한수지 54(3), 280-286, 2021
280
Copyright © 2021 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815 Korean J Fish Aquat Sci 54(3),280-286,2021
Original Article
서 론
생물들은외부미생물의침입으로부터자신을방어하기위한 효과적인 면역체계로서선천면역계 또는선천면역계와 후천
면역계를동시에함유하고있는것으로알려져있다
(Hancock
and Diamond, 2000).
선천면역계는외부의위협으로부터물 리적인장벽역할을담당하는피부/
점막등과항균물질,
보체,
히스타민등과같은화학적장벽역할을담당하는체액성물질 들및탐식세포들에의한식균작용또는염증반응과같은반응 성요소등으로구성되며비특이적1
차방어작용을담당한다(Clark and Kupper, 2005; Chung, 2019).
반면후천면역계는 항원특이적항체생산에의한체액성면역과세포독성T
세포에의한세포성면역등으로구성되며특이적
2
차방어작용을 담당한다.
이러한선천면역과후천면역은상호협력과조화를 통해서효과적인생체방어체계를구성/
유지하고있으나면역 계의구성은척추동물과무척추동물에서차이가있다고알려져 있다(Seo, 2016).
선천면역과후천면역이모두잘발달되어있 는척추동물과는달리선천면역만을포함하고있는무척추동물 은자기방어작용을선천면역에만의존한다고알려져있다(Seo et al., 2005; Rowley and Powell, 2007).
최근들어서는비특이적이면서빠른반응성을나타내는선천 면역의기능과그구성요소들에 대한관심이크게증대되고 있으며
,
특히,
해양무척추동물들에존재하는선천면역성분들 과그활용에대한연구가활발히수행되고있다(Zasloff, 2002;
진주담치(Mytilus edulis) 추출물의 항균활성 및 단백질 분해효소에 대한 안정성 탐색
이지은·서정길*
국립군산대학교 해양과학대학 식품생명공학전공
Screening of Antimicrobial Activity and Proteolytic Enzyme Stability of Extract of the Blue Mussel Mytilus edulis
Ji-Eun Lee and Jung-Kil Seo*
Department of Food Science and Biotechnology, Kunsan National University, Kunsan 54150, Korea
This study was performed to screen the antimicrobial activities and proteolytic enzyme stability of the acidified extract of the Blue mussel Mytilus edulis . The acidified extract showed potent antimicrobial activities against Gram- positive bacteria, Bacillus subtilis , and Gram-negative bacteria, Escherichia coli D31, but had no activity against Candida albicans . Treatment of extract with trypsin completely abolished all or significant antibacterial activity against the tested bacteria, but slightly decreased antimicrobial activity against B. subtilis , and treatment of extract with chymotrypsin retained almost antibacterial activity against the tested bacteria except for E. coli D31. To confirm the additional enzyme stability of the extract, antimicrobial activity of the extract was tested after treated with sev- eral enzymes. Enzymes treated extract showed potent antimicrobial activity against B. subtilis and its activity was also retained for 5 h after trypsin treatments. Non-proteinaceous materials in the acidified extract also showed strong DNA-binding ability but did not show bacterial membrane permeabilizing ability. All our results indicate that mussel extract might contain the proteinaceous or non-proteinaceous antibacterial materials target not bacterial membrane but intracellular components. These results could be used to develop mussel extract as an additive for the improve- ment of stability or antimicrobial activity of antibiotics against specific bacteria.
Keywords: Mytilus edulis , Extract, Antimicrobial activity, Enzyme stability
*Corresponding author: Tel: +82. 63. 469. 1827 Fax: +82. 63. 469. 7448 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 15 March 2021; Revised 13 April 2021; Accepted 23 April 2021 저자 직위: 이지은(대학원생), 서정길(교수)
https://doi.org/10.5657/KFAS.2021.0280
Korean J Fish Aquat Sci 54(3), 280-286, June 2021
진주담치(Mytilus edulis) 추출물의 항균활성 및 효소 안정성 탐색
281
Tom et al., 2011).
해양무척추동물은혈구세포(hemocytes)
의 탐식작용과이들세포로부터생성/
분비되는다양한물질들의 효과적인작용에의해1
차방어기능이수행되고있으며,
대표 적인구성요소로서lectin, pro-phenoloxidase activating factor, antimicrobial peptide
및phagocytosis
와inflammation
등이잘 알려져있다(Relf et al., 1999; Tincu and Taylor, 2004; Yu et al., 2005; Wang et al., 2014).
현재까지밝혀진해양무척추동물 의선천면역관련물질들중에서단백질성항균물질의직접적 이고효과적인방어작용이알려지면서,
이것들이포함된해양 무척추동물추출물들의면역학적활용을위한연구가활발히 진행되고있다(van’'t Hof et al., 2001).
그러나해양무척추동물 추출물에포함된단백질성항균물질은단백질분해효소에대한 민감성이매우높다는것이확인되었고이에따라이것들의활용을위한안정성개선에대한연구가필요한실정이다
(Lee et
al., 1997).
따라서추출물내에포함된단백질성및비단백질성 항균물질의존재유무와단백질성항균물질을보조할수있는 비단백질성항균에대한탐색과활용가능성에대한연구가선 행되어야할분야로인식되고있다(Kim et al., 2013).
해양무척추동물중진주담치와같이부착생활을하는무척추 동물은운동성
/
회피성의부재로다양한해양환경의변화나오 염원또는병원성인자들에대한효과적인방어를할수없으므 로환경인자에효과적인방어작용을담당하는선천면역체계/
요소들을포함하고있을가능성이매우높다.
특히,
우리나라해 안의조간대주변에서가장많이발견되는대표적인부착성해 양무척추동물중의하나인진주담치(Mytilus edulis)
는연체동 물부족류홍합목홍합과에속하며환경적응력과번식력에서 큰주목을받고있는산업적가치가높은패류중의하나로알려 져있다.
현재까지Mytilus edulis (Charlet et al., 1996), Mytilus galloprovincialis (Hubert et al., 1996; Mitta et al., 1999)
및Mytilus coruscus (Oh et al., 2018)
등의홍합또는진주담치추출물들이포함하고있는
defensins
류를포함한단백질성항균물질의면역학적역할에대한연구는활발하게진행되었지만 단백질분해효소에대한민감성등을개선하기위한효소저항 성항균물질에대한연구는부족한실정이다
.
따라서본연구에서는효소저항성항균물질에대한연구가 비교적부족한진주담치추출물의항균활성과단백질분해효 소에대한안정성을탐색함으로써진주담치추출물의항균활 성분야에서의활용가능성을고찰하고자한다
.
재료 및 방법
실험 재료 및 시약
본실험에사용한진주담치
(M. edulis)
는경상남도고성만에 서구입하였으며살아있는상태로실험실로운반된후에아가 미와외투막조직을적출한즉시추출과정에사용하였다.
항균활성 측정을 위한 배지성분으로서
tryptic soy broth
(TSB)
와sabouraud dextrose broth (SDB)
및agarose type I (Low EEO Agar)
는Merck
사(Merck, Darmstadt, Germany)
와Sigma
사(St. Louis, MO, USA)
에서각각구입하여사용하였 다.
효소처리를위해서trypsin
과chymotrypsin
은Fisher Sci- entific (Fairlawn, NJ, USA)
사에서구입을하였으며, protein- ase K
와α-Amylase
및α-Lipase
는Sigma
사(St. Louis, MO, USA)
에서각각구입하여사용하였다.
세균내막투과성을측 정하기 위해서 사용된o-nitrophenyl-β-D-galactopyranoside (ONPG)
는Sigma
사(St. Louis, MO, USA)
에서 구입하였으 며, DNA-binding ability
측정에사용된100 bp DNA ladder
는Bioneer
사(Daejeon, Korea)
에서구입하여사용하였다.
그 리고연구에사용된그외의시약들은모두특급을사용하였다. 조직추출
실험실로운반된진주담치는흐르는수돗물에씻어서패각에 부착된오염물을제거한후
,
아가미와외투막조직을취하여5
mL volume
이될때까지얼음속에서모았다.
모아진아가미와외투막조직에끓고있는
4
배량의1% acetic acid (v/v)
를 첨가하여100°C
에서5
분동안끓인후,
얼음에보관하여충분 히냉각시켰다.
충분히냉각된조직은homogenizer (T10 basic ULTRA-TURRAX; IKA, Wilmington, NC, USA)
를사용하 여완전히파쇄시켰다(Speed #6, 3
분,
실온).
조직파쇄액은4°C
에서20
분동안8,000 rpm
으로원심분리(VS-21SMT; Vision Scientific, Buchon, Korea)
를행한후에상층액을취해서항균 활성탐색과정등의연구에사용될때까지-70
oC
에보관하였다(Seo et al., 2005).
항균활성 측정방법 및 사용 균주
추출물들의항균활성측정을위해서그람양성균
3
종(Bacillus subtilis KCTC1021, Staphylococcus epidermidis KCTC1917, Streptococcus mutans KCCM4105),
그람음성균5
종(Esch- erichia coli D31, Escherichia coli ML35p, Cutibacterium acnes (Propionibacterium acnes) KCTC11946, Shigella flex- neri KCTC2009, Vibrio parahaemolyticus RIMD2110633)
및yeast 1
종(Candida albicans KCTC7965)
을사용하였다.
항균 활성측정방법으로는서로다른농도를포함한두층의배지 를 사용하는ultrasensitive radial diffusion assay (URDA)
법 을이용하였다(Lehrer at al., 1991).
항균활성측정에사용된세 균은tryptic soy broth (TSB)
에접종하고, C. albicans
은sab- ouraud dextrose broth (SDB)
에접종한후18
시간동안37°C (V. parahaemolyticus
는25°C)
에서pre-culture
를 수행한 후colorimeter (Product No. 52-1210; BioMerieux Inc., Durham, NC, USA)
를사용하여균농도를84%T (≒1×10
8CFU/mL) (C. albicans
의경우는10
6CFU/mL)
가되도록조정하였다. C.
acnes
은RCM
배지에접종한후anaerobic jar
에서Gaspack
system (BBL Gaspack 100TM; Franklin Lakes, NJ, USA)
을 이용하여밀봉한후37°C
에서3
일동안혐기배양후에동일한이지은
ㆍ
서정길282
방법으로균농도를맞추었다
.
그후, 9.5 mL
의0.03% TSB (
또는0.03% SDB), 1% type I agarose
및10 mM phosphate buffer (PB) (pH 6.5)
를포함하는underlay gel
에각각의농도로 희석된균액0.5 mL
을첨가하여잘섞은후에plate
에편평하게 부어굳혔다.
이때E. coli D31
균주(streptomycin
내성균)
를사 용할경우에는streptomycin
을10 μg/mL
농도로첨가하여잘 섞은후에plate
에편평하게부어굳혔다.
굳은plate
에punch
를 사용하여직경2.5 mm
의well
을뚫은후에5 μL
의각추출물 을도입시켰다.
추출물이배지에완전히스며들도록3
시간동 안37°C
에서1
차배양한후,
그위에10 mL
의6% TSB (
또는6% SDB), 1% type I agarose
및10 mM phosphate buffer (pH 6.5)
를포함하는overlay gel
을붓고굳힌후에동일한온도에 서18
시간동안2
차배양하였다(Seo et al., 2005).
다음날well
주위에생긴
clear zone
의크기를측정하여항균활성을확인하였다
.
항균활성측정동안positive control
로는미국산잡종농 어(Morone saxatilis×Morone chrysops)
의mast cell
에서정제 된항균펩타이드인piscidin 1
을사용하였고, negative control
로는1% acetic acid
또는0.01% acetic acid
를사용하였다(Sil- phaduang and Noga, 2001). Piscidin 1
은㈜
펩트론(Peptron, Daegeon, Korea)
에서합성및95%
이상의순도로정제된것을 구입하여사용하였다.
Enzyme 처리에 의한 항균물질의 특성 확인
추출물에포함된항균물질의단백질성및지질이나탄수화물 성을확인하기위해서단백질분해효소인
trypsin, chymotryp- sin, proteinase K
등과α-amylase, α- lipase
등의처리전·
후의 항균활성변화를확인하였다.
이를위하여진주담치추출물5 μL
에enzyme
용액(1,000 μg/mL in 50 mM PB, pH 7.4) 1 μL
를첨가하고37°C
에서60
분간반응시킨뒤, URDA
법으로B.
subtilis KCTC1021
과E. coli D31
에대한항균활성을측정하 였다(Seo et al., 2017).
추출물의 세포독성 확인
진주담치 추출물의
human dermal fibroblasts (HDF)
에 대 한세포독성을확인하기위해서MTT assay
를 이용하여cell viability
를측정하였다(Kim and Li, 2012).
이를위해서다양 한농도의진주담치추출물(0, 0.5, 1.0, 10 mg/mL)
을human dermal fibroblasts cell
에처리한후추출물의농도에따른세포 생존율을확인하였다.
추출물들의 cytoplasmic membrane permeabilization assay
진주담치 추출물의 세균 내막 투과성 측정을 위해서
β-galactosidase
활성을포함하는E. coli ML35p
를사용해서nonmembrane-permeative chromogenic
기질인o-nitrophenyl- β-D-galactopyranoside (ONPG)
에대한추출물도입후의E.
coli ML35p
의세포질에서유출된β-galactosidase
의활성을확인하였다
(Skerlavaj et al., 1990).
배양된mid-log phase
의E.
coli ML35p
를10 mM sodium phosphate buffer (pH 7.4)
로세 척을한후, 1.5 mM
의ONPG
를포함하는동일butter
에용해시 켰다.
그후,
측정할추출물들을첨가한뒤37°C
에서60
분간배 양하면서10
분간격으로유출된β-galactosidase
에의한ONPG
의o-nitrophenol
로의가수분해정도를405 nm
에서측정하였 다.
막투과성측정의표준물질로는강한막투과성을나타내는 것으로알려진항균펩타이드인piscidin 1
을사용하였다. DNA Binding Assay
진주담치추출물에존재하는
DNA
결합성물질의존재유무를확인하기위해서
DNA binding
에의한DNA band
들의aga- rose gel-electrophoresis
에서의이동의저해정도를확인하는DNA-binding assay
를수행하였다(Hsu et al., 2005).
이를위 해서100 bp DNA ladder (0.2 ng)
와추출물을혼합해서37°C
에서60
분동안반응시키고0.5 μg/mL EtBr
을포함하는1.4%
agarose gel
에서전기영동을수행한후DNA band
의유무와이 동정도를확인함으로써DNA-binding
유무를확인하였다.
결과 및 고찰
진주담치 추출물의 항균활성
진주담치
(M. edulis)
로부터 채취한 아가미(gill)
와 외투막(mantle)
조직들은1% HAc (
초산)
를이용한약산 추출과정 을통해서추출되었으며,
각추출물의항균활성은URDA
법을 이용하여B. subtilis KCTC1021, E. coli D31
및C. albicans KCTC7965
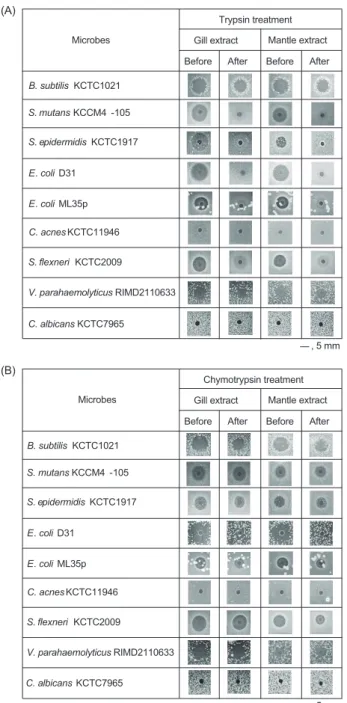
에대해서측정하였다(Fig. 1).
실험결과,
각추출 Fig. 1. Antimicrobial activity of the acidified crude extracts of the blue mussel Mytilus edulis. Antimicrobial activity of the acidified gill or mantle extract against Bacillus subtilis KCTC1021, Esch- erichia coli D31 and Candida albicans KCTC7965. Scale bar in- dicates 5 mm.Bacillus subtilis KCTC1021
Escherichina coli D31
Gill Mantle
Candida albicans KCTC7965
, 5 mm
(A)
B. subtilis KCTC1021 S. mutans KCCM4 -105 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After Trypsin treatment
Before After Gill extract Mantle extract
, 5 mm Microbes
(B)
B. subtilis KCTC1021 S. mutans KCCM4 -105
C. albicans KCTC7965 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After
Chymotrypsin treatment Before After Gill extract Mantle extract
, 5 mm Microbes
B. subtilis Gill
Mantle
T C A L P N
, 5 mm
B. subtilis Gill
Mantle
0 h 1 h 2 h 3 h 4 h 5 h
, 5 mm
0 0.2 0.4 0.6 0.8 1
0 10 20 30 40 50 60
Optical Density (O.D.) 405
Time (min)
Trypsin-treated Gill extract Proteinase K-treated Gill extract Piscidin 1
C. albicans KCTC7965
진주담치(Mytilus edulis) 추출물의 항균활성 및 효소 안정성 탐색
283
물은
B. subtilis
와E. coli D31
에대해서는강한항균활성을나 타낸반면에C. albicans
에대해서는항균활성을나타내지않았다
.
이러한결과는진주담치추출물의항균활성범위는세균에 한정되어있을가능성을의미하는것이다.
각추출물내의
positive charge
아미노산(Lys
과Arg)
과방향 족아미노산(Trp, Phe, Tyr)
을포함하는선천면역관련단백질 성항균물질의존재유무를확인하기위하여각추출물(5 μL)
의trypsin
및chymotrypsin
처리전∙
후의항균활성변화를URDA
법을이용하여B. subtilis
및E. coli D31
를포함하는9
종의세 균과C. albicans
에대해서측정하였다(Fig. 2).
측정결과,
추출 물들은trypsin
처리후에는B. subtilis
와V. parahaemolyticus
에대한항균활성은거의유지가된반면에E. coli D31
을포함 하는대부분의세균들에대한항균활성은거의소실되었다.
또 한,
추출물들은chymotrypsin
처리후에는E. coli D31
을제외 한대부분의세균들에대한항균활성은거의유지되었다.
그리 고추출물또는균주간의활성도비교에서는아가미추출물 과외투막추출물은그람음성균과그람양성균에대해서유사 한항균활성을나타냈지만C. acnes
와C. albicans
에대해서는 항균활성이없는것으로나타났다.
이러한결과는진주담치추 출물에포함된항균물질은chymotrypsin
보다는trypsin
에더 민감한반응을보이는물질임을의미하는것으로서아르기닌(Arg)
또는라이신(Lys)
과같은양이온성아미노산을포함하는단백질성물질일가능성이있음을의미하는것이다
.
또한B.
Fig. 2. Antimicrobial activity of the acidified crude extracts of the blue mussel Mytilus edulis. Antimicrobial activity of the acidified gill or mantle extract (before) and trypsin (A) or chymotrypsin (B) treated extract (after) against Gram-positive bacteria including Ba- cillus subtilis, Streptococcus mutans, Staphylococcus epidermidis and Gram-negative bacteria including Escherichia coli D31, Cuti- bacterium acnes, Shigella flexneri, Vibrio parahaemolyticus, and Candida albicans. Scale bar indicates 5 mm.
Bacillus subtilis KCTC1021
Escherichina coli D31
Gill Mantle
Candida albicans KCTC7965
, 5 mm
(A)
B. subtilis KCTC1021 S. mutans KCCM4 -105 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After Trypsin treatment
Before After Gill extract Mantle extract
, 5 mm Microbes
(B)
B. subtilis KCTC1021 S. mutans KCCM4 -105
C. albicans KCTC7965 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After
Chymotrypsin treatment Before After Gill extract Mantle extract
, 5 mm Microbes
B. subtilis Gill
Mantle
T C A L P N
, 5 mm
B. subtilis Gill
Mantle
0 h 1 h 2 h 3 h 4 h 5 h
, 5 mm
0 0.2 0.4 0.6 0.8 1
0 10 20 30 40 50 60
Optical Density (O.D.) 405
Time (min)
Trypsin-treated Gill extract Proteinase K-treated Gill extract Piscidin 1
C. albicans KCTC7965
Fig. 3. Antimicrobial activity of the enzyme-treated crude extracts of the blue mussel Mytilus edulis. Antimicrobial activity of gill and mantle extract after treated several enzymes including trypsin (T), chymotrypsin (C), α-amylase (A), α-lipase (L), proteinase K (P) or not treated (N) against Bacillus subtilis. Scale bar indicates 5 mm.
Bacillus subtilis KCTC1021
Escherichina coli D31
Gill Mantle
Candida albicans KCTC7965
, 5 mm
(A)
B. subtilis KCTC1021 S. mutans KCCM4 -105 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After Trypsin treatment
Before After Gill extract Mantle extract
, 5 mm Microbes
(B)
B. subtilis KCTC1021 S. mutans KCCM4 -105
C. albicans KCTC7965 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After
Chymotrypsin treatment Before After Gill extract Mantle extract
, 5 mm Microbes
B. subtilis Gill
Mantle
T C A L P N
, 5 mm
B. subtilis Gill
Mantle
0 h 1 h 2 h 3 h 4 h 5 h
, 5 mm
0 0.2 0.4 0.6 0.8 1
0 10 20 30 40 50 60
Optical Density (O.D.) 405
Time (min)
Trypsin-treated Gill extract Proteinase K-treated Gill extract Piscidin 1
C. albicans KCTC7965
Fig. 4. Antimicrobial activity of the trypsin-treated crude extracts of the blue mussel Mytilus edulis. Antimicrobial activity of gill or mantle extract after treated with trypsin for 5 h against Bacillus subtilis KCTC1021. Scale bar indicates 5 mm.
Bacillus subtilis KCTC1021
Escherichina coli D31
Gill Mantle
Candida albicans KCTC7965
, 5 mm
(A)
B. subtilis KCTC1021 S. mutans KCCM4 -105 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After Trypsin treatment
Before After Gill extract Mantle extract
, 5 mm Microbes
(B)
B. subtilis KCTC1021 S. mutans KCCM4 -105
C. albicans KCTC7965 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After
Chymotrypsin treatment Before After Gill extract Mantle extract
, 5 mm Microbes
B. subtilis Gill
Mantle
T C A L P N
, 5 mm
B. subtilis Gill
Mantle
0 h 1 h 2 h 3 h 4 h 5 h
, 5 mm
0 0.2 0.4 0.6 0.8 1
0 10 20 30 40 50 60
Optical Density (O.D.) 405
Time (min)
Trypsin-treated Gill extract Proteinase K-treated Gill extract Piscidin 1
C. albicans KCTC7965
이지은
ㆍ
서정길284
subtilis
와V. parahaemolyticus
에대한진주담치추출물들의항 균활성은trypsin
과chymotrypsin
처리에대해서거의영향을받지않는것으로나타났다
(Fig. 2).
이것은대부분의균주들에대한항균작용에는추출물에포함된단백질성물질이작용을 하지만
B. subtilis
와V. parahaemolyticus
같은특정세균에대 한항균작용에는단백질성물질보다는비단백질성물질이중요 한역할을담당할가능성이있음을의미하는것이다.
진주담치 추출물의 효소 안정성
각추출물내에포함된항균활성물질의특성을확인하기위 해서각추출물
(5 μL)
에trypsin, chymotrypsin
및proteinase K
를포함하는단백질분해효소와lipase
및amylase
를처리한후효소처리전
∙
후의항균활성변화를URDA
법을이용하여B. subtilis
에대해서측정하였다(Fig. 3).
측정결과,
추출물들 은trypsin, amylase, lipase
처리후에도B. subtilis
에대한항균 활성은큰변화없이유지되었으며, chymotrypsin
과proteinase K
처리후에도항균활성이조금감소되었지만거의항균활성 이유지되는것으로나타났다.
이러한결과는추출물에포함된B. subtilis
에대해서항균활성을나타내는물질은단백질,
지 질,
탄수화물과관계가없는물질일가능성이있음을의미하는 것이다.
또한
,
추출물내에포함된특정세균에대한항균활성을나타 내는물질의효소처리에대한시간의존성을확인하기위해서각추출물
(5 μL)
에단백질성항균물질에특히큰영향을미치는trypsin
을1-5
시간처리한후항균활성변화를URDA
법을이 용하여B. subtilis
에대해서측정하였다(Fig. 4).
측정결과,
추 출물들은trypsin
처리후5
시간내에는B. subtilis
에대한항균 활성이큰변화없이유지되는것으로나타났다.
이러한결과는추출물내의항균활성물질은
trypsin
에대한효소안정성이적어도
5
시간정도는유지가될수있음을의미하는것이다.
추출물의 세포독성 확인
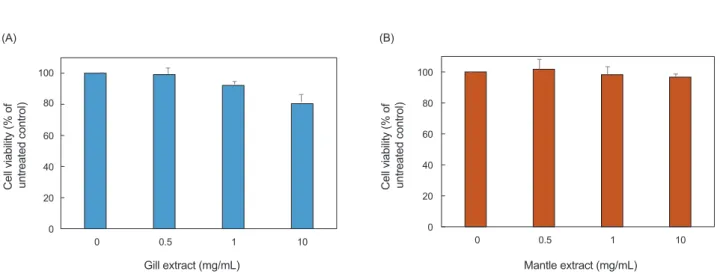
진주담치추출물의세포독성을확인하기위해서
HDF
의세 포증식에 대한영향을MTT assay
를통해서확인하였다(Fig.
5).
추출물을첨가하지않은것(0 mg/mL)
을대조군으로농도별(0.5, 1.0, 10 mg/mL)
로추출물을처리하여cell viability
를측 정하였다.
측정결과,
외투막추출물은10 mg/mL
농도까지거 의세포독성을나타내지않은반면에아가미추출물은약한세 포독성을나타내었다.
이러한결과는HDF
에대해서진주담치 Fig. 5. Cell viability of human dermal fibroblasts after treatment with gill extract (A) and mantle extract (B). The data are expressed cell viability as % of untreated control (Mean±SD, *P<0.05, Mussel extract vs untreated control).(A) (B)
0 20 40 60 80 100
0 0.5 1 10
Cell viability (% of untreated control)
Gill extract (mg/mL)
0 20 40 60 80 100
0 0.5 1 10
Cell viability (% of untreated control)
Mantle extract (mg/mL)
1. Marker (100 bp DNA ladder) 2. Marker+Gill extract 3. Marker+Gill extract + 1% HAc 4. Marker+Proteinase K-treated Gill extract 5. Marker+Trypsin-treated Gill extract 6. Marker+1% HAc
1 2 3 4 5 6
bp
20001600 12001000 900800 700600 500 400 300 200 100
Fig. 6. Cytoplasmic membrane permeabilization of Escherichia coli ML35p by the trypsin/Proteinase K-treated gill extract and pi- scidin 1. Cytoplasmic membrane permeabilization was monitored as an increase in fluorescence intensity by the hydrolysis of the impermeable, chromogenic substrate ONPG in the presence of pi- scidin 1 (1 μg/mL) or extract (5 μL) for 60 min.
Bacillus subtilis KCTC1021
Escherichina coli D31
Gill Mantle
Candida albicans KCTC7965
, 5 mm
(A)
B. subtilis KCTC1021 S. mutans KCCM4 -105 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After Trypsin treatment
Before After Gill extract Mantle extract
, 5 mm Microbes
(B)
B. subtilis KCTC1021 S. mutans KCCM4 -105
C. albicans KCTC7965 S. epidermidis KCTC1917
S. flexneri KCTC2009 E. coli D31 E. coli ML35p C. acnes KCTC11946
V. parahaemolyticus RIMD2110633
Before After
Chymotrypsin treatment Before After Gill extract Mantle extract
, 5 mm Microbes
B. subtilis Gill
Mantle
T C A L P N
, 5 mm
B. subtilis Gill
Mantle
0 h 1 h 2 h 3 h 4 h 5 h
, 5 mm
0 0.2 0.4 0.6 0.8 1
0 10 20 30 40 50 60
Optical Density (O.D.) 405
Time (min)
Trypsin-treated Gill extract Proteinase K-treated Gill extract Piscidin 1
C. albicans KCTC7965
진주담치(Mytilus edulis) 추출물의 항균활성 및 효소 안정성 탐색
285
추출물들은세포독성이낮다는것을의미하는것이다
.
진주담치 추출물에 포함된 항균물질의 막 투과성 확인 실험
진주담치추출물에포함된비단백질성항균물질의상호작용 부위를예측하기위해서
trypsin
과proteinase K
처리된아가미 추출물들의E. coli ML35p
에대한inner membrane permeabi- lization assay
를수행하였다(Fig. 6).
측정결과,
양성대조군으 로사용한piscidin 1
은강한내막투과성을나타낸반면trypsin
또는proteinase K
처리된진주담치아가미추출물은막투과성 을거의나타내지않았다.
이러한결과는진주담치아가미추 출물에포함된비단백질성항균물질은세균세포질내물질의 유출을일으킬정도로세균막에대해서투과성을가지지않으 며세포막과의직접적인상호작용가능성이낮다는것을의미 하는것이다.
진주담치 추출물에 포함된 DNA-결합성 확인 실험
비단백질성항균물질의작용부위가핵내부의DNA
인지를 확인하기위해서trypsin
과proteinase K
가처리된아가미추 출물의DNA migration
저해활성을조사하기위하여DNA- binding assay
를수행하였다(Fig. 7).
측정결과, trypsin
과pro- teinase K
처리된아가미추출물은강한DNA-binding ability
를나타내었다.
이러한결과는진주담치아가미추출물에존재 하는비단백질성항균물질은핵내DNA
와의결합을통해상호 작용할가능성이있다는것을의미하는것이다.
따라서진주담 치추출물에포함된비단백질성항균물질은세포막과직접적 인상호작용보다는intracellular components (
예, DNA)
들에 결합함으로써DNA
복제등의기능에영향을주어서세균증식이억제됨으로써항균작용을나타낼가능성이있음을의미 하는것이다
.
본연구결과들을통해서진주담치추출물은다양한세균에 대해서유의한항균활성을나타내는것으로확인되었으며
,
특 히B. subtilis
와같은그람양성균에대해서는비단백질성항균 물질이중요한역할을담당하는것으로나타났다.
그리고추출 물내의비단백질성항균물질은5
시간동안효소에대해서저 항성을나타내었으며,
세균막과직접상호작용하기보다는세 포내구성물질(intracellular components, DNA
등)
들과의상호 작용을통해서항균활성을나타낼가능성이있을것으로예상 된다.
이러한결과를토대로진주담치추출물은B. subtilis
를포 함하는특정세균에대한효소저항성강화를위한첨가물로서 활용가능할것으로판단된다.
사 사
이논문은
2014
년도정부(
해양수산부)
의재원으로해양수산 과학기술진흥원해양수산생명공학기술개발사업의지원을받 아수행된연구입니다(No. 20140447).
References
Chung KT. 2019. Aging of immune system. J Life Sci 29, 817- 823. https://doi.org/10.5352/JLS.2019.29.7.817.
Charlet M, Chernysh S, Philippe H, Hetru C, Hoffmann JA and Bulet P. 1996. Innate immunity: Isolation of several cyste- ine-rich antimicrobial peptides from the blood of a mollusk Mytilus edulis. J Biol Chem 271, 21808-21813. https://doi.
org/10.1074/jbc.271.36.21808.
Fig. 7. Gel retardation analysis or the binding of the proteinase K or trypsin treated gill extract to DNA. Binding of the enzyme-treated ex- tract (5 μL) to DNA was assessed by measuring the retardation of commercial 100 bp DNA ladder migration through an agarose gel (1.4%
gel). Lane 1, 100 bp DNA ladder (0.2 ng); lane 2, gill extract in D.W.; lane 3, gill extract in 1% HAc; lane 4, Proteinase K-treated gill extract;
lane 5, trypsin-treated gill extract; lane 6, negative control, 1% HAc.
(A) (B)
0 20 40 60 80 100
0 0.5 1 10
Cell viability (% of untreated control)
Gill extract (mg/mL)
0 20 40 60 80 100
0 0.5 1 10
Cell viability (% of untreated control)
Mantle extract (mg/mL)
1. Marker (100 bp DNA ladder) 2. Marker+Gill extract 3. Marker+Gill extract + 1% HAc 4. Marker+Proteinase K-treated Gill extract 5. Marker+Trypsin-treated Gill extract 6. Marker+1% HAc
1 2 3 4 5 6
bp
20001600 12001000 900800 700600 500 400 300 200 100
이지은
ㆍ
서정길286
Clark R and Kupper T. 2005. Old meets new: the interac- tion between innate and adaptive immunity. J Invest Dermatol 125, 629-637. https://doi.org/10.1111/j.0022- 202X.2005.23856.x.
Hancock REW and Diamond G. 2000. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol 8, 402-410. https://doi.org/10.1016/s0966- 842x(00)01823-0.
Hsu CH, Chen C, Jou ML, Lee AYL, Lin YC, Yu YP, Huang WT and Wu SH. 2005. Structural and DNA-binding studies on the bovine antimicrobial peptide, indolicidin, evidence for multiple conformations involved in binding to mem- branes and DNA. Nucl Acids Res 33, 4053-4064. https://
doi.org/10.1093/nar/gki725.
Hubert F, Noel T and Roch P. 1996. A member of the arthro- pod defensing family from edible Mediterranean mussels Mytilus galloprovincialis. Eur J Biochem 240, 302-306.
https://doi.org/10.1111/j.1432-1033.1996.0302h.x.
Kim CS and Li SH. 2012. The effects of pini folium extract on collagen expression and synthesis in normal human dermal fibroblasts. Kor J Aesthet Cosmetol 10, 817-822.
Kim H, Jang JH, Kim SC and Cho JH. 2013. De novo genera- tion of short antimicrobial peptides with enhanced stability and cell specificity. J Antimicrob Chemother 69, 121-132.
https://doi.org/10.1093/jac/dkt322.
Lee IH, Cho Y and Lehrer RI. 1997. Effects of pH and salin- ity on the antimicrobial properties of clavanins. Infect Im- mun 65, 2898-2903. https://doi.org/10.1128/IAI.65.7.2898- 2903.1997.
Lehrer RI, Rosenman M, Harwig SSL, Jackson R and Eisenhaur P. 1991. Ultrasensitive assay for endogenous antimicrobial polypeptides. J Immunol Methods 137, 167-173. https://doi.
org/10.1016/0022-1759(91)90021-7.
Mitta G, Hubert F, Noel T and Roch P. 1999. Myticin, a novel cysteine-rich antimicrobial peptide isolated from haemo- cytes and plasma of the mussel Mytilus galloprovincialis.
Eur J Biochem 265, 71-78. https://doi.org/10.1046/j.1432- 1327.1999.00654.x.
Oh R, Lee MJ, Kim YO, Nam BH, Kong HJ, Kim JW, Park JY, Seo JK and Kim DG. 2018. Purification and characteriza- tion of an antimicrobial peptide myitichitin-chitin binding domain from the hard-shelled mussel Mytilus coruscus. Fish Shellfish Immunol 83, 425-435. https://doi.org/10.1016/j.
fsi.2018.09.009.
Relf JM, Chisholm JRS, Kemp GD and Smith VJ. 1999. Purifi- cation and characterization of a cystein-rich 11.5-kDa anti- bacterial protein from the granular haemocytes of the shore crab Carcinus maenas. Eur J Biochem 264, 350-357. https://
doi.org/10.1046/j.1432-1327.1999.00607.x.
Rowley AF and Powell A. 2007. Invertebrate immune systems specific, quasi-specific, or nonspecific?. J Immunol 179, 7209-7214. https://doi.org/10.4049/jimmunol.179.11.7209.
Seo JK. 2016. Screening and purification of an antimicrobial peptide from the gill of the Manila clam Ruditapes philip- pinarum. Korean J Fish Aquat Sci 49, 137-145. https://doi.
org/10.5657/KFAS.2016.0137.
Seo JK, Crawford JM, Stone KL and Noga EJ. 2005. Purifica- tion of a novel arthropod defensin from the American oyster Crassostrea virginica. Biochem Biophys Res Commun 338, 1998-2004. https://doi.org/10.1016/j.bbrc.2005.11.013.
Seo JK, Kim DG, Oh R, Park KS, Lee IA, Cho SM, Lee GY and Nam BH. 2017. Antimicrobial effect of the 60S ribosomal protein L29 (cgRPL29), purified from the gill of pacific oys- ter Crassostrea gigas. Fish Shellfish Immunol 67, 675-683.
https://doi.org/10.1016/j.fsi.2017.06.058.
Silphaduang U and Noga EJ. 2001. Peptide antibiotics in mast cells of fish. Nature 414, 268-269. https://doi.
org/10.1038/35104690.
Skerlavaj B, Romeo D and Gennaro R. 1990. Rapid membrane permeabilization and inhibition of vital functions of Gram- negative bacteria by bactenecins. Infect Immun 58, 3724- 3730. https://doi.org/10.1128/IAI.58.11.3724-3730.1990.
Tincu JA and Taylor SW. 2004. Antimicrobial peptides from ma- rine invertebrates. Antimicrob Agents Chemother 48, 3645- 3654. https://doi.org/10.1128/AAC.48.10.3645-3654.2004.
Tom D, Patrick S and Peter B. 2011. Alternatives to antibiot- ics for the control of bacterial disease in aquaculture. Curr Opin Microbiol 14, 251-258. https://doi.org/10.1016/j.
mib.2011.03.004.
van’'t Hof W, Veerman EC, Helmerhorst EJ and Amerongen AV. 2001. Antimicrobial peptides: properties and applica- bility. Biol Chem 382, 597-619. https://doi.org/10.1515/
BC.2001.072.
Wang XW, Zhao XF and Wang JX. 2014. C-type lectin binds to beta-integrin to promote hemocytic phagocytosis in an invertebrate. J Biol Chem 289, 2405-2414. https://doi.
org/10.1074/jbc.M113.528885.
Yu XQ, Tracy ME, Ling E, Scholz FR and Trenczek T. 2005.
A novel C-type immulectin-3 from Manduca sexta is trans- located from hemolymph into the cytoplasm of hemo- cytes. Insect Biochem Mol Biol 35, 285-295. https://doi.
org/10.1016/j.ibmb.2005.01.004.
Zasloff M. 2002. Antimicrobial peptides of multicellular organ- isms. Nature 415, 389-395. https://doi.org/10.1038/415389a.