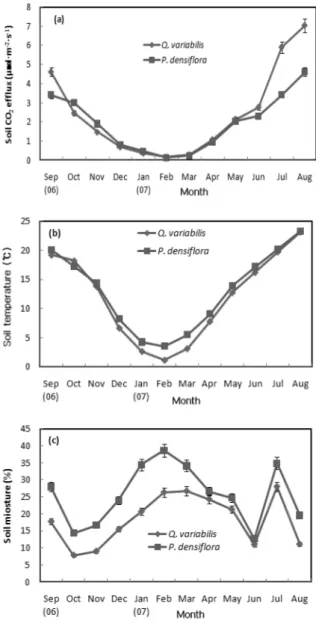
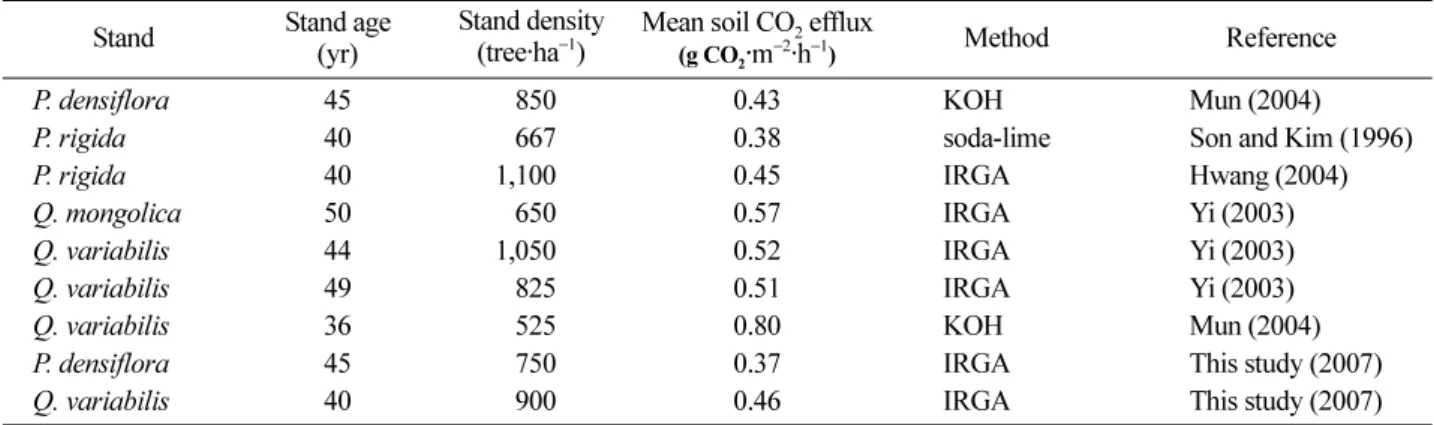
관련 문서
Days to emergence and heading, duration of growth, leaf traits and stem diameter of sweet sorghum as affected by soils. Soil type Days
A laboratory experiment was performed to investigate the removal of nitrogen and phosphorus from plating factory effluent using the soil column.. Soil,
The purpose of this study was to compare the microleakage of different adhesive systems (three 2-step adhesives and two 1-step adhesives) in class V
In this study, we investigated the effects of low-power CO 2 laser on proliferation on human gingival fibroblast cells so that determine laser
In this study, it is shows that the stream of CO 2 is more effective in the reduction of unreacted TEGDMA and the increase of surface microhardness than that of N 2
Rasmussen, The use of soil-gas CO 2 in the exploration for sulfide-bearing breccia pipes in northern Arizona, 1990, Journal of Geochemical Exploration, vol.38, pp..
In this context, this study compared and analyzed a variety of soil decontamination methods used to evaluate the wide-area soil contaminated with Cs:
To estimate the change of riverbed geography by the operation of movable weir, two-dimensional RMA-2 and SED-2D were used in this study. Hydraulic elements