Copyrightⓒ 2010, The Korean Academy of Oral Biology
137
Journal of Oral Biology
Properties of Extremely Low Frequency Electromagnetic Fields and their Effects on Mouse Testicular Germ Cells
Yeon Sook Kim and Suk Keun Lee1*
Department of Dental Hygiene, College of Health, Cheongju University, Cheongju, Korea
1Department of Oral Pathology, College of Dentistry, Gangneung-Wonju National University, Gangneung, Korea (received July 14, 2010 ; revised November 6, 2010 ; accepted November 12, 2010)
To evaluate the biohazard properties of an extremely low frequency electromagnetic field (ELF-EMF), we explored the physical properties of the ELF-EMF that generates the electric current induction in the secondary coil from the chamber of a primary solenoid coil. We subsequently ex- plored the biological effects of a strong alternating electro- magnetic field (EMF), ranging from 730-960 Gauss, on the mouse testis. Mice were exposed to an alternating EMF field induced by a rectangular electric current at 1, 7, 20, 40, and 80 Hertz, for 1, 3, 5, and 7 hours. The mouse testes were examined for proliferative activity and apoptosis using the in situ terminal deoxynucleotidyl transferase (TdT) method and by immunostaining of proliferating cell nuclear antigen (PCNA), respectively. We found that the electric current induction increased in the 6-8 Hertz range, and that ex- posure to an ELF-EMF induced the apoptosis of mouse spermatocytes. In situ TdT staining was found to be most prominent in 7 Hertz group, and gradually reduced in the 20, 40, and 80 Hertz groups. These data suggest that a strong EMF can induce reproductive cell death within a short time, and the harmful effects of the EMF are maximal at low frequency alternating EMFs.
Key words: extremely low frequency, electromagnetic field, mouse, testis, apoptosis
Introduction
The potential effects of extremely low frequency (ELF) electromagnetic field (EMF) on living systems have been studied by many authors (Adey, 1980; Kirschvink et al., 1992;
Portet & Cabanes, 1988; Saunders & Jefferys, 2002). How- ever, the ELF-EMF induced variable biological effects, i.e., calcium ion efflux from brain tissue (Ayrapetyan et al., 1994;
Bawin et al., 1978; Blackman et al., 1979; Blackman et al., 1985; Grant et al., 1994), stimulation of healing in fractured bone (Bassett, 1984; Diniz et al., 2002; Inoue et al., 2002; Ko et al., 2008; Takano-Yamamoto et al., 1992), changes in spermatogenesis (De Vita et al., 1995), etc. It is generally agreed that the effects of ELF-EMF could be interpreted as resulting from electrical currents induced via the Faraday effect (Engstroem, 1996; Malko et al., 1994). Therefore, the biological effects of EMF could be influenced by the variation of frequency, flux density of EMF and exposure time. Many authors have observed variable biological effects using different conditions, i.e., structural changes in the hepatocytes of guinea pig after 0.005 Tesla and 0.3 Tesla constant EMF for 3 to 7 weeks (Parafiniuk et al., 1992), no influence of 1.5 Tesla constant EMF on growth of yeast cells (Malko et al., 1994), no gross impairment after prenatal exposure to 50 Hz/20 mT EMF on development in mice, enhanced teratogenic effects of cytosine arabinoside in mice exposed to pulsed EMF from video display terminals (Chiang et al., 1995), blockade of sensory neuron action potentials by a static EMF in the 10 mT range (Cavol et al., 1995), and low intensity millimeter wave EMF radiation stimulation of the regeneration of the sciatic nerve in rat (Kolosova et al., 1996). However, there are many controversies on these biological effects of various EMF.
Previously we designed a Magno-DR machine (Hanil Co., Korea) that produces a strong alternating EMF in order to
*Corresponding author: Suk Keun Lee, DDS, Ph.D., Department of Oral Pathology, College of Dentistry, Gangneung-Wonju National University, 123 Chibyun-dong, Gangneung, 210-702, Korea Tel : +82-33-640-2228, Fax : +82-33-642-6410
E-mail : [email protected], [email protected]
enhance the decalcification and devisceration of bone tissues.
We observed that the alternating EMF induced rapid decrease of pH in decalcification solutions, and the acidity slowly recovered to the original point when the alternating EMF exposure stopped (Lee et al., 1993). It is supposed that the alternating EMF can induce proton reaction and activate ionic free radicals in decalcification solutions. In this study we explored the hazardous effects of the alternating EMF on the mouse testes by detecting an increase of apoptotic cells.
Materials and methods
Magnetic exposure
We used the Magno-DR apparatus that was made for the enhancement of decalcification or devisceration of bony tissues. This machine produces ELF-EMF by square pulsating currents, changeable frequencies from 1 to 99 Hertz. The peak EMF, 730-960 Gauss, generated by the square pulsating cur- rents, from 1 to 90 Hertz, were measured by Tesla meter (Model 4048, F.W. Bell, U.S.A.) in the chamber of Magno- DR. To evaluate the physical property of the EMF a secondary enamel coil, 200 wire wraps around 1 cm diameter plastic tube with enamel coated wire (0.3 mm diameter), was placed in the center of Magno-DR chamber, and the induced electric cur- rents at different frequencies were analyzed by an oscilloscope (Tektronix TDS 380; Digital real-time, 400 MHz-2GS/s).
For the animal experiment the Magno-DR machine was redesigned to set sufficient fan ventilation, food and water supply. Air temperature within the exposure chamber was maintained at 20-21oC by fan ventilation. The sham treatment was performed in the similar temperature and noise conditions.
However, during field generation the noise level was up to 55 dB, while the ambient noise level, generated by air conditioner, refrigerator and freezer in the laboratory was about 60 dB.
Experimental design
Adult male mice at four to six weeks of age were divided into three groups; an EMF field exposure group (n=200), a sham treatment group (n=50), and a control group (n=10).
Fifty mice were used in one schedule of EMF exposure. We performed the schedule four times as follows. Two mice were placed in the Magno-DR chamber and exposed to EMF of 1, 7, 20, 40, or 80 Hertz for 1, 3, 5, or 7 hours, respectively.
Additionally, two mice were exposed at 1, 7, 20, 40, or 80 Hertz EMF for 7 hours, and then placed under ordinary breeding conditions for 24 hour, respectively.
After EMF exposure, the mice were sacrificed by cervical dislocation, and their abdomen immediately opened. Their abdominal temperature was measured with a digital therm- ometer (Summit, SDT62, Korea). Testes were removed, fixed in 10% neutral formalin, and embedded in paraffin. 4µm thick microsections were stained for apoptotic cells by using an in situ apoptosis detection kit (Oncor, USA). Briefly, the sections were deparaffinized and rehydrated by routine
procedures, and covered with 0.3% H2O2 for 10 minutes at room temperature in order to inactivate the endogenous peroxidase. The sections were then incubated with 1x equili- bration buffer (containing proteinase K) for 5 minutes at room temperature. After rinsing in PBS solution, the sections were applied with working strength terminal deoxynucleotidyl transferase (TdT) enzyme solution (containing digoxigenin labeled dUTP) in a humidified chamber at 37oC for 1 hour. The sections were washed in working strength stop/wash buffer for 10 minutes at room temperature and applied with peroxidase conjugated anti-digoxigenin antibody for 30 minutes at room temperature. The sections were incubated in peroxide substrate solution containing 0.01% diaminobenzidine tetrachloride (DAB, 0.1 mg/ml) and 0.02% hydrogen peroxide made in 0.1M Tris buffer, (pH 7.2) for 3-5 minutes, and washed in tap water for 5 minutes. The sections were counterstained, if necessary, and dehydrated in ethanol solution and mounted in Canada balsam.
Analysis
The apoptotic index (AI) was calculated as the percentage of in situ TdT positive cells among 2000 germ cells in sem- iniferous tubules. Mean and standard error of measurements of AI were analyzed in each group by frequency of the alternating EMF and the exposure time. Statistically the AI of experimental group was compared with that of sham treated control group by Student’s t-test.
Results
Electromagnetic property
The electric current induction in the secondary enamel coil by square-wave form directional current in the primary enamel coil of Magno-DR was characterized by alternating current in each frequency. Sharp initial spike was observed in 1-20 Hertz. When the frequency increased, the spike plateaued and almost disappeared in 30-90 Hertz (Fig. 3).
1 Hertz showed the highest initial spike (1.5 V), and up to 10 Hertz, the initial spike was still prominent (1.25-1.3 V).
From 20 Hertz the initial spike gradually decreased, then the initial spike almost disappeared and the current induction curve became plateaued (0.8-1.0V) in 30 to 90 Hertz. The total induced electric current per second increased rapidly until 6- 8 Hertz, and thereafter showed a slowly increasing curve until 30-40 Hertz and plateaued until 90 Hertz (Fig. 4). Therefore, 6-8 Hertz alternating EMF showed high efficiency of alternating EMF compared to 30-90 Hertz alternating EMF.
Biological effect on mouse testis
We explored the biological effect on testis by using different ELF-EMFs. After 1, 3, 5, 7 hours exposure of the ELF-EMF the mouse testis was examined with immunos- taining of PCNA and with in situ TdT staining methods. The proliferative index of negative control and sham treated
groups was about 20.4-25.1%, while that of ELF-EMF treated groups was much reduced, up to 12.4%. His- tologically as the ELF-EMF exposure increased until 7 hours, the layer of spermatocytes became deranged and produced immature spermatids. The number of proliferating spermatocytes detected by the immunostaining of PCNA was dramatically reduced in the ELF-EMF treated groups, and this anti-proliferative activity gradually increased as the ELF-EMF exposure was prolonged until 7 hours. Among the experimental groups the 7 Hertz EMF group showed the weakest reaction of PCNA in the mouse testis (Table 1, Fig.
1). On the other hands, the control testis showed rare positive reaction of in situ TdT staining (AI=0.3 ± 0.03%), and the AI of sham treatment group also showed minimum reaction (AI=0.2-0.4%) similar to the control group (Table 2, Fig. 2).
The alternating EMF induced a positive reaction of in situ TdT staining not only for the spermatogonia and primary sper- matocyte at the periphery but also for the secondary sper- matocyte and spermatid in the center of seminiferous tubules without inflammatory reaction (Fig. 4). When the exposure time increased, up to 5-7 hours, all experiment groups showed
frequent positive reaction of in situ TdT staining (Fig. 4).
Especially the 7 Hertz group showed a maximum increase of in situ TdT positivity in the spermatogonia (AI=4.7%, 4.6%, respectively) when compared to other groups. 5-7 hours exposure of 7 Hertz alternating EMF induced the maximum positive reaction of in situ TdT staining for the germ cells (Table 2, Fig. 2). However, by 24 hours rest after 7 hours exposure of different frequencies the mouse testis showed remarkable decrease of in situ TdT reaction (Fig. 4).
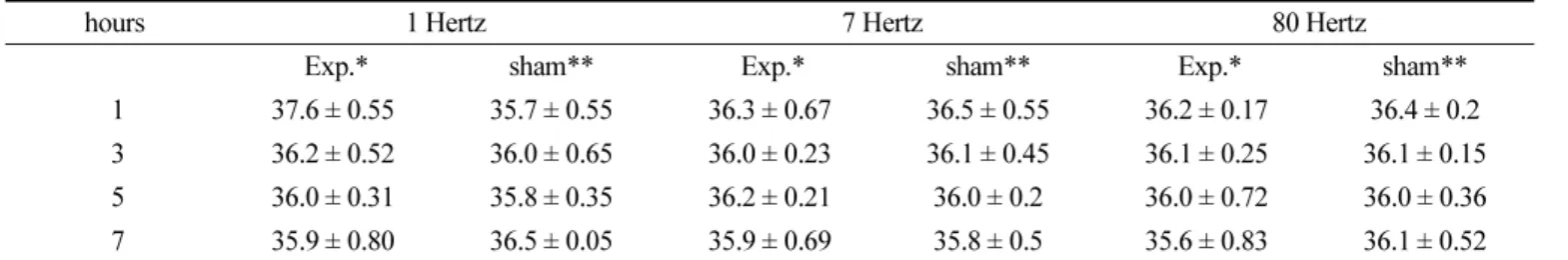
Additionally, the temperature of mouse abdominal cavity after scheduled EMF exposure in the chamber of the Magno-DR machine was not elevated throughout the whole experimental period compared to the normal condition of the sham treatment group (Table 3).
Discussion
In terms of weak ELF-EMF exposure, at 50-60 Hz, there Table 1. Mean and standard error of the PCNA index (PI, %) in each group of the ELF-EMF. PI represents the percentage of PCNA positive cells among 2000 germ cells in seminiferous tubules.
hours sham treat (n=10) 1 Hz (n=10) 7 Hz (n=10) 20 Hz (n=5) 40 Hz (n=5) 80 Hz (n=10) 1
3 5 7 7 (24h rest)
24.7 ± 2.33*
25.1 ± 1.51 22.4 ± 1.68 25.4 ± 1.32 23.5 ± 0.54
17.6 ± 0.54 15.7 ± 1.51 14.1 ± 0.74 12.7 ± 0.60 20.4 ± 1.47
15.1 ± 0.21 13.1 ± 1.25 13.7 ± 2.34 12.4 ± 1.24 22.4 ± 0.87
17.7 ± 1.11 15.1 ± 2.22 15.3 ± 1.43 14.1 ± 0.03 23.4 ± 2.02
17.2 ± 1.18 16.6 ± 2.24 16.1 ± 1.23 15.3 ± 1.31 22.4 ± 0.14
19.5 ± 1.29 19.7 ± 0.17 17.4 ± 2.27 18.1 ± 2.22 21.3 ± 2.02 normal control : 0.3 ± 0.03 (n=10), * p < 0.01
Fig. 1. Representative oscilloscope observations of the electric cur- rent induction at various frequencies, i.e., 1, 5, 7, 10, 30, 50, 90 Hertz. a: Applied voltage to the magnetic apparatus, b: Induced voltage by the pick-up coil in the chamber of magnetic apparatus, c: A magnetic apparatus used in this study, d: A pick-up coil.
Table 2. Mean and standard error of the apoptotic index (AI, %) in each group of the ELF-EMF. AI represents the percentage of in situ TdT positive cells among 2000 germ cells in seminiferous tubules.
Hours sham treat (n=10) 1 Hz (n=10) 7 Hz (n=10) 20 Hz (n=5) 40 Hz (n=5) 80 Hz (n=10) 1
3 5 7 7 (24h rest)
0.2 ± 0.05*
0.3 ± 0.02 0.2 ± 0.31 0.4 ± 0.25 0.4 ± 0.14
1.1 ± 0.14 1.5 ± 0.21 2.1 ± 0.32 2.8 ± 0.28 0.3 ± 0.22
1.8 ± 0.21 2.1 ± 0.25 4.7 ± 0.34 4.6 ± 0.24 0.4 ± 0.17
1.7 ± 0.11 2.1 ± 0.22 3.3 ± 0.43 4.1 ± 0.31 0.4 ± 0.12
1.2 ± 0.18 1.6 ± 0.24 2.1 ± 0.23 3.3 ± 0.31 0.4 ± 0.24
1.5 ± 0.29 1.7 ± 0.17 2.4 ± 0.27 3.1 ± 0.22 0.3 ± 0.22 normal control : 0.3 ± 0.03 (n=10), * p < 0.01
Fig. 2. Representative microscopic observations of seminiferous tubule treated by 7 Hertz EMF for 1, 3, 5, 7 hours. a1-a5: Hematox- ylin and eosin staining, the layer of spermatocytes was much de- ranged (arrows), b1-b5: PCNA immunostaining, some spermatogo- nias were negative (arrows), c1-c5: in situ TdT staining (×400), increased positive reaction in the experimental groups (arrows), C:
control, H: hour.
were already many negative results in human and animal pregnancy studies (Carratore et al., 1995; Li & Neutra, 2002;
Rogers et al., 1995; Savitz & Ananth, 1994; Sienkiewicz et
al., 1994; Svedenstael & Johanson, 1995), in physiological studies on enzymes and hormons (Carratore et al., 1995;
Grota et al., 1995; Lee et al., 1995; Rogers et al., 1995), and in Table 3. Changes of mouse abdominal temperature after of ELF-EMF exposure.
hours 1 Hertz 7 Hertz 80 Hertz
Exp.* sham** Exp.* sham** Exp.* sham**
1 37.6 ± 0.55 35.7 ± 0.55 36.3 ± 0.67 36.5 ± 0.55 36.2 ± 0.17 36.4 ± 0.2 3 36.2 ± 0.52 36.0 ± 0.65 36.0 ± 0.23 36.1 ± 0.45 36.1 ± 0.25 36.1 ± 0.15 5 36.0 ± 0.31 35.8 ± 0.35 36.2 ± 0.21 36.0 ± 0.2 36.0 ± 0.72 36.0 ± 0.36
7 35.9 ± 0.80 36.5 ± 0.05 35.9 ± 0.69 35.8 ± 0.5 35.6 ± 0.83 36.1 ± 0.52
normal control (n=10): 37.1 ± 0.74
*experimental group; n=5, **sham treatment
Fig. 3. Representative oscilloscope observations of the electric current induction at various frequencies, i.e., 1, 5, 7, 10, 30, 50, 90 Hertz. a:
Applied voltage to the magnetic apparatus, b: Induced voltage by the pick-up secondary coil in the chamber of magnetic apparatus, c: A mag- netic apparatus used in this study, d: A pick-up secondary coil.
animal behavior and cell culture studies (Malko et al., 1994;
Margonato et al., 1995; Orr et al., 1995; Rogers et al., 1995;
Spadinger et al., 1995), while only a few positive results have been reported using variable frequencies, different flux densities, and prolonged exposure (Guzelsu et al., 1994;
Lindstroem et al., 1995). In contrast to these studies using weak ELF-EMF, we used a strong ELF-EMF induced by square-wave pulsating current in this study. An important factor to be considered in the response of biological systems to a time-varying EMF is the waveform. Many different types of EMF waveform are used in practice, including sinusoidal, square-wave, sawtooth, and pulsed fields. For such fields, the two parameters of key importance are the rise and decay time of the signal, which determine the maximum time rate of
change of the field, (dB/dt), and the maximum instantaneous current density induced in tissues. An additional important parameter is tissue conductivity, which is frequency dependent and differs between tissues. In this study we used square-wave form directional current in different frequencies. The physical properties of the alternating EMF were analyzed and we observed that 6-8 Hertz alternating EMF showed high efficiency of alternating EMF compared to the 60-90 Hertz groups. Contrast to the high frequency alternating EMF, the low frequency alternating EMF produced an amplified current spike which may enhance the magneto-electric induction, and especially the square-wave form directional current can produce more acute inductance than the sinusoidal pulsating current. Thus, in this study we produced the alternating EMF Fig. 4. Representative microscopic observations of seminiferous tubule treated by 7 Hertz EMF for 1, 3, 5, 7 hours. a1-a5: Hematoxylin and eosin staining, the layer of spermatocytes was much deranged (arrows), b1-b5: PCNA immunostaining, some spermatogonias were negative (arrows), c1-c5: in situ TdT staining (× 400), increased positive reaction in the experimental groups (arrows), C: control, H: hour.
by square-wave pulsating current.
It is reported that exposure to 50 Hz ELF-EMF can produce apoptosis and small variations in cell cycle dis- tribution on different cell lines (Gluck et al., 2001; Johnson et al., 2001; Tofani et al., 2001). Others observed a gradual decrease in apoptosis and proliferation and a gradual increase in cells with a necrotic morphology after EMF exposure, and they suggested that these phenomena were variable depending on the cell type (Buemi et al., 2001). It is also known that weak EMF exposure results in protection from heat induced apoptosis in human cancer cell lines in a time dependent manner (Robison et al., 2002). However, cell- death induction was observed with EMF of more than 1 mT, which was independent of the EMF frequency and the cell death increased when modulated EMF (static with a superimposition of ELF-EMF at 50 Hz) was used (Tofani et al., 2001). The presence of dexamethasone or X-ray irradia- tion during the EMF exposure could probably produce a de- crease in the cell growth or a increase of apoptosis, respectively (Ruiz Gomez et al., 2001).
The testis is a reproductive organ showing active prolif- eration and continuous germ cell differentiation (De Kretser &
Kerr, 1988). Seminiferous tubules are composed of somatic cells (myoid cells and Sertoli cells), and germ cells (spermat- ogonia, spermatocytes, and spermatids). Activity of these germ cells divides spermatogenesis into spermatocytogenesis, meiosis, and spermiogenesis, while germ cell degeneration occurs throughout spermatogenesis (Johnson, 1995). Many kinds of toxic chemicals and irradiation can produce harmful effects on the germ cells of testis (De Vita et al., 1995). It is also suggested that the accumulation of alternating free radicals by long-term exposure of alternating EMF may be harmful to the germ cells in the processes of sper- matocytogenesis and meiosis (Smith et al., 1986; Smith et al., 1984). Additionally, the exposure of tissue to alternating EMF might elevate body temperature, and such an elevation of temperature, if it occurred, could also be harmful to the germ cells of testes. In this study Magno-DR machine had an effective cooling system to prevent any elevation of abdominal temperature in mouse during exposure. Thus we thought thermal factor did not affect the animal during the EMF exposure.
We observed an increase of apoptosis in the testes after exposure to a strong ELF-EMF, about 730-960 Gauss. The in situ TdT staining was positive in the spermatogonia and primary spermatocytes within the first hour of exposure.
With prolonged exposure, 5-7 hours, the TdT staining became frequently positive even in the secondary spermatocytes and spermatid. Although the development of particular type of germ cells is synchronized around a radial axis from the center to the periphery of the tubules, the mouse testis could be classified into 12 stages of spermatogenesis through the seminiferous tubules in helical arrangement (Clermont, 1972). The multiple observation of tubules indicated that the peripheral germ cells, including spermatogonia and primary
spermatocytes, were more easily affected by the square alternating EMF than the central germ cells composed of secondary spermatocytes and spermatid. This may imply that the alternating EMF produces a harmful effect on the rapidly proliferating germ cells. On the other hands, by 24 hour rest after 7 hours exposure of the EMF the germ cells became negative for in situ TdT staining. Thus we presumed that the damaged germ cells could be recovered during 24 hours rest in normal condition. Although this study cannot explain the precise mechanism of the hazardous effect of the alternating EMF, it is suggested that the strong alternating EMF induces apoptotic changes in the germ cells of the mouse testis.
The present study showed that the alternating EMF is harmful depending on its frequency. The harmful effect was most efficient in 6-8 Hertz, while it became much reduced in 30-90 Hertz. As the 6-8 Hertz has a time interval, 0.125- 0.167 sec, similar to the T2 relapse time of magnetic proton reaction by 800 Gauss, about 0.15 sec. Therefore, we thought that the harmful effect of alternating EMF is related to the magnetic proton reaction rather than the inductance of electric current. One of possible basic mechanism is the forced-vibration of all the free ions on the surface of plasma membrane of the cell, caused by an external oscillating EMF.
This coherent vibration of electric charge is able to irregularly gate electrosensitive channels on the plasma membrane and thus cause disruption of the cell's electrochemical balance and function, leading to cellular apoptosis (Panagopoulos et al., 2002; Panagopoulos et al., 2000). However, although further study should be followed to elucidate the mechanism of various biological effects of ELF-EMF, in this study we have observed that the 7 Hertz of alternating EMF induced most harmful effect on the germ cells of mouse testis compared to the higher frequency groups.
References
Adey WR. Frequency and power windowing in tissue inter- actions with weak electromagnetic fields. Proceedings of the Institute of Electrical and Electronics Engineers. 1980;68:
119-25.
Ayrapetyan SN, Grigorian KV, Avanesian ASStamboltsian KV. Magnetic fields alter electrical properties of solutions and their physiological effects. Bioelectromagnetics. 1994;
15:133-42.
Bassett CA. The development and application of pulsed electromagnetic fields (PEMFs) for ununited fractures and arthrodeses. Orthop Clin North Am. 1984;15:61-87.
Bawin SM, Adey WRSabbot IM. Ionic factors in release of 45Ca2+ from chicken cerebral tissue by electromagnetic fields. Proc Natl Acad Sci USA. 1978;75:6314-8.
Blackman CF, Benane SG, Elder JA, House DE, Lampe JAFauk JM. Induction of calcium-ion efflux from brain tissue by radio-frequency radiation: effects of modulation frequency and field strength. Radio Science. 1979;14:93-8.
Blackman CF, Benane SG, House DEJoines WT. Effects of ELF (1-120 Hz) and modulated (50 Hz) RF fields on the efflux of calcium ions from brain tissue in vitro. Bioelec- tromagnetics. 1985;6:1-11.
Buemi M, Marino D, Di Pasquale G, Floccari F, Senatore M, Aloisi C, Grasso F, Mondio G, Perillo P, Frisina NCorica F.
Cell proliferation/cell death balance in renal cell cultures after exposure to a static magnetic field. Nephron. 2001;87:
269-73.
Carratore RD, Morichetti E, Croce CDBronzetti G. Effect of magnetic fields on rodent monooxygenase enzymes. Bioelec- tromagnetics. 1995;16:324-9.
Cavol AV, Wamil AW, Holcomb RRMclean KJ. Measurement and analysis of static magnetic fields that block action potentials in cultures neurons. Bioelectomagnetics. 1995;16:
197-206.
Chiang H, Wu RY, Shao BJ, Fu YD, Yao GDLu DJ. Pulsed magnetic field from video display terminals enhances teratogenic effects of cytosine arabinoside in mice. Bioelec- tromagnetics. 1995;16:70-4.
Clermont Y. Kinetics of spermatogenesis in mammals: Sem- iniferous epithelium cycle and spermatogonial renewal.
Physiol Rev. 1972;52:198-236.
De Kretser DMKerr JB. The cytology of testis. Raven Press, New York 1988.
De Vita R, Cavallo D, Raganella L, Eleuteri P, Grollino MGCalugi A. Effect of 50 Hz magnetic fields on mouse spermatogenesis monitored by flow cytometric analysis.
Bioelectromagnetics. 1995;16:330-4.
Diniz P, Shomura K, Soejima KIto G. Effects of pulsed electromagnetic field (PEMF) stimulation on bone tissue like formation are dependent on the maturation stages of the osteoblasts. Bioelectromagnetics. 2002;23:398-405.
Engstroem S. Dynamic properties of Lednev's parametric res- onance mechanism. Bioelectromagnetics. 1996;17:58-70.
Gluck B, Guntzschel VBerg H. Inhibition of proliferation of human lymphoma cells U937 by a 50 Hz electromagnetic field. Cell Mol Biol (Noisy-le-grand). 2001;47:OL115-7.
Grant G, Cadossi RSteinberg G. Protection against focal cerebral ischemia following exposure to a pulsed electromagnetic field. Bioelectromagnetics. 1994;15:205-16.
Grota LJ, Reiter RJ, Keng PMichaelson S. Electric field exposure alters serum melatonin but not pineal melatonin synthesis in male rats. Bioelectromagnetics. 1995;15:4237.
Guzelsu N, Salkind AJ, Shen X, Patel U, Thaler SBerg RA.
Effect of electromagnetic stimulation with different waveforms on cultured chick tendon fibroblasts. Bioelectromagnetics.
1994;15:115-31.
Inoue N, Ohnishi I, Chen D, Deitz LW, Schwardt JDChao EY.
Effect of pulsed electromagnetic fields (PEMF) on late-phase osteotomy gap healing in a canine tibial model. J Orthop Res.
2002;20:1106-14.
Johnson L. Efficiency of spermatogenesis. Microscopy Research and Technique. 1995;32:385-422.
Johnson MT, Vanscoy-Cornett A, Vesper DN, Swez JA, Chamberlain JK, Seaward MBNindl G. Electromagnetic fields used clinically to improve bone healing also impact lymphocyte proliferation in vitro. Biomed Sci Instrum.
2001;37:215-20.
Kirschvink JL, Kobayashi-Kirschvink A, Diaz-Ricci JCKirschvink SJ. Magnetite in human tissues: a mechanism for the biological effects of weak ELF magnetic fields. Bioelectromagnetics (Suppl). 1992;1:101-13.
Ko SH, Lee JHKim SH. The Effects of Mechanical Strain on Bone Cell Proliferation and Recruitment Induced by Osteocytes. Int J Oral Biology. 2008;33:179-86.
Kolosova LI, Akoev GN, Avelev VD, Riabchikova OVBaba KS. Effect of low-intensity millimeter wave electromagnetic radiation on regeneration of the sciatic nerve in rats.
Bioelectromagnetics. 1996;17:44-7.
Lee JM, Stormshak F, Thompson JM, Hess DLFoster DL.
Melatonin and puberty in female lambs exposed to EMF: A replicate study. Bioelectromagnetics. 1995;16:119-23.
Lee SK, Chung EY, Kim GJ, Song DB, Kim JHChi JG. An effective role of pulsed magnetic field for bony decalci- fication. Kor J Pathol. 1993;27:125-33.
Li DKNeutra RR. Magnetic fields and miscarriage.
Epidemiology. 2002;13:237-8.
Lindstroem E, Lindstroem P, Berglund A, Lundgren EMild KH. Intracellular calcium oscillation in a T-cell line after exposure to extremely-low-frequency magnetic fields with variable frequencies and flux densities. Bioelectromag- netics. 1995;16:41-7.
Malko JA, Constantinidis I, Dillehay DFajman WA. Search for influence of 1.5 Tesla magnetic field on growth of yeast cells. Bioelectromagnetics. 1994;15:495-501.
Margonato V, Nicolini P, Conti R, Zecca L, Veicsteinas ACerretelli P. Biologic effects of prolonged exposure to ELF electromagnetic fields in rats: II. 50 Hz magnetic fields.
Bioelectromagnetics. 1995;16:343-55.
Orr JL, Rogers WRSmith HD. Exposure of baboons to combined 60 Hz electric and magnetic fields does not produce work stoppage or affect operant performance on a match-to- sample task. Bioelectromagnetics. 1995;3:61-70.
Panagopoulos D, Karabarbounis AMargaritis L. Mechanism for action of electromagnetic fields on cells. Biochem Biophys Res Commun. 2002;298:95.
Panagopoulos DJ, Messini N, Karabarbounis A, Philippetis ALMargaritis LH. A mechanism for action of oscillating electric fields on cells. Biochem Biophys Res Commun. 2000;
272:634-40.
Parafiniuk M, Gorczynska E, Gutsch AParafiniuk W. Effect of constant magnetic field on the liver of guinea pig. Electron microscopic studies. Folia Histochem Cytobiol. 1992;30:
119-23.
Portet RTCabanes J. Development of young rats and rabbits exposed to strong electric field. Bioelectromagnetics.
1988;9:95-104.
Robison JG, Pendleton AR, Monson KO, Murray BKO'Neill KL. Decreased DNA repair rates and protection from heat induced apoptosis mediated by electromagnetic field exposure. Bioelectromagnetics. 2002;23:106-12.
Rogers WR, Orr JLSmith HD. Nonhuman primates will not respond to turn off strong 60 Hz electric fields. Bioelectro- magnetics. 1995;3:48-60.
Rogers WR, Reiter RJ, Barlow-Walden L, Smith HDOrr JL.
Regularly scheduled, day-time, slow onset 60 Hz electric and magnetic field exposure does not depress serum mela- tonin concentration in nonhuman primates. Bioelectromag- netics, Supplement. 1995;3:111-8.
Ruiz Gomez MJ, De la Pena L, Pastor JM, Martinez Morillo MGil L. 25 Hz electromagnetic field exposure has no effect on cell cycle distribution and apoptosis in U-937 and HCA- 2/1cch cells. Bioelectrochemistry. 2001;53:137-40.
Saunders RDJefferys JG. Weak electric field interactions in the central nervous system. Health Phys. 2002;83:366-75.
Savitz DAAnanth CV. Residential magnetic fields, wire codes, and pregnancy outcome. Bioelectromagnetics. 1994;15:271-3.
Sienkiewicz Z, Robbins L, Haylock RGESaunders RD. Effects of prenatal exposure to 50 Hz magnetic fields on development in mice: II. Postnatal development and behavior. Bioelectro- magnetics. 1994;15:363-75.
Smith A, Bugge HP, Berg KA, Moller OHansson V. Seasonal changes in testicular structure and function in the blue fox (Alopex lagopus), as quantified by morphometric analysis and measurement of adenylate cyclase activity. Int J Androl.
1986;9:53-66.
Smith AJ, Clausen OP, Kirkhus B, Jahnsen T, Moller OMHansson V. Seasonal changes in spermatogenesis in the blue fox (Alopex lagopus), quantified by DNA flow cytometry and measurement of soluble Mn2 -dependent adenylate cyclase activity. J Reprod Fertil. 1984;72:453-61.
Spadinger I, Agnew DPalcic B. 3T3 cell motility and mor- phology before, during, and after exposure to extremely-low- frequency magnetic fields. Bioelectromagnetics. 1995;16:178- 87.
Svedenstael B-MJohanson K-J. Fetal loss in mice exposed to magnetic fields during early pregnancy. Bioelectromagnetics.
1995;16:284-9.
Takano-Yamamoto T, Kawakami MSakuda M. Effect of a pulsing electromagnetic field on demineralized bone-matrix- induced bone formation in a bony defect in the premaxilla of rats. J Dent Res. 1992;71:1920-5.
Tofani S, Barone D, Cintorino M, de Santi MM, Ferrara A, Orlassino R, Ossola P, Peroglio F, Rolfo KRonchetto F. Static and ELF magnetic fields induce tumor growth inhibition and apoptosis. Bioelectromagnetics. 2001;22:419-28.