산 및 효소 가수분해를 이용한 홍조류로부터 바이오 에탄올 생산
최수정⋅이성목⋅이재화
†
신라대학교 의생명과학대 생명공학과(2012년 2월 6일 접수, 2012년 2월 24일 심사, 2012년 3월 11일 채택)
-
Production of Bio-ethanol from Red Algae by Acid Hydrolysis and Enzyme Treatment
Soo-Jeong Choi, Sung-Mok Lee, and Jae-Hwa Lee
†
Department of Bioscience and Biotechnology, College of Medical and Life Science, Silla University, Busan 617-736, Korea (Received February 6, 2012; Revised February 24, 2012; Accepted March 11, 2012)
화석연료로 인한 환경오염 등의 문제를 해결하기 위해서 다양한 원료를 이용하여 바이오 에탄올 생산에 대한 연구가 진행되고 있다. 해조류 중에 홍조류는 agar, carrageenan, porphyran으로 구성되어 있어 산 처리를 통해 바이오에탄올 생산에 유용한 바이오매스로 전환이 가능하다. 본 연구는 홍조류의 가수분해물을 이용하여 바이오에탄올 생산의 최적 조건을 찾으려고 한다. 바이오에탄올 생산하기 위해 전처리 된 우뭇가사리에 Saccharomyces cerevisiae KCCM112를 접 종해 발효하였다. 우뭇가사리 가수분해의 최적조건은 1.5% H 2 SO 4 를 121 ℃에서 30 min 반응시켰을 때 7.04 g/L의 gal- actose 와 1.94 g/L의 glucose가 생산되었다. 그리고 CH 3 COOH 의 경우 2.0% 농도로 처리하였을 때, galactose 0.75 g/L가 생산되었다. 이와 반대로 도박에서는 H 2 SO 4 1.5%를 처리하였을 때 galactose를 6.38 g/L 생산하였으며, CH 3 COOH을 처리했을 때 0.368 g/L이 생산되었다. 우뭇가사리에서 에탄올 생산은 1.0% H 2 SO 4 를 121 ℃에서 30 min 간 처리하였을 때 가장 높았으며, 96 h 배양하였을 때 3.77 g/L의 에탄올을 생산했다.
Bio-ethanol production research using various material has been problemed for solving problems of environment pollution caused by fossil fuels. Red-algae consists of agar, carrageenan, and porphyran. If it is treated by acid, it is able to change useful bio-mass for bio-ethanol. In this study, we found an optimal condition for bio‐ethanol production from acid hydrolysate in red-algae. To produce bio-ethanol, Saccharomyces cerevisiae KCCM1129 inoculated to acid hydrolysate of Gelidium amansii. The optimal condition for Gelidium amansii hydrolysis was found to be 30 min reaction at H 2 SO 4 concentration of 1.5% and 121 ℃. At this condition, its produced to 7.04 g/L galactose and 1.94 g/L glucose. And acetic acid concentration of 2.0% in agar produced 0.75 g/L galactose. In contrast, Pachymeniopis elliptica was treated with H 2 SO 4 concentration of 1.5%, it produced 6.38 g/L galactose. And Pachymeniopis elliptica treated with acetic acid concentration of 2% produced 0.368 g/L galactose. The optimal condition of ethanol production was found to be 96 h reaction at H 2 SO 4 concentration of 1.0% and 30 ℃, which produced 3.77 g/L ethanol.
Keywords: bio-ethanol, pretreatment, Saccharomyces cerevisiae, Gelidium amansii
1. 서 론
1)
오늘날 인류는 인구증가와 산업화에 의한 화석연료의 고갈에 대한 문제를 가지는데, 특히 화석연료 사용으로 겪어야 하는 지구온난화에 의한 문제와 국제유가의 상승 문제는 심각하다. 이러한 문제로 인해 신 재생 에너지 개발과 함께 에너지 절감의 필요성을 느낀 세계 각국 은 이산화탄소 배출량 감소를 위해서 화석연료를 대체할 안정적인 에 너지 자원을 탐색 중에 있으며 그 중 환경오염 정도가 낮으며, 탄소효 율이 높고 휘발유와 혼합 사용이 가능한 바이오 에너지인 에탄올 생 산에 대해 관심을 가지고 있다. 에탄올을 생산하기 위한 대표적인 기 질로는 크게 당질계, 전분계, 목질계로 나눌 수 있다. 식량으로 쓰는
† 교신저자 (e-mail: [email protected])
당질계 작물인 사탕수수와 당밀 그리고 전분계 작물인 옥수수와 같은
곡류, 고구마 등은 인류의 식량 문제와 연관을 가지면서 작물이 풍부
한 국가에서만 에탄올 연구가 이루어진다는 단점을 가지고 있다. 목
질계인 폐 목재와 농업 부산물의 경우 에탄올을 생산하는 수율은 높
지만, 자원이 한정되어 있고 열대 우림을 파괴해 자연을 훼손한다는
점에서 문제가 되고 있다[1,2]. 이러한 단점을 극복하기 위해 해조류
를 이용한 바이오 에너지 생산이 주목 받고 있다. 해조류는 광합성을
통해서 이산화탄소의 처리가 가능한데, 이때 소비되는 이산화탄소는
지구 온난화 정도를 낮추는데 크게 기여할 것으로 생각된다. 또한 해
조류는 수중의 유기물을 흡수하며 부영양화를 감소시키는 기능을 하
므로 환경 정화에도 유용하게 이용될 것으로 보인다[3-5]. 크게 해조
류 바이오매스는 녹조류와 갈조류, 홍조류로 나눌 수 있다. 그 중 홍
조류는 물이나 다른 비료가 필요 없을 정도로 성장이 매우 우수하며,
에너지로 전환되는 정도가 높아 목질계 수준의 바이오 에탄올 생산도
가능하기 때문에 새로운 바이오매스 자원으로의 개발 가능성이 높을 것으로 예상된다[3].
홍조류는 주요 당 성분으로 agar, carrageenan, porphyran 등으로 구성 되어 있으며, 산지와 채취한 시기, 종류에 따라서 성분 함량이 다르게 나타난다. Agar는 홍조류를 구성하는 다당류 중 하나로 세포벽을 이루는 물질이다. Agar의 구조는 α(1-4)-3,6-anhydro-D-galactose와 β(1-3)-D-galactose가 교차 결합한 상태로 약 6%의 황산기가 ester 결합을 가지고 있다[7]. Carrageenan은 점질성 다당류로, 분자량은 10
∼80만 정도이다. 기본구조는 D-galactose와 3, 6-anhydro-D-galactose 가 α-1, 3 및 β-1, 4 결합으로 황산기의 위치에 따라서 여러 종류로 나눌 수 있다[8,9]. Porphyran은 세포벽이나 세포 사이 충진 물질로 존재하는 수용성 산성 다당류로 agarose와 유사하다. 또한 황산기와 methoxy 가 다량 함유되어 있는 물질로 3, 6-anhydro-L-galactose, 6-0- methyl-D-galactose, D, L-galactose 및 ester sulfate가 구성성분이다.
홍조류에 포함되어 있는 다당류들은 효소나 물리적, 화학적 가수분 해를 통한 단당류로 전환이 가능하다. 홍조류 중 agar와 carrageenan을 많이 함유하고 있는 우뭇가사리(Gelidium amansii)와 도박(Pachymeniopis elliptica) 을 이용해 실험을 진행하였으며, 전처리 과정을 통해 가수분 해하고 이 중 환원당 생성량이 높은 우뭇가사리를 이용하여 에탄올 발효과정을 통해 에탄올 생산 가능성을 확인하였다.
2. 실 험
2.1. 균주 및 배지
에탄올 발효를 위한 균주로 효모인 Saccharomyces cerevisiae KCCM- 1129 를 사용하였고, 균주는 -70 ℃에서 20% glycerol로 stock해서 보관 하였다. Stock한 균주는 YPD배지(glucose 20.0 g/L, peptone 20.0 g/L, yeast extract 10.0 g/L)를 30 ℃, 150 rpm에서 진탕 배양 시킨 후 이용 하였다[12]. 에탄올 발효에 사용한 우뭇가사리는 부전시장에서 구입 하였으며, 도박은 국립수산과학원을 통해 입수하였다. 홍조류는 실온 에서 완전히 건조 후 ball mill을 이용하여 분쇄하였다. 에탄올 발효를 위한 우뭇가사리와 도박 배지는 우뭇가사리와 도박만을 20 g/L 농도로 첨가하여 사용하였다. 에탄올 발효는 300 mL flask에서 working vol- ume 을 100 mL로 하였으며, 전 배양한 효모 3 mL을 접종하여 30 ℃, 150 rpm 에서 진탕 배양하였다.
2.2. 산 종류 및 농도에 따른 홍조류 가수분해
산 처리 후 홍조류에서 일어나는 가수분해 정도를 확인하기 위해서 H 2 SO 4 , CH 3 COOH 를 0.1%에서 2.0%의 농도로 처리하였다. 산 농도 별 배지 80 mL 당 우뭇가사리와 도박을 각각 2 g씩 넣고 고압멸균기 로 121 ℃에서 30 min 동안 처리하였다. 멸균이 끝난 후에는 NaOH로 중화를 하였고 최종 working volume을 100 mL로 조절하여(pH 6.8∼
7.2) 진탕배양기의 30 ℃, 150 rpm 환경에서 발효하였다. 효소 전처리 실험도 진행하였는데, 사용하는 효소는 celluclast로 홍조류 내 cellu- lose 를 분해하여 galactose나 glucose 같은 당을 생산하기 위해 이용하 였다. 전처리 환경에 따른 당의 가수분해 정도를 파악하기 위해서 HPLC를 이용해 측정하였다. 가수분해된 당 성분 및 함량 측정을 위 해 시료 1.0 mL을 syringe filter로 여과 후 HPLC를 이용하여 확인했 으며 정량적 분석을 위해 다양한 농도의 galactose와 glucose를 표준시 약으로 이용하였다.
2.3. 에탄올 함량 측정
산 처리가 되어 있는 우뭇가사리에서 생성된 에탄올을 측정하기 위 해서 발효된 시료를 14000 rpm에서 5 min 동안 원심분리 후 얻은 상 층액을 gas chromatography (GC)를 이용하여 분석하였다. 분석을 위 해 사용한 GC는 HP5890 series II로, 칼럼은 HP-FFAP (Cross-Linked PEG-TPA 30 m / 0.25 mm / 0.25 µL)를 사용하였다. 이동상으로는 N 2 를 0.6 mL/min으로 이용하였으며, injection temperature 는 150 ℃, detector temperature 200 ℃, 승온조건은 45 ℃ (2 min) / (1 ℃/min) / 50 ℃ (1 min) / (20 ℃ /min) / 90 ℃ (1 min) / (30 ℃/min) / 150 ℃ (1 min) 으로 설정하였다. 분리비는 70 : 1로 하였고 내부 표준물질은 1% 의 isopropanol을 이용했다[8].
2.4. 당 함량 측정
생성된 당의 함량 측정을 위해서 산과 효소로 전처리 한 시료를 HPLC를 이용하였다. 즉 시료 1.0 mL을 0.22 µm millipore filter를 이 용하여 불순물을 제거한 후, HPLC용 vial에 넣어 측정을 하였다. 실험 에 이용한 HPLC의 detector는 RID-10A를 쓰고 column은 Bio-Rad의 Aminex HPX-87H 300 mm × 7.8 mm 을 사용하였으며 oven 온도는 40 ℃로 설정하였다. HPLC의 유속은 0.6 mL/min이고, 용매는 HPLC 용 water를 이용하였다[9,10]. 표준 검량선은 우뭇가사리에서 분해되 어 나올 수 있는 glucose와 galactose를 1 L당 2.5, 5, 10 g으로 만들어 표준시약으로 사용하였다.
에탄올 발효과정에서 환원당의 감소량 확인에는 DNS법을 이용하 였다. DNS법은 시료를 14000 rpm, 5 min 동안 원심분리 후에 얻어진 상층 액 500 µL를 DNS용액 2.0 mL와 혼합시킨 후 시료를 10 min 동안 가열하고 나서 UV spectrometer를 이용하여 540 nm에서 측정하 였다. 표준 검량선의 경우 galactose를 이용해 생성된 환원당을 정량하 였다. 환원당 전환율은 (생성된 환원당 / 전처리에 사용된 배지의 시 료) × 100으로 계산하였다[12].
3. 결 과
3.1. 홍조류의 산 전처리
Agar 와 carrageenan이 많이 포함되어 있는 홍조류 가운데 우뭇가사 리와 도박을 이용하여 산 가수분해 실험을 진행하였다. 산은 강산인 H 2 SO 4 와 유기산인 CH 3 COOH 을 이용하였으며, 전처리 농도는 0.1%
에서 2.0%로 조절하였다. 각각의 산을 첨가한 시료는 121 ℃, 30 min 동안 고압멸균 과정을 거쳤다. 이후 배지의 pH를 7.0으로 맞추어 조절 한 뒤, 배지에 부유물이 생기는 것을 방지하기 위해서 30 ℃, 150 rpm 으로 24 h 동안 교반하였다. 시료들에게서 생긴 당 성분은 HPLC를 이용해 분석하였다.
우뭇가사리에 산으로 전처리할 경우 agar가 분해되어 galactose가 생산되는 것을 HPLC 분석 결과로 확인 할 수 있었다(Figure 1)[9,10].
H 2 SO 4 이 CH 3 COOH 에 비해서 많은 galactose가 생성되는 결과도 얻을 수 있었다. 우뭇가사리와 도박에 H 2 SO 4 과 CH 3 COOH 을 처리한 결과 를 비교해보면 H 2 SO 4 1.5%를 처리한 우뭇가사리 배지에서는 7.04 g/L 의 galactose가 생산되어 투입한 우뭇가사리의 35%가 환원당으로 전 환되었으며, CH 3 COOH 2.0% 를 처리한 배지는 H 2 SO 4 에 비하면 약 10% 에 해당되는 0.7 g/L의 galactose를 생산하는 것으로 확인되었다.
도박의 경우 H 2 SO 4 1.5% 를 처리하면 6.38 g/L의 galactose가 생산되어 31.9% 의 수율을 보였으며, CH 3 COOH 2.0% 를 처리한 배지는 H 2 SO 4
에 비해 훨씬 낮은 0.36 g/L의 galactose를 생산하였다. 도박과 우뭇가
(a)
(b)
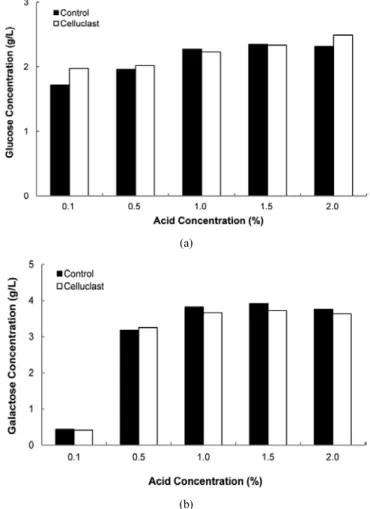
Figure 2. Hydrolysis of Gelidium amansii by enzyme and acid treatment : The hydrolysis enzyme (celluclast) wasused cellulose hydrolysis in the experiment. Enzyme and acid hydrolysate was produced fermentable sugar such as glucose and galactose. (a) Glucose, (b) Galactose.
(a)
(b)
Figure 1. Hydrolysis of red algae using organic acid : The red-algae were treated with H 2 SO 4 , CH 3 COOH in the various concentrations.
The galactose was produced from red algae hydrolysate. (a) Gelidium amansii, (b) Pachymeniopis elliptica.
사리의 환원당 생성량을 측정한 결과 동일한 환경임에도 불구하고 우뭇가사리에서 도박보다 많은 galactose가 생산되었다. 처리한 산의 종류에 따라 galactose의 생산수율이 달랐는데 이는 유기산에 해당되는 CH 3 COOH 이 강산인 H 2 SO 4 보다 산성이 약하기 때문인 것으로 보인 다. 실험을 통해 홍조류 성분인 agar 또는 carageenan을 단당류로 산을 이용하여 가수분해 할 때 전처리에는 H 2 SO 4 이 적합하다는 것을 알 수 있었고, 우뭇가사리에서 도박에 비해 작게는 10%에서부터 많게는 2배 이상의 더 많은 단당류가 생성되는 것을 확인할 수 있었다. 따라서 기질인 우뭇가사리에 최적의 전처리 조건을 적용하여 효소 가수분해 및 에탄올 발효 실험을 진행하였다.
3.2. 홍조류의 효소 전처리
우뭇가사리와 같은 홍조류는 주요 다당류로 agar를 포함하고 있으며 5 ∼10% 정도의 cellulose도 포함하고 있다. 따라서 우뭇가사리에 포함 된 cellulose를 효과적으로 이용하기 위해 산으로 전처리된 우뭇가사 리에 celluclast를 첨가하여 시간에 따른 당 성분의 증가를 확인하였다.
30 min 간격으로 시료를 추출하여 홍조류 가수분해 과정에서 생성될 수 있는 glucose와 galactose 성분에 대한 확인실험을 진행하였다. 실 험에 사용된 산의 종류는 H 2 SO 4 으로 0.1%에서 2.0%까지 농도별로 처 리하였다. 우뭇가사리에 cellulose 가수분해 효소인 celluclast를 첨가
해 가수분해를 하고 HPLC를 이용하여 생성된 당 성분을 분석한 결과 galactose 와 glucose 모두 확인할 수 있었다. 반응 초기와 비교하였을 때 glucose와 galactose 모두 2 h 반응시켰을 때 높은 생성률을 보였다.
Glucose 의 경우 처리한 산의 농도가 2.0% 일 때, 반응 초기에서는 2.31 g/L, 2 h 반응에서는 2.49 g/L를 생산하였다. 반응시간에 따른 glucose 의 생산량을 비교해 본 결과 celluclast의 최적 반응 시간은 2 h 으로 생각된다. 그러나 galactose는 glucose와 다른 생산 양상을 보인 다. 효소 첨가 후 반응 2 h일 때, glucose 의 농도가 증가하는 반면 gal- actose는 일부 감소하는 것으로 나타났다. Galactose를 최대로 얻을 수 있는 산의 농도는 1.5%로, 최대 3.91 g/L의 galactose를 얻을 수 있었 다. 그러나 효소처리 후에는 galactose 일부가 감소한 3.72 g/L가 생산 되었다. 따라서 이것은 산 처리만 진행한 홍조류에서 보다 낮은 수치 로 투입한 우뭇가사리의 약 18.6%가 환원당으로 전환된 것으로 나타 남을 알 수 있다(Figure 2).
3.3. 홍조류 가수분해 산물을 이용한 에탄올 발효
에탄올 발효를 위해서 YPD 배지에서 전 배양한 Saccharomyces
cerevisiae KCCM1129를 이용하였다. 우뭇가사리와 도박이 첨가되어
있는 배지에 이전 조건과 마찬가지로 H 2 SO 4 을 0.1%에서 2.0%까지
처리하였다. 전처리에 따라 세가지 방법을 이용하여 실험을 진행하였다.
(a) (b) (c)
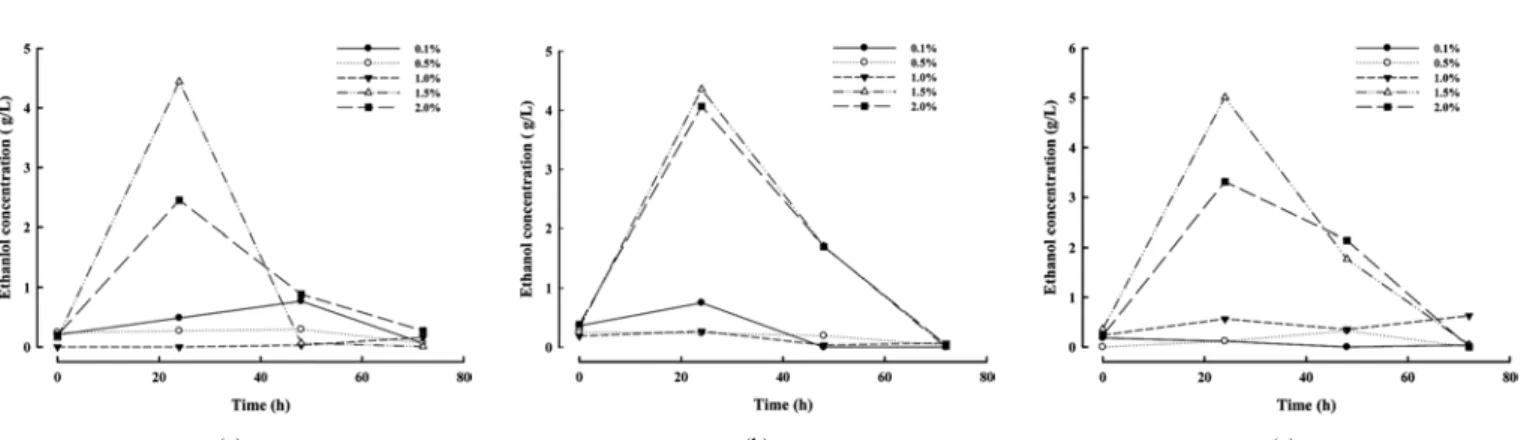
Figure 3. Ethanol production from Gelidium amansii : H 2 SO 4 and hydrolysis enzyme (celluclast) was used in the experiment. The celluclast was treated for 2 h in the shaking incubator at 35 ℃. (a) Acid hydrolysis, (b) simultaneous saccharification and fermentation, (c) separate hydrolysis fermentation.
(a) (b) (c)
Figure 4. Time courses of reducing sugar concentration during ethanol fermentation : H 2 SO 4 and hydrolysis enzyme (celluclast) wasused in the experiment. The celluclast was treated for 2 hin the shaking incubator at 35 ℃. (a) Acid hydrolysis, (b) simultaneous saccharification and fermentation, (c) separate hydrolysis fermentation.
즉, 산 처리만 한 배지를 이용한 발효, celluclast와 전 배양시켜 놓은 균을 함께 첨가 하는 동시당화발효(simultaneous saccharification and fermentation), 그리고 효소 최적 처리시간인 2 h 동안 celluclast를 첨가 하여 반응시킨 배지에 균을 분리하여 첨가하는 분리당화발효(separate hydrolysis fermentation) 를 각각 실험하여 비교하였다.
우뭇가사리가 첨가 되어 있는 배지에 산을 농도별로 처리하여 24 h 마다 시료를 추출하였다. 에탄올을 최대로 생산하는 산의 농도는 1.5% 이며, 가장 많이 생산되는 시간은 24 h 배양 하였을 때로 확인되 었다(Figure 3). 기본 배지에 산 처리만 하였을 경우 4.44 g/L의 에탄올 이 생산되었으며, 분리당화발효의 결과 4.35 g/L, 효소와 균을 동시에 첨가한 동시당화발효에서 5.00 g/L의 에탄올이 생산되는 것으로 확인 되었다. 산 전처리와 분리당화발효는 에탄올 생산량이 거의 차이가 없는 것으로 나타났다. 그러나 동시당화발효에서는 에탄올 생산량의 증가가 확인되었으며 이는 효소의 첨가에 의해 glucose의 함량이 증가 했기 때문인 것으로 보인다. 따라서 우뭇가사리 배지에 산 처리만 하 는 것보다 효소와 균을 동시에 첨가해 배양하는 것이 에탄올 생산에 더욱 효과적 이라는 것을 확인할 수 있었다.
4. 결 론
홍조류 중 우뭇가사리와 도박에 H 2 SO 4 과 CH 3 COOH 을 0.1%에서부터
2.0% 까지 각각 농도 별로 처리하였다. 우뭇가사리와 도박의 경우
H 2 SO 4 으로 처리한 것이 CH 3 COOH 으로 처리한 것보다 galactose의 생
성이 보다 높은 것으로 나타났다. 우뭇가사리는 H 2 SO 4 을 1.5%의 농
도로 처리하였을 때 7.04 g/L, 도박은 우뭇가사리와 동일한 조건에서
6.38 g/L의 galactose를 생산하였다. 환원당 생성에 효소처리가 어떠한
영향을 미치는지 확인해 보기 위해서 효소를 산 처리가 되어있는 우뭇
가사리 배지에 접종 시켜서 가수분해 정도를 알아보았다. 효소 처리
전과 비교했을 때 2 h 배양한 경우 2.49 g/L의 glucose와 3.71 g/L gal-
actose 가 생산되었다. 이후 효소의 처리가 바이오 에탄올 생산에 미치는
직접적인 영향을 확인하기 위해 Saccharomyces cerevisiae KCCM1129
를 이용하여 직접적인 에탄올 발효 실험을 진행하였다. 우뭇가사리에
산 전처리한 배지, 우뭇가사리에 산 전처리를 먼저하고 균과 효소를
따로 넣는 방법, 우뭇가사리에 산 전처리를 한 후 균과 효소를 함께
넣는 방법으로 나누어 실험하였다. 세 가지 배양 조건은 공통적으로
배양시간이 24 h이면서 처리한 H 2 SO 4 의 농도가 1.5%일 때에 가장
많은 에탄올 생산을 하였다. H 2 SO 4 만을 이용하여 가수분해한 경우와
효소반응과 에탄올 발효를 분리하여 배양한 경우 에탄올 생산량은 각
각 4.44 g/L와 4.35 g/L로 큰 차이를 보이지 않았다. 그러나 우뭇가사
리 효소반응과 발효를 동시에 진행한 동시당화발효에서는 최대 5.00
g/L 의 에탄올을 생산하여 공정을 분리하였을 때에 비해 114.94% 증
가한 것을 확인할 수 있었다. 또한 에탄올 발효 과정에서 일어난 배지
의 환원당 정도를 파악하기 위해서 이후 실험을 진행하다. DNS법을 이용해 환원당을 확인한 경우 배양시간 48 h까지 지속적으로 감소하 는 것으로 나타났다. 배양시간이 48 h 전까지는 최대 37%까지의 환원 당 감소가 일어났지만, 이후에는 16%까지의 환원당이 소비되었다.
감 사
본 연구는 국토해양부 소관 해양생명공학기술개발사업의 연구비 지원에 의해 수행되었습니다.
참 고 문 헌