반지련 (Scutellaria barbata D. Don) 추출물이 lipopolysaccharide에 의해 활성화된 대식세포에 미치는 영향
센 팅*·이용진**·조재열*†
*강원대학교 BT학부대학 생명공학부, **(재) 춘천바이오산업진흥원
Effect of Hot Water Extract from Scutellaria barbata on the Macrophages Activated by Lipopolysaccharide
Ting Shen*, Yong Jin Lee**, and Jae Youl Cho*†
*
School of Bioscience and Biotechnology, and Institute of Bioscience and Biotechnology, Kangwon National University, Chuncheon 200-701, Korea.
**
Laboratory of functional food development, Chuncheon Bioindustry Foundation, Chuncheon 200-957, Korea.
ABSTRACT : Scutellaria barbata was examined to evaluate its modulatory effects on the functional activation of macroph- ages under lipopolysaccharide (LPS) treatment. To do this, hot water extract (Sb-HWE) was prepared from Scutellaria bar- bata and several inflammatory parameters such as nitric oxide (NO) production, phagocytosis, reactive oxygen species (ROS) determination and intracellular signaling pathway were selected to be tested. Sb-HWE strongly blocked NO produc- tion in LPS-activated RAW264.7 cells in a dose-dependent manner. However, it did not suppress inducible NO synthase (iNOS). In agreement, Sb-HWE did not diminish inflammatory signaling composed of NF- κ B and its upstream activation signaling enzymes such as Akt and I κ B α . Sb-HWE protected RAW264.7 cells from LPS-induced cytotoxicity up to 80% at 400 ㎍ / ㎖ . Furthermore, this extract blocked phagocytic uptake of FITC-dextran, while sodium nitroprusside (SNP)- induced ROS generation in RAW264.7 cells was not decreased. Therefore, our data suggest that Sb-HWE may have differ- ential immunoregulatory function depending on macrophage-mediated immune responses.
Key Words : Scutellaria barbata, Hot Water Extract, Macrophages, NO Production, Phagocytosis, ROS Generation
대식세포는 내재성 면역반응을 담당하는 면역세포로서 , 외부 로부터 침입한 세균들에서 유래된 세포구성 성분들 [ 예를들면 , lipopolysaccharide (LPS)] 이나 체내 다른 면역세포에서 분비한
사이토카인 [ 예를 들면 , interferon (IFN)- γ에 의해 활성화되어 종 양괴사인자 (tumor necrosis factor- α ) 와 같은 사이토카인이나 일산화질소 (nitric oxide: NO) 나 활성산소 (reactive oxygen species: ROS) 등과 같은 독성물질 , 그리고 프로스타그란딘류와 같은 염증 매개물질의 분비를 조절한다 (Tetley, 2005). 특별히 ,
이들 세포는 세균이나 암세포 등과 같은 면역유발원을 탐식하 고 탐식된 면역원을 펩타이드로 가공하여 주조직 적합성 항원 에 공여함으로써 , T cell 의 분열 및 분화를 촉진시키는 항원제 시 세포 (Antigen-presenting cell [APC]) 로서의 역할을 수행하 는 것으로 알려져 있다 (Edwards et al ., 2006). 이들 단백질의 활성 및 발현은 외부 자극에 반응하는 pattern-recognition receptor 와 같은 수용체의 매개 및 이들 수용체와 결합된 신호
전달 단백질 복합체의 활성에 의해 조절되는 것으로 알려져 있 다 . 특별히 관련 신호전달 단백질은 non-receptor type tyrosine kinases (Src, Syk 및 JAK2), phosphoinositide-3-kinase (PI3K), Akt (protein kinase B) 및 mitogen-activated protein kinases (MAPK) 등으로 구성되어 있다 (Fang et al ., 2004).
이들 단백질의 활성은 최종적으로 NF- κ B 및 AP-1 과 같은
transcription factor 의 활성과 밀접한 관련이 있으며 , 이들의 활 성은 염증 및 내재성 면역반응 관련 단백질의 발현을 매개하게 된다 (Chen & Shi, 2002; Dijkstra et al. , 2002).
반지련 [ Scutellaria barbata D. Don (Lamiaceae)] 은 성질
은 차고 독은 없으며 약간 쓴맛이 나는 약재이다 (Jiangsu
New Medical College, 1977). 현재까지의 주성분으로서 neo- clerodane diterpenoids 류 및 flavonoid 류 등이 반지련의 주요 활성 성분으로 확인되었다 (Dai et al. , 2007; Sato et al. , 2000). 반지련은 혈액순환을 돕고 , 인후염 , 편도선염 , 타박상
†
Corresponding author: (Phone) +82-33-250-6488 (E-mail) [email protected]
Received August 27 , 2008 / Revised September 29 / Accepted Octber 8, 2008
및 피부발진 등의 치료 효능이 있는 것으로 보고되어져 있다 .
특별히 최근에는 항암효과가 있음이 입증되어 자궁암 , 간암 ,
직장암 , 식도암 , 유방암 , 폐암 , 비인암 , 및 융모상피암 등에 널 리 처방되고 있다 (Dai et al. , 2008; Suh et al. , 2007). 그 러나 아직까지 자세한 면역관련 기전연구나 면역질환의 치료 효능에 관한 구체적인 치료기전 연구들이 진행된 바는 없다 .
본 연구에서는 아직까지 보고되지 않은 대식세포에 의해 수 행되는 내재성 면역반응에 관한 반지련 열수추출물의 조절효 과를 확인해 보고자 한다 . 즉 , LPS 에 의해 활성화된 대식세포
의 주요 기능인 nitric oxide (NO) 생성 , 탐식작용 및 대식세 포 활성신호전달 과정 등에 있어서 반지련 열수 추출물의 조 절 작용에 대해 검토하였다 .
재료 및 방법
1. 실험재료
Kim 등 (2005) 의 방법에 의해 제조된 Scutellaria barbata 의 열수추출물 (Sb-HWE) 은 상지대학교 한의과대학 이선구 박 사님으로부터 제공받았다 . 실험에 사용된 대식세포주는 마우 스 RAW264.7 세포를 사용하였다 . 세포배양을 위한 배양용 배 지 RPMI1640 media, fetal bovine serum (FBS) 그리고 항 생제 (penicillin/streptomycin) 는 Hyclone (Hyclone, South Logan, UT, USA) 사 제품을 사용하였다 . Akt 및 I κ B α에 대 한 phospho-specific 항체 및 β -actin 항체는 Cell Signaling Technologies 사 (Danvers, MA, USA) 제품을 사용하였다 . 그 밖에 , DHR123, sodium nitroprusside (SNP), FITC-dextran
및 MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphinyltetrazolium bromide) 를 포함한 화학시약은 Sigma 제품 (St. Louis, MO, USA) 을 사용하였다 .
2. 세포배양
RAW 264.7 및 U937 세포들은 5% CO
2, 37 ℃ incubator
에서 배양하였다 . 배지는 penicillin (100 IU/ ㎖ ) 및 strepto- mycin (100 ㎍ / ㎖ ) 과 10% FBS 가 포함된 RPMI 1640 을 사 용하였다 .
3. 세포 생존율 검정
반지련 열수추출물 (Sb-HWE) 을 다양한 농도 (0, 12.5, 25, 50, 100, 200 및 400 ㎍ / ㎖ ) 별로 처리한 후 세포생존에 미치 는 영향을 MTT assay 법으로 분석하였다 (Cho, 2007). 96- well plate 에 1 × 10
6개의 RAW264.7 세포를 plating 하고 Sb- HWE 를 농도 별로 처리하였다 . 37 ℃에서 각 면역실험 조건에 상응하는 배양시간 동안 CO
2incubator 에서 배양하였다 . 이후 10 ㎕ MTT 용액 (stock concentration : 5 ㎎ / ㎖ ) 을 첨가하고 3 시간 동안 추가반응을 유도하였다 . 반응 종료 및 formazan
crystal 의 용해를 위해 각 well 에 100 ㎕ MTT stopping solution (10% sodium dodecyl sulfate in 0.01 M HCl) 을 분주하였다 .
세포 생존율은 MTT 가 formazan 으로 환원된 양을 570 ㎚에서 흡광도를 측정하여 얻어진 OD 값을 통해 산출하였다 .
4. Nitric Oxide의 정량
배지에서의 nitric oxide 정량은 Sb-HWE 와 LPS 가 처리된
RAW264.7 세포 (2 × 10
6cell/ ㎖ ) 를 이용하여 실시하였다 (Cho et al ., 2001). 배양 배지 100 ㎕와 Griess 시약 [5% (v/
v) phoshophoric acid 용액 내 1% (w/v) sulfanilamide 및
0.1% (w/v) naphthylethylenediamide] 100 ㎕를 혼합한 후 상 온에서 10 분 동안 방치한 후에 발색된 정도를 540 ㎚에서
microplate reader 를 이용하여 측정하였다 . 표준 정량곡선은
sodium nitrite 를 적정한 농도로 희석하여 동일 방법으로 흡광 도를 측정한 후 완성하였다 .
5. RT-PCR법
iNOS 발현 정도를 전사수준에서 조사하기 위해 각 시료를
일정시간 동안 처리하고 Trizol reagent 를 사용하여 total RNA
를 추출하였다 . 추출한 total RNA 를 First strand cDNA synthesis kit (Fermentas) 를 사용하여 cDNA 를 제조한 다음 ,
동량의 cDNA 를 PCR 로 증폭하였다 . PCR amplication 은 i- Master PCR kit (iNtRON) 을 사용하여 각 실험군 cDNA 와 표적단백질들의 sense 및 antisense primers, 대조군 GAPDH primers 를 dNTP 250 mM, Tris-HCL (pH8.3) 10 mM, KCl 50 mM, NgCl
21.5 mM 를 포함한 i-mater solution 20 ul 에서 시행하였다 . PCR 은 95 ℃에서 45 초 간 denaturing, 55 ℃에서
45 초 간 annealing 그리고 72 ℃에서 1 분간 extension 하는 조 건으로 시행하며 , 총 30 cycles 을 수행하였다 . PCR 로 증폭된 DNA 는 1.5% agarose gel 에서 전기영동하였고 분획된 DNA band 의 intensity 를 측정하였다 . 실험에 사용된 DNA primer 의 서열은 Table 1 에 정리되어 있다 .
6. NF-κB reporter gene assay
HEK293 세포 (1 × 10
6cells/well) 를 12 시간 동안 전배양한 후 , calcium-phosphate 법을 이용하여 NF- κ B-luciferase plasmid (1 ㎍ ) 를 transfection 하였다 . 48 시간이 지난 후 세포에 Sb- HWE 및 TNF- α (20 ng/ ㎖ ) 를 각 농도 별로 처리하고 18 시 간 배양 후 , luciferase 활성을 조사하기 위해 세포를 harvest
하였다 . Luciferase assay 는 Promega 에서 제공한 luciferase
assay system 을 이용하여 세포파쇄 후 정량하였다 .
Luminescence 는 luminometer (Thermo Labsystems Oy, Helsinki, Finland) 를 이용하여 측정하였다 . Transfection 의 효
율에 관한 normalization 은 β -galactosidase 활성을 측정하여
실시하였다 .
7. Western blotting
RAW263.7 세포 (5 × 10
6cells/well) 에 Sb-HWE 및 LPS (2 ㎍ / ㎖ ) 를 각 농도 별로 처리하고 세포들을 lysis buffer 에서 용해시킨 후 total lysate 단백질을 12,000 × g 에서 15 분간 원
심분리 하였다 . 동일한 양의 단백질을 10% SDS-
polyacrylamide gels 에서 전기영동하고 Wet-blotting transfer
방법을 이용하여 PVDF membranes 으로 단백질을 transfer 하였 다 . Membrane 을 5% skim milk 를 포함하는 blocking buffer
에서 반응시킨 후 특정 단백질에 특이한 1 차 항체를 4 ℃에서 하루 동안 반응시켰다 . 이 후 2 차 항체를 적용한 다음 ECL chemiluminescence 로 반응 정도를 확인하였다 .
8. 라디칼 소거능 정량
RAW264.7 세포를 penicillin (100 IU/ ㎖ ) 및 streptomycin (100 ㎍ / ㎖ ) 과 10% 의 FBS 를 함유하는 RPMI 1640 배지를
이용해서 2 × 10
6cells/ ㎖의 농도로 조절한 후 , 6 well plate
에 1 ㎖를 분주하고 , 5% CO
2및 37 ℃에서 18 시간 동안 전 배양하였다 (Lee et al ., 2007b). 전배양 후 배지를 제거하고 , PBS 495 ㎕ 를 분주하였다 . 그리고 4 배 농도로 조제된
DHR123 ( 최종농도 20 µ M) 250 ㎕를 해당 well 에 분주 후
10 분 배양하였다 . 이후 , 200 배 농도로 조제된 Sb-HWE 5 ㎕ 를 처리 후 30 분 배양하고 , 4 배 농도로 조제된 SNP 250 ㎕ ( 최종농도 0.25 mM) 를 처리하여 20 분 배양하였다 . 라디칼 소 거 정도는 flow cytometer 를 이용하여 측정하였다 .
9. 탐식작용 측정
RAW264.7 세포를 2 × 10
6cells/well 로 조정하여 6-well plate
에 분주하고 18 시간 동안 전배양하였다 . 전배양 후 Sb-HWE
를 처리하고 1 시간 동안 37 ℃에서 배양하였다 . 그리고 형광으
로 표지된 particle FITC-dextran (1 ㎎ / ㎖ ) 을 세포에 처리하고 빛을 차단하여 30 분간 배양하였다 . Cold-PBS 로 세포를 세척하 여 미탐식된 입자를 제거하고 3.7% formaldehyde 로 고정 후
flow cytometer 로 형광정도를 측정하였다 (Lee et al. , 2007a).
10. 통계처리
각 data 는 얻어진 결과는 평균 ± 표준편차로 나타낸 것이며 ,
그룹 간의 통계적 유의성은 SAS package 를 이용하여 P <
0.05 수준에서 Duncan 의 다중비교법에 의해 분석하였다 .
결과 및 고찰
1. S. barata 유래 열수 추출물 (Sb-HWE)의 세포독성
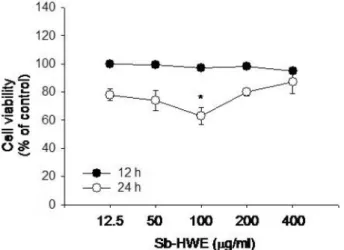
Sb-HWE 의 세포독성 효과를 알아보기 위해 최대 400 ㎍ / ㎖ 농도까지 대식세포주 (RAW264.7 cells) 에 12 및 24 시간 동안 처리하였다 . Fig. 1 에서 나타난 바와 같이 Sb-HWE 은 400
㎍ / ㎖ 농도까지도 마우스 대식세포인 RAW264.7 세포에 아무 런 독성효과를 나타내지 않았다 . 따라서 면역약리학적 효능을 조사하기 위해 , 활성화된 대식세포를 이용하여 Sb-HWE 의 조
절능을 확인하였다 .
2. NO 분비에 미치는 Sb-HWE의 효과
혈관 확장조절 기능 이외에도 NO 는 대식세포에서 분비되는 대표적인 세포사멸 유도 독성물질이다 (Maiorana et al. , 2003; Taylor et al. , 2003). 특별히 NO 는 대식세포의 내재성 면역반응 수행시 유리되는 주요 면역물질로 알려져 있다
(Brune et al ., 2003). 즉 , 염증이나 면역반응을 유도하는 물질 에 의해 활성화된 대식세포는 inducible NO synthase (iNOS)
의 발현을 유도하여 NO 를 분비하게 된다 . 따라서 , 본 연구에 서는 먼저 반지련에 의한 대식세포 면역조절능 탐색을 위해
LPS 에 의해 활성화된 대식세포로부터 NO 분비에 미치는 영 향을 조사하였다 .
흥미롭게도 반지련으로 부터 얻어진 열수추출물 (Sb-HWE)
은 농도의존적으로 NO 생성을 억제한 것으로 나타났다 . 즉
400 ㎍ / ㎖의 농도로 처리된 시료에서는 최대 60% 까지 NO 생 성을 억제하는 것으로 확인되었다 (Fig. 2A). 게다가 , 이들 농
Fig. 1.Effect of Sb-HWE on the viability of RAW264.7 cells.
RAW264.7 cells (1
×10
6cells/
㎖) were incubated with Sb-HWE for 12 or 24 h. Cytotoxicity of this drug was determined by conventional MTT assay as described in Materials and Methods. Data represent mean ± SEM of three independent observations performed in triplicate.
*:p < 0.05 compared to normal.
Table 1.
Primer list used in this experiment (F: forward, R: reverse).
Gene Primer sequence
iNOS F 5'-CCCTTCCGAAGTTTCTGGCAGCAGC-3' R 5'-GGCTGTCAGAGCCTCGTGGCTTTGG-3' GAPDH F 5'-CACTCACGGCAAATTCAACGGCAC-3'
R 5'-GACTCCACGACATACTCAGCAC-3'
도에서는 세포독성을 유도하지 않았기 때문에 (Fig. 1), NO
억제 효과는 반지련의 비특이적인 세포독성 작용에 의해 기인 된 것으로 판단되지는 않았다 .
앞서 언급한대로 , NO 는 iNOS 의 활성에 의해 매개되므로 , iNOS mRNA 를 조사하여 이들 억제 작용이 mRNA 의 전사과 정 억제와 관련이 있는지 조사하여 보았다 . Fig. 2B 에서 확인
할 수 있듯이 , 반지련 열수추출물은 LPS 에 의해 유도된
iNOS 발현에 전혀 영향을 나타내지 않았다 . 또한 , Sb-HWE 는
iNOS 발현을 조절하는 대표적인 전사인자인 NF- κ B 의 활성을 억제하지 못하였다 (Fig. 2C). 즉 NF- κ B reporter gene assay 에 따르면 , Sb-HWE 는 PMA 에 의해 유도된 luciferase
활성을 전혀 억제하지 못하였으며 , 오히려 활성을 상승시킨 것
으로 보아 NF- κ B 의 binding 활성을 더 유도한 것으로 판단된
다 . 게다가 , 반지련의 Sb-HWE 는 NF- κ B 활성을 조절하는 상 위 신호전달 단백질의 인산화 과정을 억제하지 못하였다 . 즉
Fig. 2D 에서 관찰할 수 있듯이 , LPS 에 의해 유도된 Akt 및
I κ B α의 활성을 전혀 저해하지 못한 것으로 확인되었다 . 따라 서 본 결과로 미루어볼 때 , 반지련에 의한 NO 억제작용은 더 덕 추출물 (Lee et al. , 2007b), 케나프추출물 (Lee et al. , 2007a), curcumin (Lee & Cho, 2007) 및 davallialactone (Lee et al ., 2008) 등 에서 관찰되는 transcription factor (NF- κ B) 억제기전과는 다른 방법에서 진행되는 것으로 판단
된다 . Activator protein (AP)-1 및 cAMP responsive element binding protein (CREB) 등과 같은 전사인자의 발현이 NF-
κ B 비의존성 염증유발 관련 단백질의 발현을 조절한다고 보 고된 바가 있다 (Chen et al ., 2008). 그러나 , Fig. 2B 에서 보여지듯 , 반지련 분획이 iNOS 발현을 억제하지 않았다는 사 실은 , 반지련 활성성분에 의한 억제과정이 단순히 iNOS
mRNA 의 전사수준 보다는 iNOS 의 단백질 번역과정이나
iNOS mRNA 의 stability 수준에서 진행될 가능성이 있다는 것
을 시사한다 . 실제로 AH23848 및 AgC10 등은 이들 현상을
통해 염증물질의 생성을 억제하는 것으로 보고되어져 있다
Fig. 2.Effects of Sb-HWE on NO production, iNOS mRNA level, and NF-
κB-relevant signaling in LPS-treated RAW264.7 cells. (A)
RAW264.7 cells (1
×10
6cells/
㎖) were incubated with Sb-HWE in the presence or absence of LPS (2
㎍/
㎖) for 12 or 24 h. NO
levels were determined by Griess assay as described in Materials and Methods. (B) RAW264.7 cells (5
×10
6cells/
㎖) were
incubated with Sb-HWE in the presence or absence of LPS (2
㎍/
㎖) for 2 and 30 min. iNOS mRNA levels were analyzed by RT-
PCR as described in Materials and Methods. (C) HEK293 cells (5
×10
6cells/
㎖) were incubated with Sb-HWE in the presence or
absence of TNF-
α(20 ng/
㎖) for 18 h. Luciferase activity was determined by luminometer as described in Materials and
Methods. Data (A, C) represent mean ± SEM of three independent observations performed in triplicate. (D) RAW264.7 cells
(5
×10
6cells/
㎖) were incubated with Sb-HWE in the presence or absence of LPS (2
㎍/
㎖) for 15 and 30 min. After
immunoblotting, the total or phospho-protein levels of Akt, I
κB
α, and
β-actin were identified by their total protein- or phospho-
specific antibodies. The results (B and C) show one experiment out of three. *:p < 0.05 and **:p < 0.01 compared to control.
(Lin et al ., 2005; Lin et al ., 2008). 관련 내용들에 관한 추 가적인 연구는 차후 진행될 예정이다 .
한편 NO 생성을 유도하는 LPS 나 IFN- γ와 같은 대식세포
활성 자극원들은 24 시간이나 48 시간 처리시 이들 물질에 의 해 분비된 NO 에 기인된 세포독성 현상 ( 세포사멸 ) 을 유도하 게 된다 (Lee et al ., 2006). 실제로 Fig. 4 에서도 정상군
(100%) 대비 40% 에서 50% 정도의 세포가 사멸된 결과를 확 인할 수 있었으며 , 본 연구자는 이들 현상이 NO 억제제인 N-
MMA 처리시 완벽히 저해될 수 있다는 것을 보고한 바 있다
(Cho et al ., 2001). Fig. 4 는 확실히 Sb-HWE 가 이들 대식세
포 활성 자극원에 의한 세포독성 과정을 매우 효과적으로 억 제할 수 있다는 것을 보여주고 있는데 , 특별히 400 ㎍ / ㎖ 농
도에서는 거의 정상군의 80% 수준까지 세포사멸로부터
RAW264.7 cell 을 보호하는 것으로 나타났다 . 더덕 추출물이나
케나프 추출물에서도 확인되었듯이 (Lee et al. , 2007a; Lee
et al ., 2007b), Sb-HWE 의 세포독성 보호효과는 NO 억제효과 에서 기인된 것으로 판단된다 .
3. Sb-HWE의 라디칼 소거능
ROS 는 대개 산소가 필요한 대사과정 중에서 부산물로 생성
되는데 , 세포의 신호전달에서 중요한 역할을 수행하기도 한다 (Forman & Torres, 2001). 그러나 환경적인 스트레스로 인해
ROS 가 급격히 증가되면 , 라디칼이 발생되어 세포구조에 심각 한 손상을 초래하여 세포사멸과 같은 현상을 유도하게 된다
(Migliaccio et al ., 2006). 반면에 면역반응에서는 활성산소 유 래 라디칼을 이용하여 대식세포나 과립구 등이 몸 안으로 침 입한 미생물이나 종양세포를 공격하는 방어용 수단으로 사용 한다 . 병리적으로는 이들 활성산소종 유래 라디칼이 과량으로
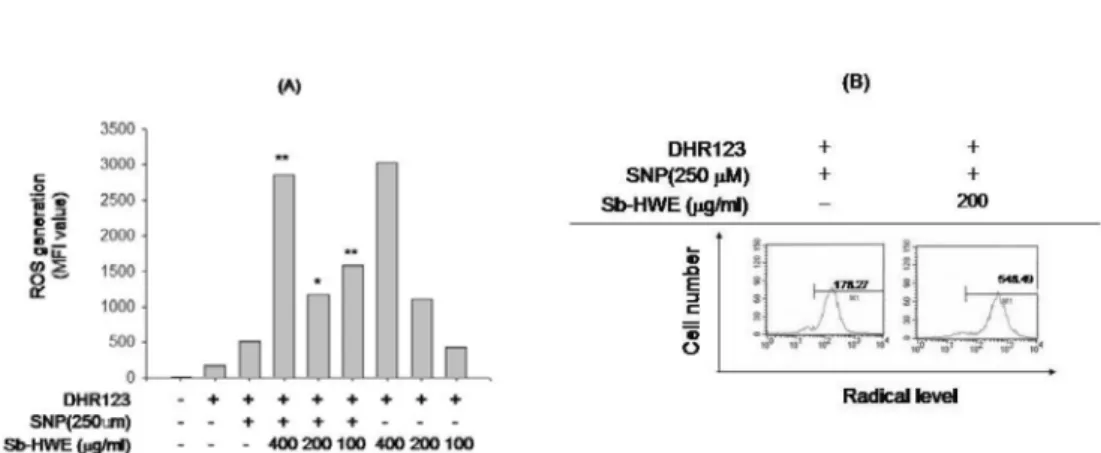
분비될 경우 노화나 암 및 염증질환의 발생과 깊은 관련이 있 어 ROS 의 라디칼 소거능을 가지는 플라보노이드류 같은 항 산화제들은 관련 질환의 치료 효능이 있는 것으로 보고되고 있다 (Katiyar et al ., 1999). 따라서 , Sb-HWE 이 대식세포내 에서 유도되는 라디칼 발생에 어떤 영향을 미치는지 조사하였 다 . 그러나 , Fig. 3 에서 보여지듯이 , Sb-HWE 은 SNP 에 의해
유리되는 라디칼을 중화시키지 못하는 것으로 나타났다 . 오히
려 , 라디칼 생성을 더 증가시킨 것으로 나타났다 . 반면에 대조 군으로 사용된 토코페롤은 강력하게 라디칼 생성을 억제하는 것으로 확인되었다 (data not shown). 따라서 본 결과로 미루 어 볼 때 , 반지련의 NO 생성 억제효능은 라디칼 중화에서 기 인된 것은 아닌 것으로 판단된다 .
4. 식세포 작용에 미치는 Sb-HWE의 효과
내재성 면역반응에서 보여지는 중요한 현상이 식세포 ( 대식 세포와 호중구 ) 매개에 의한 식세포 작용으로 알려져 있다
Fig. 3.Scavenging effect of Sb-HWE on SNP-released radical generation. (A and B) RAW264.7 cells (1
×10
6) were pretreated with Sb- HWE for 30 min and further incubated in the presence of absence of SNP (250
µM) for 0.5 h. The level of radicals was determined by flow cytometric analysis as described in Materials and Methods. *:p < 0.05 and **:p < 0.01 compared to control.
Fig. 4.
Cytoprotective effect of Sb-HWE on the LPS-induced cell death of RAW264.7 cells. RAW264.7 cells (1
×10
6cells/
㎖) were incubated with Sb-HWE for 12 or 24 h.
LPS-induced cell death was determined by conventional
MTT assay as described in Materials and Methods. Data
represent mean ± SEM of three independent observations
performed in triplicate. *:p < 0.05 and **:p < 0.01
compared to control.
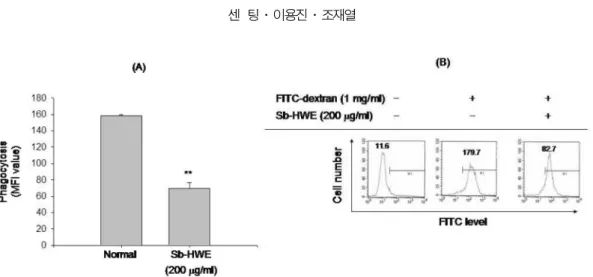
(Savina & Amigorena, 2007). 특별히 이 과정은 세균들을 제 거하기 위한 과정이면서 동시에 APC 의 항원가공 및 제시 기 능을 위해 필수적인 염증 단계의 하나이므로 Sb-HWE 의 조절 작용을 조사하여 보았다 . 그러나 Fig. 5 에서 확인되듯이 , Sb- HW 은 마우스 대식세포주인 RAW264.7 세포에서 보여지는
FITC-dextran 의 식세포작용을 200 ㎍ / ㎖ 농도에서 50% 정도
억제하였다 .
적 요
S. barbata의 열수 추출물 (Sb-HWE) 은 대표적인 염증과정 인 LPS 에 의해 활성화된 대식세포로 부터의 NO 생성 , LPS
매개에 의한 세포사멸작용 , 및 FITC-dextran 의 대식세포내 탐
식작용을 매우 효과적으로 억제하였다 . 그러나 본 추출물은
SNP 로 유도된 라디칼 소거능은 매우 미약한 것으로 나타났다 .
NF- κ B- 매개에 의한 루시퍼라제 활성 , 및 NF- κ B 활성 관련 신호전달 단백질 (Akt 및 I κ B α ) 에 대한 저해작용은 관찰되지 않은 것으로 보아 이들 추출물의 대식세포 면역반응 조절 기 전은 기존의 알려진 방법과는 다른 기전에 의해 진행되는 것 으로 판단된다 .
감사의 글
반지련의 열수 추출물을 제공해주신 상지대학교 이선구 박 사님께 감사드린다 .
LITERATURE CITED
Brune B, Zhou J, von Knethen A (2003) Nitric oxide, oxidative stress, and apoptosis. Kidney Int. Suppl. 84:S22-24.
Chen F, Shi X (2002) NF-kappaB, a pivotal transcription factor in silica-induced diseases. Mol. Cell. Biochem. 234-235(1-2):169- Chen LG, Hung LY, Tsai KW, Pan YS, Tsai YD, Li YZ, Liu 176.
YW (2008) Wogonin, a bioactive flavonoid in herbal tea, inhibits inflammatory cyclooxygenase-2 gene expression in human lung epithelial cancer cells. Mol. Nutr. Food Res. May 21. [Epub ahead of print].
Cho JY (2007) Inhibitory effect of extracts from woody plants on tumor necrosis factor-alpha production in lipopolysaccharide- stimulated RAW264.7 cells. Korean J. Medicinal Crop. Sci.
15(4):271-275.
Cho JY, Park J, Kim PS, Yoo ES, Baik KU, Park MH (2001) Savinin, a lignan from
Pterocarpus santalinusinhibits tumor necrosis factor-alpha production and T cell proliferation. Biol.
Pharm. Bull. 24(2):167-171.
Dai SJ, Liang DD, Ren Y, Liu K, Shen L (2008) New neo- clerodane diterpenoid alkaloids from
Scutellaria barbatawith cytotoxic activities. Chem. Pharm. Bull. (Tokyo) 56(2):207-209.
Dai SJ, Wang GF, Chen M, Liu K, Shen L (2007) Five new neo-clerodane diterpenoid alkaloids from
Scutellaria barbatawith cytotoxic activities. Chem. Pharm. Bull. (Tokyo) 55(8):1218-1221.
Dijkstra G, Moshage H, Jansen PL (2002) Blockade of NF- kappaB activation and donation of nitric oxide: new treatment options in inflammatory bowel disease? Scand. J. Gastroenterol.
Suppl. 236:37-41.
Edwards JP, Zhang X, Frauwirth KA, Mosser DM (2006) Biochemical and functional characterization of three activated macrophage populations. J. Leukoc. Biol. 80(6):1298-1307.
Fang H, Pengal RA, Cao X, Ganesan LP, Wewers MD, Marsh CB, Tridandapani S (2004) Lipopolysaccharide-induced macrophage inflammatory response is regulated by SHIP. J.
Immunol. 173(1):360-366.
Forman HJ, Torres M (2001) Signaling by the respiratory burst in macrophages. IUBMB Life 51(6):365-371.
Jiangsu New Medical College (1997). Dictionary of Chinese Materia Medica. Science and Technology Press of Shanghai,
Fig. 5.