 2008).
이에 비해 플라보노이드 물질은 식물 생존에 필수적인 것은 아니며, 대사경로 조절을 통한 변경은 상대적으로 쉬운 것 으로 알려져 있다. 베타라인 또한 식물생존에 필수적인 물 질은 아니지만, 생합성 경로가 밝혀져 있지 않고 석죽목 (Caryophyllales)의 한정된 식물 종에서만 존재하기 때문에
베타라인을 이용한 화색 변경보고는 아직까지는 없다. 본 총설에서는 플라보노이드 색소관련 유전자조작을 통하여 화훼작물의 화색을 변경시킨 최근 연구들을 요약하였고, 담 배와 애기장대 같은 모델작물의 기초연구를 바탕으로 화훼 작물의 화색변경을 위한 가능성 있는 방법들을 언급하고자 한다.
식물의 플라보노이드 생합성 경로
플라보노이드는 식물 특이적인 이차 대사산물인 리그닌 (lignin), 스틸베노이드(stilbenoids), 쿠마린(coumarins)과 파 이토알렉신(phytoalexins)을 합성하는 페닐프로파노이드 (phenylpropanoid) 생합성 경로로부터 합성된다. 전형적인 플라보노이드 생합성 경로는 Fig. 1에 나타내었다. 간략하게 살펴보면, chalcone synthase(CHS)에 의해서 1개 분자의 coumaroyl-CoA와 3개 분자의 malonyl CoA가 축합반응으 로 노란색을 띄는 4,2’,4’,6’-tetrahydroxy chalcone을 형성하 게 된다. Chalcone을 기질로 하여 여러 효소들에 의해 가수 화, 환원, 산화반응을 거치면서 붉은색에서 파란색을 띄는 안토시아닌 아글리콘(예로 펠라고니딘(pelargonidin), 펠라 고니딘(cyanidin), 델피리딘(delphinidin))이 합성된다. 파란 색 안토시아닌의 생성은 안토시아니딘 B링의 가수화 수가 증가됨에 따라 짙어진다. 펠라고니딘계열의 색소(B링의 가 수화 1개)는 주황색 또는 붉은 색을 나타내며, 시아니딘계열의 색소 (B링의 가수화 2개)는 붉은색 또는 마젠타 색을 나타내며, 델피리딘계열의 색소 (B링의 가수화 3개)는 보라색 또는 파란 색을 나타낸다. 일반적으로 chalcone과 안토시아니딘은 당화 (glycosylations), 말로닐화(malonylations), 아실화(acylations) 등의 여러 가지 수식작용을 거친 다음 액포 내로 저장된다 . 금 어초의 노란색은 chalcone 4’-O-glucosyltransferase(C4’GT)와 aureusidin synthase(AS) 효소 반응에 의해 노란색의 aureusidin 6-O-glucoside가 생성되어 나타난다(Nakayama et al., 2000;
Ono et al., 2006). 또한 Sinningia cardinalis와 같은 몇몇 종은 플라바논(flavanone)으로부터 여러 단계를 거쳐서 주황색이나 붉은색을 띄는 3-데옥시안토시아니딘(3-deoxyanthocyanidin) 을 합성한다(Nakatsuka and Nishihara, 2010; Winefield et al., 2005).
위에 언급된 착색 플라보노이드 이외에 플라본(flavones) 과 플라보놀(flavonol)도 화색에 기여하는 것으로 알려졌다.
이들 색소는 무색이거나 옅은 노란 색조의 플라보노이드로
구성된 ‘조색소’라고 불리며 안토시아닌 색소의 파란색으로
의 발달과 안정성에 중요한 역할을 하는 것으로 알려져 있
다. 플라본과 플라보놀은 플라바논과 디하이드로플라보놀
(dihydroflavonol)을 기질로 하여 flavone synthase(FNS)와
Fig. 1. Flavonoid biosynthetic pathway and flavonoid compounds accumulated in flowers. A simplified pathway derived from several plant species is depicted for ease explanation. The painted colors show image of each compound. ANS, anthocyanidin synthase;
AS, aureusidin synthase; AT, acyltransferase; C4′GT, chalcone 4′-O-glucosyltransferase; CHI, chalcone isomerase; CHS, chalcone synthase; DFR, dihydroflavonol 4-reductase; F3H, flavanone 3-hydroxylase; F3′H, flavonoid 3′-hydroxylase; F3′5′H, flavonoid 3′,5′-hydroxylase; FLS, flavonol synthase; FNR, flavanone 4-reductase; FNS, flavone synthase; GT, glycosyltransferase; MT, methyltransferase.
flavonol synthase(FLS)에 의해서 합성된다(Fig. 1).
모델식물의 연구를 통해서 플라보노이드 생합성 유전자들 의 발현은 MYB, basic helix-loop-helix(bHLH), WD40 전사인 자의 중합체에 의해서 조절되는 것으로 보고되고 있다(Davies, 2009; Grotewold, 2005; Koes et al., 2005; Quattrocchio et al., 2006). 이러한 삼중 중합체 전사인자는 금어초, 페튜니아, 나팔꽃, 거베라, 용담 등을 포함한 화훼작물의 꽃잎에서 색 소발달에 영향을 주는 것으로 알려져 있다(Davies, 2009;
Morita et al., 2006; Nakatsuka et al., 2008c).
플라보노이드 생합성 대사경로 조절을 통한 화색 변경 플라보노이드 대사 경로를 조절하여 화색을 변경하기 위 해서는 내재 플라보노이드 경로를 조절하는 방법과 새로운 플라보노이드 합성 또는 특정 플라보노이드 생산을 위해 외
래 유전자를 도입하는 방법이 있고, 플라보노이드 대사 전 체를 조절하는 전사인자 조절을 이용한 방법이 보고되고 있 다. 현재까지 제시된 전략으로 화색이 변경된 다양한 사례 들을 보고하고자 한다(Fig. 2 and Table 2).
내재 플라보노이드 대사조절을 통한 화색 변경
플라보노이드 생합성 경로의 주요 유전자를 발현 억제시켜
서 화색을 변경시킨 다양한 보고가 있다. 플라보노이드 생합
성 경로의 상위유전자인 CHS 유전자를 역방향으로 발현시켜
담배와 페튜니아의 화색을 하얀색으로 변경하는데 성공하였
다(van der Krol et al., 1988). 내재 CHS, anthocyanidin
synthase(ANS) 유전자를 역방향으로 각각 발현시켜서 짙은
파란색 용담 (gentian)은 하얀색으로, 짙은 파란색의 토레니아
(torenia)는 옅은 파란색을 지니게 되었다(Fig. 2A; Nakamura
Table 1. The most widely traded cut-flowers and flowering pot plant species. Listed species produce one or more of the major group of anthocyanins, with reference to the three principal anthocyanidin types, pelagonidin, cyanidin, and delphinidin.
Plant species Uses Type of anthocyanidin
African violet Potted flower Cyanidin, delphinidin
Alstromeria Cut flower Cyanidin, delphinidin
Azalea Potted flower Cyanidin, delphinidin
Begonia Potted plant Cyanidin
Carnation Cut and potted flower Cyanidin, pelargonidin
Chrysanthemum Cut and potted flower Cyanidin
Cyclamen Potted flower Cyanidin, delphinidin
Freesia Cut flower Cyanidin, delphinidin, pelargonidin
Gerbera Cut flower Cyanidin, pelargonidin
Hortensia Potted flower Cyanidin, delphinidin
Hypericum Potted flower Cyanidin
Iris Cut flower Cyanidin, delphinidin
Kalanchoe Potted flower Cyanidin, delphinidin
Lily Potted and cut flower Cyanidin
Lisianthus Cut flower Delphinidin, pelargonidin
Pelargonium Potted flower Delphinidin, pelargonidin
Petunia Potted flower Cyanidin, delphinidin
Phalaenopsis Potted flower Cyanidin, delphinidin
Poinsettia Potted flower Cyanidin, pelargonidin
Primula Potted flower Delphinidin, pelargonidin
Rose Cut and potted flower Cyanidin, pelargonidin
Tulip Cut flower Cyanidin, delphinidin, pelargonidin
et al., 2006; Nishihara et al., 2006; Ueyama et al., 2006).
내재 F3’5’H 유전자를 발현 억제하여 보라색의 니에렘베르 기아(nierembergia)는 옅은 파란색으로, 보라색의 시클라멘 (cyclamen)은 빨간색 또는 분홍색으로 변경되었다(Fig. 3A;
Boase et al., 2010).
최근 대사조절 방법으로 RNA interference(RNAi)를 이용 하는 방법이 사용되고 있다. RNAi 방법을 이용하여 토레니 아와 담배에서 각각 CHS와 chalcone isomerase(CHI) 발현을 억제함으로써 화색 변경이 가능함을 보고하였다(Fukusaki et al., 2004; Nishihara et al., 2005). 짙은 파란색의 용담은 내 재 F3’5’H 유전자를 발현억제시킨 결과 화색이 빨간색으로 변경되었고(Nakatsuka et al., 2008a), F3’5’H와 anthocyanin 5,3’-aromatic acyltransferase(A5/3’AT) 유전자를 동시에 발 현 억제시켜 옅은 파란색 화색을 얻었다(Fig. 3B; Nakasuka et al., 2010). 토레니아에서 F3’5’H와 F3’H 유전자를 동시 에 발현억제시킨 결과 펠라고니딘계열의 안토시아닌 축적 이 유도되었다(Nakamura et al. 2010). 역방향 또는 RNAi 방법을 이용하여 플라보노이드 생합성을 조절하는 경우, 식 물 종이나 유전자에 따라서 발현억제 정도나 화색변경 정도 가 다르게 나타남으로(Nakamura et al., 2006, Nakatsuka et
al., 2008a), 목적하는 특정 식물에 적합한 유전자 발현 억제 기술을 선별하여 응용해야 한다.
유전자의 발현조절을 위해서 프로모터의 선발 또한 매우
중요하다. 상시 발현을 유도하는 cauliflower mosaic virus
35S(CaMV35S) 프로모터를 이용하여 장미, 카네이션, 토레
니아와 같은 화훼류들은 성공적으로 화색 변경을 이루어졌
다(Katsumoto et al., 2007; Nakamura et al., 2010; Ono et
al., 2006). 그러나, 용담과 국화 같은 몇몇 종의 작물에서
CaMV35S 프로모터는 도입유전자의 발현에 효과적이지 않
았다(Aida et al., 2004; Yamasaki et al., 2011). 용담의 경우
에는 CaMV35S 프로모터를 이용하여 형질 전환시킨 결과,
CaMV35S 프로모터 부위의 메틸화(methylation)가 관찰되
어 화색변경에는 효과적이지 않았고, 플라보노이드 생합성
구조유전자의 프로모터 또는 아그로박테리움의 rolC 프로모
터를 사용했을 때 화색변경이 관찰되었다(Nakatsuka et al.,
2008a). 국화의 경우도 CaMV35S 프로모터를 이용한 형질전
환보다는 국화의 ubiquitin extension 단백질의 프로모터 또는
chlorophyll a/b binding 단백질의 프로모터가 더 효과적으로
외래유전자를 발현시켰다(Aida et al., 2004; Annadana et al.,
2002).
Fig. 2. Strategies of flavonoid pathway engineering in flower. (A) Shematic overview of flavonoid pathway in flower. (B) Control of endogenous flavonoid pigments by gene suppression. (C) Novel flavonoid pigments production by introducing foreign genes.
(D) Upregulating anthocyanin biosynthesis using transcription factor gene. Upregulating genes by transcription factor are displayed in blue. An antisense and RNAi approach were used to block specific steps in the pathway (red crosses). Newly produced flavonoid branches are depicted in red. Painted colors show image of each compound.
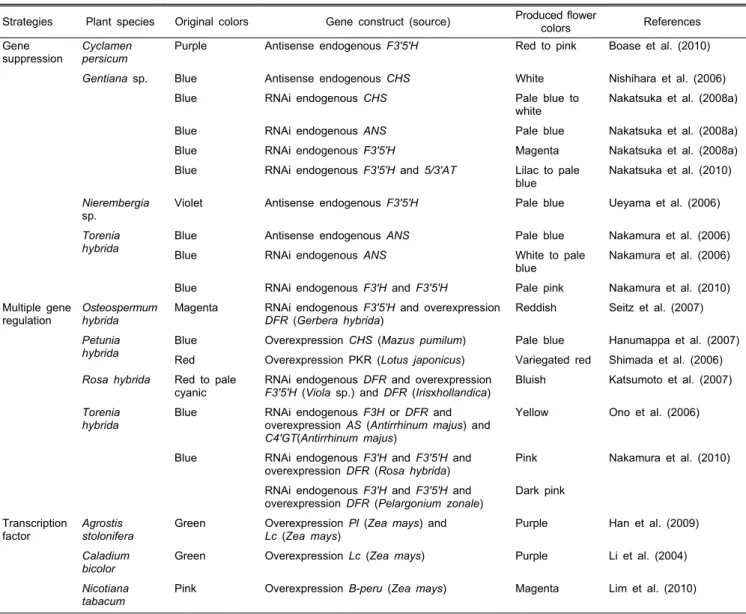
Table 2. Examples of flower color modifications by regulating flavonoid biosynthesis.
Strategies Plant species Original colors Gene construct (source) Produced flower
colors References
Gene
suppression Cyclamen
persicum Purple Antisense endogenous F3'5'H Red to pink Boase et al. (2010)
Gentiana sp. Blue Antisense endogenous CHS White Nishihara et al. (2006)
Blue RNAi endogenous CHS Pale blue to
white Nakatsuka et al. (2008a)
Blue RNAi endogenous ANS Pale blue Nakatsuka et al. (2008a)
Blue RNAi endogenous F3'5'H Magenta Nakatsuka et al. (2008a)
Blue RNAi endogenous F3'5'H and 5/3'AT Lilac to pale
blue Nakatsuka et al. (2010) Nierembergia
sp. Violet Antisense endogenous F3'5'H Pale blue Ueyama et al. (2006)
Torenia
hybrida Blue Antisense endogenous ANS Pale blue Nakamura et al. (2006)
Blue RNAi endogenous ANS White to pale
blue Nakamura et al. (2006) Blue RNAi endogenous F3'H and F3'5'H Pale pink Nakamura et al. (2010) Multiple gene
regulation Osteospermum
hybrida Magenta RNAi endogenous F3'5'H and overexpression
DFR (Gerbera hybrida) Reddish Seitz et al. (2007)
Petunia
hybrida Blue Overexpression CHS (Mazus pumilum) Pale blue Hanumappa et al. (2007) Red Overexpression PKR (Lotus japonicus) Variegated red Shimada et al. (2006) Rosa hybrida Red to pale
cyanic RNAi endogenous DFR and overexpression
F3'5'H (Viola sp.) and DFR (Irisxhollandica) Bluish Katsumoto et al. (2007) Torenia
hybrida Blue RNAi endogenous F3H or DFR and
overexpression AS (Antirrhinum majus) and C4'GT(Antirrhinum majus)
Yellow Ono et al. (2006)
Blue RNAi endogenous F3'H and F3'5'H and
overexpression DFR (Rosa hybrida) Pink Nakamura et al. (2010) RNAi endogenous F3'H and F3'5'H and
overexpression DFR (Pelargonium zonale) Dark pink Transcription
factor Agrostis
stolonifera Green Overexpression Pl (Zea mays) and
Lc (Zea mays) Purple Han et al. (2009)
Caladium
bicolor Green Overexpression Lc (Zea mays) Purple Li et al. (2004)
Nicotiana
tabacum Pink Overexpression B-peru (Zea mays) Magenta Lim et al. (2010)
따라서 다양한 GM 화훼작물을 개발하기 위해서는 다양 한 프로모터의 개발이 필요하며 다양한 프로모터의 활성 및 화색변경 정도 등을 고려하여 이용한다면 보다 정교한 화색 변경이 가능하리라 생각된다.
외래 유전자 추가 도입을 통한 화색 변경
꽃잎의 플라보노이드 성분은 종이나 품종에 따라 매우 다 양하다. 따라서 식물이 지니지 않는 색깔의 플라보노이드를 생산하기 위하여 내재유전자의 발현조절과 더불어 목적하 는 새로운 플라보노이드 합성과 관련된 외래유전자를 도입 해야 한다. 화훼작물에서 내재유전자의 조절과 더불어 외래 유전자를 도입하여 화색변경이 성공한 예들을 소개하고자 한다.
파란색 꽃 개발: 대부분의 파란색 꽃은 델피리딘계열 안 토시아닌 유래의 물질들이 축적됨으로 관찰된다. 델피리딘
계열 안토시아닌의 생산을 위해서는 F3’5’H 유전자의 도입
과 디하이드로미리세틴(dihydromyricetin)을 기질로 하는
DFR 유전자가 요구된다(Grotewold, 2006). 카네이션과 장미는파란색의 델피리딘계열 안토시아닌 색소 합성을 유도하는
F3’5’H 유전자의 결핍에 의해서 보라색 또는 파란색 계통의화색이 존재하지 않는다. 카네이션과 장미에서 플라보노이드
생합성 경로를 조절하여 델피리딘이 축적된 파란카네이션과
파란장미가 개발하였다. 카네이션의 경우 내재 DFR 유전자
가 제대로 작동하지 않은 하얀색 카네이션을 이용하여 페튜
니아의 F3’5’H 유전자와 디하이드로미리세틴을 기질로 선호하
는 페튜니아 DFR 유전자를 도입하여 파란카네이션이 개발되
었다. 파란장미의 경우에는 RNAi 방법을 이용하여 내재 DFR
발현을 억제하고 , 제비꽃의 F3’5’H와 붓꽃의 DFR 유전자를
도입하여 파란장미가 개발되었다(Fig. 3C; Katsumoto et al.,
2007). 팬지의 F3’5’H 유전자와 토네리아의 anthocyanin 5-
Fig. 3. Color modification by genetic engineering of the flavonoid biosynthetic pathway. (A) Flower color modification of cyclamen.
Left; host, right; transgenic line with co-suppressed CpF3’5’H gene (Boase et al., 2010). (B) Color modification of gentian. Left;
host, right; transgenic line co-suppressed Gt5’3’AT and GtF3’5’H gene shows the pale blue and white flower (Nakatsuka et al., 2010). (C) Color modification of Rosa hybrida. Left; host, middle and right: transgenic lines co-expressing VsF3’5’H and IhDFR, and knockdown of RhDFR RNAi accumulates blue delphinidin-based pigment (Katsumoto et al., 2007). (D) Torenia cv. Summerwave blue (left) accumulates delphinidin-based pigments, transgenic torenia (right) co-expressing AmAS1 and Am4’CGT, and knockdown of ThF3H RNAi accumulates yellow aurone-based pigments (Ono et al., 2006). (E) Purple tomato producing delphinidin was made from a red one producing cyanidin by co-expression of Antirrhinum majus Delia and Rosea gene (Butelli et al., 2008).
(F) Transgenic caladium expressing maize Lc gene. Left; host, right; transgenic line shows the red color in plant body (Li et al., 2005).
acyltransferase(5AT) 유전자만을 과발현시킨 파란장미도 개 발되었다.
붉은색 또는 주황색 꽃 개발: 붉은색 또는 주황색 꽃은 주로 시아니딘계열 또는 펠라고니딘계열 안토시아닌이 축 적됨으로 관찰된다. 작물에 따라서 지니고 있는 유전자의 기질특이성에 따라서 대사흐름이 일정한 방향으로만 이루 어진다. 따라서 작물의 내재 대사흐름을 원하는 대사방향으 로 조절하여 원하는 화색을 만들 수 있다. 페튜니아의 내재
DFR 유전자는 디하이드로캠페롤(dihydrokaempferol)을 기질로 이용하지 못하여서 주황색을 지닌 페튜니아가 자연계 에서 찾아보기가 어려웠다. 주황색 페튜니아를 개발하기 위 해서 디하이드로캠페롤을 특이적으로 이용하는 옥수수의
DFR 유전자를 페튜니아에 도입하여 꽃잎에서 주황색의 펠라고니딘이 생성되었다(Meyer et al., 1987). 국화과 다년초 인 오스테오스펄멈(osteospermum)의 경우 시아니딘계열 안
토시아닌을 축적함으로 붉은색을 나타내는데 , RNAi 방법 을 이용해서 내재 F3’H 유전자의 발현을 억제하고, 거베라 의 DFR 유전자를 도입하여 주황색의 펠라고니딘을 합성하 였다(Seitz et al., 2007). 담배의 경우에는 내재 FLS와 F3’H 유전자의 발현을 RNAi 방법을 이용하여 억제하고, 거베라 의 DFR 유전자를 과발현시킴으로 펠라고니딘의 생성을 유 도하였다(Nakatsuka et al., 2007b). 파란색 토레니아는 RNAi 방법을 이용해서 내재 F3’H 유전자와 F3’5’H 유전자의 발 현을 억제함과 동시에 제라늄 DFR 또는 장미 DFR 유전자 를 과발현시킴으로 분홍색 또는 짙은 분홍색을 나타내었다 (Nakamura et al., 2010).
노란색 꽃 개발: 금어초 꽃은 오론(aurone)이라고 불리는
색소 물질이 축적되어 노란색이 나타나는데, 오론 생합성에
관련한 유전자로 C4’GT와 AS 유전자가 동정되었다. 파란색
토레니아를 이용하여 오론 색소합성 관련 유전자인 금어초
C4’GT와 AS 유전자를 과발현시킴으로 노란색의 aureusidin
6-O-glucoside의 축적은 확인되었으나 내재 안토시아닌의 존재로 인해서 눈에 띄는 화색변화는 유도되지 못하였다.
내재 안토시아닌 색소 축적을 억제하기 위해서 내재 F3H와
DFR 유전자를 RNAi 방법으로 발현 억제하고, 동시에 금어초 유래의 C4’GT와 AS 유전자를 과발현시킴으로 노란색 화 색을 지니게 하였다(Fig. 3D; Ono et al., 2006). 코스모스나 다알리아 등은 polyketide reductase(PKR1)에 의해서 6’- deoxychalcone 물질이 축적되어 노란색이 나타난다. 노란 돌콩으로부터 PKR1 유전자를 분리하여 빨간색 페튜니아에 과발현시킨 결과 형질 전환된 페튜니아는 노란 색소물질인 isoliquritigenin 물질들은 축적되었으나, 얼룩덜룩한 빨간색 을 나타내었다(Shimada et al., 2006). 유전자 조작을 통해서 색소관련 물질의 생성은 확인되었으나 성공적인 노란색 꽃 의 개발을 위해서는 내재 안토시아닌 합성과 관련한 정교한 대사 흐름 조절이 요구된다. 이러한 유전자 조작기술을 응 용한다면 노란 화색을 지니지 않는 펠라고니늄, 베고니아, 시클라멘 등에서 노란색 화훼 작물 개발이 기대된다.
특정 플라보노이드 생산을 통한 화색 변경
색소가 없는 플라본과 플라보놀도 색소합성을 돕는 조색 소로 화색조절에 중요하며 조색소의 생성은 삼색단 이동 (bathchromic shift)과 안토시아닌의 안전성에 기여한다고 알려져 왔다. 특히 이러한 조색소의 생성은 파란색 꽃을 만 드는 데에 중요하다고 알려져 왔다(Yoshida et al., 2009).
플라본, 플라보놀, 안토시아닌은 공통의 전구체를 공유하여 색소물질을 합성하기 있기 때문에 조색소와 안토시아닌의 합성은 서로 영향을 준다. 플라본의 생합성을 조절하여 화색 을 변경된 예들을 살펴보면, 파란색 토레니아에서 DFR 유전 자를 역방향으로 발현시켰더니 플라본이 증가되어 보다 짙 은 파란색의 꽃이 생성되었고(Aida et al., 2000), flavone synthase II(FNSII) 유전자를 발현 억제하여서 옅은 파란색 의 안토시아닌이 축적된 토레니아를 얻었다(Ueyama et al., 2002). 담배에서 용담 FNSII 유전자를 과발현시킨 결과, 플 라본의 함량은 증가하였고 안토시아닌의 함량은 감소되었 다(Nakatsuka et al., 2006). 플라보놀 생합성 FLS 유전자를 이용하여, 보라색의 리시안셔스(lisianthus)의 내재 FLS 유 전자를 발현 억제한 결과, 디하이드로플라보놀이 축적되고 마젠타색의 꽃잎을 지니게 되었다(Nielsen et al., 2002).
다양한 수식관련 효소인 glycosyltransferase, betaine aldehyde dehydrogenase, serine carboxypeptidase-like acyltransferase 등은 다양한 기질물질을 당화, 가수화, 아실화를 시킬 수 있 어 잠정적으로 화색조절을 위한 목표 유전자가 되고 있다
(D’Auria et al., 2006; Gachon et al., 2005; Milkowski and Strack, 2004). 아직까지는 이러한 안토시아닌 수식관련 유 전자의 조절로 인해서 화색이 극적으로 변경된 보고는 없었다.
담배식물체에 용담 유래의 anthocyanin 5-O-glucosyltransferase (A5GT) 유전자를 과발현시킨 결과, 새로운 플라보노이드의 생성은 확인되었으나, 화색변화는 감지하기 어려운 정도였 다(Nakatsuka et al., 2008b). 주황색 또는 붉은색을 나타내 는 3-데옥시안토시아니딘의 생합성에 관한 유전자는 많이 알려져 있지 않았는데, 최근에 3-데옥시안토시아니딘에 특 이적인 수식화 효소인 UDP-glucose:3-deoxyanthocyanidin 5-O-glucosyltransferase(UGT5)를 Sinningia cardinalis에서 분리하였다(Nakatsuka and Nishihara, 2010). 이들 유전자를 이용한다면 붉은색과 주황색의 색소물질인 데옥시안토시아 니딘의 축적에 유용하리라 생각된다.
전사인자 조절을 통한 화색 변경
플라보노이드 생합성에 관여하는 구조유전자의 발현은 MYB, basic helix loop helix(bHLH), WD40 전사인자(MBW) 에 의해 조절되는 것으로 알려져 있다(Koes et al., 2005).
MBW 유전자군의 발현조절로 꽃과 과실에서의 안토시아닌 성분, 색소 축적뿐 아니라 다양한 생리에도 영향을 미치는 것 으로 보고되고 있다(Davies 2009; Nishihara and Nakatsuka, 2010). MYB 유전자가 돌연 변이된 흰색 페튜니아는 나방에 의해서 수분되지만 , 정상적인 MYB 유전자가 있는 붉은색 페 튜니아는 벌과 나비에 의해서 수분이 이루어졌다 (Quattrocchio et al., 1999).
플라보노이드 생합성관련 MBW 유전자군은 담배(Pattanaik
et al., 2010), 페튜니아(Quattrocchio et al., 2006), 사과(Espley
et al., 2007), 장미(Lin-Wang et al., 2010), 백합(Yamagishi
et al., 2010), 금어초(Schwinn et al., 2006), 나팔꽃(Morita
et al., 2006), 용담(Nakatsuka et al., 2008c) 등에서 보고되
었다. MBW 유전자군은 일반적으로 플라보노이드 생합성
경로에 관한 유전자 전체를 조절하기 때문에 MBW 유전자
군의 조절을 통한 플라보노이드 생합성 여러 유전자를 조절
하는 것이 가능하다 . 토마토 MYB 유전자를 과발현시킴으로
써 잎, 꽃, 과실과 전신에서 안토시아닌 색소의 축적을 유도
하였고(Mathews et al., 2003), 옥수수 전사인자인 Pl(MYB
전사인자)과 Lc(bHLH 전사인자)를 과발현시킴으로 보라색
의 잔디가 개발되었다(Han et al., 2009). 뿐만 아니라, 전사
인자에 따라 특정 구조유전자의 발현을 효과적으로 유도하
여 특정성분의 축적을 이끌어내기도 한다. 금어초의 Rosea
와 Delia 전사인자를 토마토에 과발현시킴으로 토마토의 과
피뿐 아니라 과육에서도 델피니딘계열 보라색 안토시아닌
축적이 확인되었다(Fig. 3E; Butelli et al., 2008). 최근에 MBW 유전자군 발현을 조절하는 R3 MYB(MybL2)전사인 자에 대한 연구가 진행되고 있다. 정상적인 생육조건하에서 는 MybL2는 MBW 유전자군의 bHLH에 결합하여 MBW 유전자군에 의한 안토시아닌 생합성을 억제하다가, 강광, 에 틸렌, 질소부족 등의 불량환경조건에 의해서 MybL2 발현이 억제되어 MBW 유전자군이 활성화되어 안토시아닌 생합성 을 유도하는 것으로 알려지고 있다(Dubos et al., 2008; Jeong et al., 2010; Matsui et al., 2008). 따라서 MBW 유전자군 및 MybL2의 발현조절을 통하여 안토시아닌의 축적 또는 감소 등을 유도하여 화색조절에 적용할 수 있으리라 생각된다.
유전자의 발현을 억제하는 방법으로 제시된 역방향방법 과 RNAi 방법 이외에도 최근에는 chimeric repressor gene- silencing technology(CRES-T)를 이용하는 방법이 개발되 었다. CRES-T 방법은 목적하는 전사인자와 강력한 유전자 발현억제 부위를 연결하여 식물에서 전사인자의 기능이 소 실됨에 따라 표현형을 관찰할 수 있으며, 역방향방법과 RNAi 방법과는 달리 목적하는 유전자의 염기서열이 밝혀 지지 않은 화훼작물에서도 적용 가능하다(Hiratsu et al., 2003). 토레니아, 나팔꽃, 국화, 리시안셔스, 용담, 시클라멘 등과 같은 다양한 원예작물에서 CRES-T 방법을 이용하여 키, 화형, 화색, 웅성불임과 호르몬 민감성과 관련한 다양한 표현형의 개발이 가능함을 보여주었고, 최근에는 CRES-T 방법을 이용하여 화훼작물에서 전사인자와 표현형에 관한 데이터베이스를 구축하였다(Mitsuda et al., 2008, 2011). 최 근 분자생물학 연구를 통하여 화색을 변경에 영향을 줄 수 있는 다양한 유전자들이 보고되고 있다. 세포내 pH가 화색의 색소 안정성에 영향을 준다고 알려져 있으며 튤립의 액포 이 온 수송체(Momonoi et al., 2009), 나팔꽃과 페튜니아의 액포 산성화 유전자(Verweij et al., 2008; Yoshida et al., 2009)와 더불어 금어초, 페튜니아, 꿩의다리의 세포형태(Baumann et al., 2007; Di Stilio et al., 2009) 관련된 유전자들도 화색에 영향을 미치는 것으로 밝혀졌다. 화색변경과 관련된 많은 연구들은 현재 활발히 진행 중이다.
결 론
화훼 작물에서 플라보노이드 생합성경로의 유전자조작을 통하여 이루어진 화색 변경의 성공적인 사례들을 보고하였 다. 플라보노이드 생합성관련 유전자의 분리 및 기능분석을 통해 플라보노이드 생합성 대사에 관한 깊이 있는 연구가 이루어짐에 따라 다양한 화훼류에서 원하는 화색변경이 가 능하게 되었다. 초기 연구에서는 플라보노이드 생합성 경로
의 단일유전자를 과발현 또는 발현억제함에 의해 화색 관련 대사물질의 합성을 늘이거나 억제하는 방법이 사용되었으 나, 기존에 없던 새로운 화색을 개발하기 위해서 점차 내재 유전자의 발현조절과 더불어서 새로운 플라보노이드 생합 성관련 외래유전자를 도입하는 등 여러 가지 생명공학 기법 을 병행하는 복잡한 방법을 통하여 화색변경을 이룬 경우도 있었다. 향후 좀 더 다양하고 독특한 화색 변경을 위해서는 대상 화훼 작물별로 내재하는 플라보노이드 경로의 조절 기 작과 대사물질의 흐름에 대한 이해가 요구되고, 이를 바탕 으로 목적하는 화색 물질 생산을 위해 정교한 대사공학 전 략이 요구된다.
일부 화색이 변경된 GM 화훼작물이 GM 식량작물과 마 찬가지로 이미 상업적으로 판매는 되고 있으나 매우 한정적 인 화훼 종에서만 상업화에 성공하였다. GM 작물의 상업화 는 연구, 개발 단계와 더불어서 더 많은 비용이 소요되는 안 전성평가 단계가 요구되는데, 이 때 개발국은 물론 상업화 하려는 해당국의 관련법을 준수하여 확실하게 수행되어야 한다(Tanaka et al., 2009). 특히 안전성평가 실험은 평가대 상 작물의 종류를 기반으로 이루어지기 때문에, 많은 수의 다양한 식물종류가 포함되어 있는 화훼작물의 경우에는 각 작물에 대한 개별적인 평가 기반이 갖추어져 있어야 하는 어려움이 있다. 이렇듯 비록 비식용 GM 작물인데도 불구하 고 안전성 평가에 소요되는 시간 및 비용이 GM 화훼 개발 과정에 투입된 시간과 비용보다 더 많이 요구되는 점을 감 안할 때, 유전자조작을 이용하여 새로운 화색을 지닌 화훼 작물을 개발하기 위해서는 화색변경을 위한 좀 더 체계적인 연구와 더불어서 GM 화훼 작물의 상업화를 위한 실증적 평 가 시스템의 창출 및 개선이 요구된다.
초 록
화색은 화훼 육종의 주요한 목표형질이다. 최근 유전공학
기술의 발달로 기존의 전통육종에서는 볼 수 없었던 파란장
미와 파란카네이션과 같은 새로운 화색 개발이 성공적으로
보고되었다. 플라보노이드 생합성에 관해 축적된 지식기반
연구 결과를 바탕으로 새롭고 독특한 형질의 화색을 도입하
는 것이 가능하게 된 것이다. 이러한 화색변경은 플라보노
이드 대사경로의 조절, 즉 내재유전자의 발현조절 및 새로
운 플라보노이드 합성 또는 특정 플라보노이드 합성을 위한
외래유전자의 추가도입과 플라보노이드 대사 전체를 조절
하는 전사인자의 도입을 통해서 이루어져 왔다. 그러나 보
다 실증적으로 이러한 플라보노이드 대사를 조절하기 위해
서는 작물별 내재 플라보노이드의 조절 기작에 대한 이해를
바탕으로 목표로 하는 플라보노이드 합성을 위해 보다 정교 한 대사흐름의 조절이 요구된다. 본 총설에서는 화훼작물의 화색변경 성공 예들을 자세히 소개하고 그 요인 분석을 통 해 향후 더 성공적인 화색변경의 전략을 수립하는데 도움이 되고자 한다.
추가 주요어 : 안토시아닌, 유전자변형, 대사공학, 전사인자
인용문헌
Aida, R., K. Yoshida, T. Kondo, S. Kishimoto, and M. Sibata.
2000. Copigmentation gives bluer flowers on transgenic torenia plants with the antisense dihydroflavonol-4-redutase gene. Plant Sci. 160:49-56.
Aida, R., K. Ohira, Y. Tanaka, K. Yoshida, S. Kishimoto, M.
Shibata, and A. Ohmiya. 2004. Efficient transgene expression in chrysanthemum, Dendranthema grandiflorum (Ramat.) Kitamura, by using the promoter of a gene for chrysanthemum chlorophyll-a/b-binding protein. Breeding Sci. 54:51-58.
Annadana, S., M.J. Beekwilder, G. Kuipers, P.B. Visser, N.
Outchkourov, A. Pereira, M. Udayakumar, J. De Jong, and M.A. Jongsma. 2002. Cloning of the chrysanthemum UEP1 promoter and comparative expression in florets and leaves of Dendranthema grandiflora. Transgenic Res. 11:437-445.
Baumann, K., M. Perez-Rodriguez, D. Bradley, J. Venail, P.
Bailey, H. Jin, R. Koes, K. Roberts, and C. Martin. 2007.
Control of cell and petal morphogenesis by R2R3 MYB transcription factors. Development 134:1691-1701.
Boase, M.R., D.H. Lewis, K.M. Davies, G.B. Marshall, D. Patel, K.E. Schwinn, and S.C. Deroles. 2010. Isolation and antisense suppression of flavonoid 3′,5′-hydroxylase modifies flower pigments and colour in cyclamen. BMC Plant Biol. 10:107.
Butelli, E., L. Titta, M. Giorgio, H.-P. Mock, A. Matros, S.
Peterek, E.G.W.M. Schijlen, R.D. Hall, A.G. Bovy, J. Luo, and C. Martin. 2008. Enrichment of tomato fruit with health- promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 26:1301-1308.
Chandler, S. and Y. Tanaka. 2007. Genetic modification in floriculture. Crit. Rev. Plant Sci. 26:169-197.
D'Auria, J.C. 2006. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 9:331-340.
Davies, K.M. 2009. Modifying anthocyanin production in flowers.
p. 49-83. In: K. Gould, K. Davies, and C. Winefield (eds.).
Anthocyanins biosynthesis functions, and applications. Springer, NY.
Di Stilio, V.S., C. Martin, A.F. Schulfer, and C.F. Connelly. 2009.
An ortholog of MIXTA-like2 controls epidermal cell shape in flowers of Thalictrum. New Phytol. 183:718-728.
Dubos, C., J. Le Gourrierec, A. Baudry, G. Huep, E. Lanet, I.
Debeaujon, J.M. Routaboul, A. Alboresi, B. Weisshaar, and L. Lepiniec. 2008. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 55:940-953.
Espley, R.V., R.P. Hellens, J. Putterill, D.E. Stevenson, S.
Kutty-Amma, and A.C. Allan. 2007. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 49:414-427.
Fukusaki, E., K. Kawasaki, S. Kajiyama, C.I. An, K. Suzuki, Y. Tanaka, and A. Kobayashi. 2004. Flower color modulations of Torenia hybrida by downregulation of chalcone synthase genes with RNA interference. J. Biotechnol. 111:229-240.
Gachon, C.M., M. Langlois-Meurinne, and P. Saindrenan. 2005.
Plant secondary metabolism glycosyltransferases: The emerging functional analysis. Trends Plant Sci. 10:542-549.
Grotewold, E. 2005. Plant metabolic diversity: A regulatory perspective. Trends Plant Sci. 10:57-62.
Grotewold, E. 2006. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 57:761-780.
Han, Y.J., Y.M. Kim, J.Y. Lee, S.J. Kim, K.C. Cho, T.
Chandrasekhar, P.S. Song, Y.M. Woo, and J.I. Kim. 2009.
Production of purple-colored creeping bentgrass using maize transcription factor genes Pl and Lc through Agrobacterium- mediated transformation. Plant Cell Rep. 28:397-406.
Hanumappa, M., G. Choi, S. Ryu, and G. Choi. 2007. Modulation of flower colour by rationally designed dominant-negative chalcone synthase. J. Exp. Bot. 58:2471-2478.
Hiratsu, K., K. Matsui, T. Koyama, and M. Ohme-Takagi. 2003.
Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis.
Plant J. 34:733-739.
Jeong, S.W., P. Kumar, S.C. Jeoung , J.Y. Song, H.Y. Lee, Y.K.
Kim, W.J. Kim, Y.I. Park, S.D. Yoo, S.B. Choi, G. Choi, and Y.I. Park. 2010. Ethylene suppression of sugar-induced anthocyanin pigmentation in Arabidopsis. Plant Physiol.
154:1514-1531.
Katsumoto, Y., M. Fukuchi-Mizutani, Y. Fukui, F. Brugliera, T.A.
Holton, M. Karan, N. Nakamura, K. Yonekura-Sakakibara, J. Togami, A. Pigeaire, G.Q. Tao, N.S. Nehra, C.Y. Lu, B.K.
Dyson, S. Tsuda, T. Ashikari, T. Kusumi, J.G. Mason, and Y. Tanaka. 2007. Engineering of the rose flavonoid biosynthetic pathway successfully generated blue-hued flowers accumulating delphinidin. Plant Cell Physiol. 48:1589-1600.
Koes, R., W. Verweij, and F. Quattrocchio. 2005. Flavonoids:
A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 10:236-242.
Li, S.J., X.M. Deng, H.Z. Mao, and Y. Hong. 2005. Enhanced anthocyanin synthesis in foliage plant Caladium bicolor. Plant Cell Rep. 23:716-720.
Lim, S.-H., S.H. Ha, J.K. Kim, S.H. Sohn, D.H. Kim, J.S. Kim, Y.M. Kim, and J.R. Lee. 2010. Expression of phenylpropanoid biosynthetic genes in transgenic tobacco plants with maize B-peru Gene. Kor. J. Intl. Agri. 22:72-77.
Lin-Wang, K., K. Bolitho, K. Grafton, A. Kortstee, S. Karunairetnam, T.K. McGhie, R.V. Espley, R.P. Hellens, and A.C. Allan.
2010. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae.
BMC Plant Biol. 10:50.
Mathews, H., S.K. Clendennen, C.G. Caldwell, X.L. Liu, K.
Connors, N. Matheis, D.K. Schuster, D.J. Menasco, W.
Wagoner, J. Lightner, and D.R. Wagner. 2003. Activation tagging in tomato identifies a transcriptional regulator of
anthocyanin biosynthesis, modification, and transport. Plant Cell 15:1689-1703.
Matsui, K., Y. Umemura, and M. Ohme-Takagi. 2008. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 55:954-967.
Meyer, P., I. Heidmann, G. Forkmann, and H. Saedler. 1987.
A new petunia flower colour generated by transformation of a mutant with a maize gene. Nature 330:677-678.
Milkowski, C. and D. Strack. 2004. Serine carboxypeptidase-like acyltransferases. Phytochemistry 65:517-524.
Mitsuda N., Y. Takiguchi, M. Shikata, K. Sage-Ono, M. Ono, K. Sasaki, H. Yamaguchi, T. Narumi, Y. Tanaka, T. Terakawa, K. Gion, R. Suzuri, Y. Tanaka, T. Nakatsuka, S. Kimura, M.
Nishihara, T. Sakai, R. Endo-Onodera, K. Saitoh, K. Isuzugawa, Y. Oshima, T. Koyama, M. Ikeda, M. Narukawa, K. Matsui, M. Nakata, N. Ohtsubo, and M. Ohme-Takagi. 2011. The new FioreDB database provides comprehensive information on plant transcription factors and phenotypes induced by CRES-T in ornamental and model plants. Plant Biotechnol. 28:123-130.
Mitsuda, N., Y. Umemura, M. Ikeda, M. Shikata, T. Koyama, K. Matsui, T. Narumi, R. Aida, K. Sasaki, T. Hiyama, Y.
Higuchi, M. Ono, K. Isuzugawa, K. Saitoh, R. Endo, K. Ikeda, T. Nakatsuka, M. Nishihara, S. Yamamura, T. Yamamura, T.
Terakawa, N. Ohtsubo, and M. Ohme-Takagi. 2008. FioreDB:
A database of phenotypic information induced by the chimeric repressor silencing technology (CRES-T) in Arabidopsis and floricultural plants. Plant Biotechnol. 25:37-44.
Momonoi, K., K. Yoshida, S. Mano, H. Takahashi, C. Nakamori, K. Shoji, A. Nitta, and M. Nishimura. 2009. A vacuolar iron transporter in tulip, TgVit1, is responsible for blue coloration in petal cells through iron accumulation. Plant J. 59:437-447.
Morita, Y., M. Saitoh, A. Hoshino, E. Nitasaka, and S. Iida. 2006.
Isolation of cDNAs for R2R3-MYB, bHLH and WDR transcriptional regulators and identification of c and ca mutations conferring white flowers in the Japanese morning glory. Plant Cell Physiol. 47:457-470.
Nakamura, N., M. Fukuchi-Mizutani, K. Miyazaki, K. Suzuki, and Y. Tanaka. 2006. RNAi suppression of the anthocyanidin synthase gene in Torenia hybrida yields white flowers with higher frequency and better stability than antisense and sense suppression. Plant Biotechnol. 23:13-18.
Nakamura, N., M. Fukuchi-Mizutani, Y. Fukui, K. Ishiguro, K.
Suzuki, H. Suzuki, K. Okazaki, D. Shibata, and Y. Tanaka.
2010. Generation of pink flower varieties from blue Torenia hybrida by redirecting the flavonoid biosynthetic pathway from delphinidin to pelargonidin. Plant Biotechnol. 27:375-383.
Nakatsuka, T. and M. Nishihara. 2010. UDP-glucose:3- deoxyanthocyanidin 5-O-glucosyltransferase from Sinningia cardinalis. Planta 232:383-392.
Nakatsuka, T., C. Pitaksutheepong, S. Yamamura, and M.
Nishihara. 2007a. Induction of differential flower pigmentation patterns by RNAi using promoters with distinct tissue-specific activity. Plant Biotech. Rep. 1:251-257.
Nakatsuka, T., K. Mishiba, A. Kubota, Y. Abe, S. Yamamura, N. Nakamura, Y. Tanaka, and M. Nishihara. 2010. Genetic engineering of novel flower colour by suppression of anthocyanin
modification genes in gentian. J. Plant Physiol. 167:231-237.
Nakatsuka, T., K. Mishiba, Y. Abe, A. Kubota, Y. Kakizaki, S.
Yamamura, and M. Nishihara. 2008a. Flower color modification of gentian plants by RNAi-mediated gene silencing. Plant Biotechnol. 25:61-68.
Nakatsuka, T., K. Sato, H. Takahashi, S. Yamamura, M. Nishihara.
2008b. Cloning and characterization of the UDP-glucose:
anthocyanin 5-O-glucosyltransferase gene from blue-flowered gentian. J. Exp. Bot. 59:1241-1252.
Nakatsuka, T., K.S. Haruta, C. Pitaksutheepong, Y. Abe, Y.
Kakizaki, K. Yamamoto, N. Shimada, S. Yamamura, and M.
Nishihara. 2008c. Identification and characterization of R2R3- MYB and bHLH transcription factors regulating anthocyanin biosynthesis in gentian flowers. Plant Cell Physiol. 49:1818-1829.
Nakatsuka, T., M. Nishihara, K. Mishiba, and S. Yamamura. 2006.
Heterologous expression of two gentian cytochrome P450 genes can modulate the intensity of flower pigmentation in transgenic tobacco plants. Mol. Breeding 17:91-99.
Nakatsuka, T., Y. Abe, Y. Kakizaki, S. Yamamura, and M.
Nishihara. 2007b. Production of red-flowered plants by genetic engineering of multiple flavonoid biosynthetic genes. Plant Cell Rep. 26:1951-1959.
Nakayama, T., K. Yonekura-Sakakibara, T. Sato, S. Kikuchi, Y.
Fukui, M. Fukuchi-Mizutani, T. Ueda, M. Nakao, Y. Tanaka, T. Kusumi, and T. Nishino. 2000. Aureusidin synthase: A polyphenol oxidase homolog responsible for flower coloration.
Science 290:1163-1166.
Nielsen, K., S.C. Deroles, K.R. Markham, J.M. Bradley, E.
Podivinsky, and D. Manson. 2002. Antisense flavonol synthase alters copigmentation and flower color in lisianthus. Mol.
Breed. 9:217-229.
Nishihara, M. and T. Nakatsuka. 2010. Genetic engineering of novel flower colors in floricultural plants: Recent advances via transgenic approaches. Methods Mol. Biol. 589:325-347.
Nishihara, M., T. Nakatsuka, and S. Yamamura. 2005. Flavonoid components and flower color change in transgenic tobacco plants by suppression of chalcone isomerase gene. FEBS Lett.
579:6074-6078.
Nishihara, M., T. Nakatsuka, K. Hosokawa, T. Yokoi, Y. Abe, K. Mishiba, and S. Yamamura. 2006. Dominant inheritance of white-flowered and herbicide-resistant traits in trangenic gentian plants. Plant Biotechnol. 23:25-31.
Ono, E., M. Fukuchi-Mizutani, N. Nakamura, Y. Fukui, K.
Yonekura-Sakakibara, M. Yamaguchi, T. Nakayama, T.
Tanaka, T. Kusumi, and Y. Tanaka. 2006. Yellow flowers generated by expression of the aurone biosynthetic pathway.
Proc. Nat. Acad. Sci. USA 103:11075-11080.
Pattanaik, S., Q. Kong, D. Zaitlin, J.R. Werkman, C.H. Xie, B.
Patra, and L. Yuan. 2010. Isolation and functional characterization of a floral tissue-specific R2R3 MYB regulator from tobacco.
Planta 231:1061-1076.
Quattrocchio, F., A. Baudry, L. Lepiniec, and E. Grotewold. 2006.
The regulation of flavonoid biosynthesis. p. 97-122. In: E.
Grotewold (ed.). The science of flavonoids. Springer, NY.
Quattrocchio, F., J. Wing, K. van der Woude, E. Souer, N. de Vetten, J. Mol, and R. Koes. 1999. Molecular analysis of the anthocyanin2 gene of petunia and its role in the evolution
of flower color. Plant Cell 11:1433-1444.
Schwinn, K., J. Venail, Y. Shang, S. Mackay, V. Alm, E. Butelli, R. Oyama, P. Bailey, K. Davies, and C. Martin. 2006. A small family of MYB-regulatory genes controls floral pigmentation intensity and patterning in the genus Antirrhinum. Plant Cell 18:831-851.
Seitz, C., M. Vitten, P. Steinbach, S. Hartl, J. Hirsche, W. Rathje, D. Treutter, and G. Forkmann. 2007. Redirection of anthocyanin synthesis in Osteospermum hybrida by a two-enzyme manipulation strategy. Phytochemistry 68:824-833.
Shimada, N., T. Nakatsuka, M. Nishihara, S. Yamamura, S.
Ayabe, and T. Aoki. 2006. Isolation and characterization of a cDNA encoding polyketide reductase in Lotus japonicus.
Plant Biotechnol. 23:509-513.
Tanaka, Y., F. Brugliera, and S. Chandler. 2009. Recent progress of flower colour modification by biotechnology. Int. J. Mol.
Sci. 10:5350-5369.
Tanaka, Y., Y. Katsumoto, F. Brugliera, and J. Mason. 2005.
Genetic engineering in floriculture. Plant Cell Tiss. Org. Cult.
80:1-24.
Ueyama, Y., K. Suzuki, M. Fukuchi-Mizutani, Y. Fukui, K.
Miyazaki, H. Ohkawa, T. Kusumi, and Y. Tanaka. 2002.
Molecular and biochemical characterization of torenia flavonoid 3′-hydroxylase and flavone synthase II and modification of flower color by modulating the expression of these genes. Plant Sci. 163:253-263.
Ueyama, Y., Y. Katsumoto, Y. Fukui, M. Fukuchi-Mizutani, H.
Ohkawa, T. Kusumi, T. Iwashita, and Y. Tanaka. 2006.
Molecular characterization of the flavonoid biosynthetic pathway and flower color modification of Nierembergia sp.
Plant Biotechnol. 23:19-24.
Umehara, M., A. Hanada, S. Yoshida, K. Akiyama, T. Arite, N.
Takeda-Kamiya, H. Magome, Y. Kamiya, K. Shirasu, K.
Yoneyama, J. Kyozuka, and S. Yamaguchi. 2008. Inhibition of shoot branching by new terpenoid plant hormones. Nature 455:195-200.
van der Krol, A.R., P.E. Lenting, J. Veenstra, I.M. van der Meer, R.E. Koes, A.G.M. Gerats, J.N.M. Mol, and A.R. Stuitje. 1988.
An anti-sense chalcone synthase gene in transgenic plants inhibits flower pigmentation. Nature 333:866-869.
Verweij, W., C. Spelt, G.P. Di Sansebastiano, J. Vermeer, L.
Reale, F. Ferranti, R. Koes, and F. Quattrocchio. 2008. An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour. Nat. Cell Biol. 10:1456-1462.
Winefield, C., H. David, E. Swinny, H. Zhang, H. Arathoon, T.
Fischer, H. Halbwirth, K. Stich, C. Gosch, G. Forkmann, and K. Davies. 2005. Investigation of the biosynthesis of 3-deoxyanthocyanins in Sinningia cardinalis. Physiologia Plantarum 124:419-430.
Yamagishi, M., Y. Shimoyamada, T. Nakatsuka, and K. Masuda.
2010. Two R2R3-MYB genes, homologs of petunia AN2, regulate anthocyanin biosyntheses in flower tepals, tepal spots and leaves of Asiatic hybrid lily. Plant Cell Physiol. 51:463-474.
Yamasaki, M., M. Oda, N. Koizumi, K. Mitsukuri, M. Johkan, T. Nakatsuka, M. Nishihara, and K. Mishiba. 2011. De novo DNA methylation of the 35S enhancer revealed by high- resolution methylation analysis of an entire T-DNA segment in transgenic gentian. Plant Biotechnol. 28:223-230.
Yoshida, K., M. Mori, and T. Kondo. 2009. Blue flower color development by anthocyanins: From chemical structure to cell physiology. Nat. Prod. Rep. 26:884-915.